
刺槐、红花刺槐、四倍体刺槐花粉体外萌发对比1)
2012-06-13 06:20:30蒋晋豫胡瑞阳孙宇涵袁存权
东北林业大学学报 2012年1期
戴 丽 孙 鹏 蒋晋豫 胡瑞阳 孙宇涵 袁存权 李 云
(林木育种国家工程实验室(北京林业大学),北京,100083)
刺槐(Robinia pseudoacaciaL)又名洋槐,属于蝶形花科刺槐属。原生于北美洲阿巴拉契亚山脉和奥萨克山脉一带,17世纪初被引入欧洲,20世纪初引入我国[1],分布区面积达 1 100 km×1 500 km,是当今世上造林面积仅次于杨树和桉树的速生阔叶树种[2-3]。刺槐生长迅速,木材坚韧,纹理细致,有弹性,耐水湿,抗腐朽,是重要的速生用材树种。刺槐根系发达,具根瘤,有一定的抗旱、抗烟、耐盐碱能力,是华北、西北等地区优良的保持水土、防风固沙、改良土壤和“四旁”绿化树种[4],刺槐的花粉可酿蜜[5],枝叶可用作牲畜饲料及绿肥[6],是优良蜜源树种和饲料树种,深受群众欢迎。
自从我国开展刺槐遗传育种研究工作以来,主要进行了刺槐速生良种选育、刺槐用材无性系选育和刺槐观赏、饲用等专用品种选育,改良工作主要局限在优良无性系选育及国外引进新品种上[7]。在控制杂交育种方面涉及很少,而杂交育种是一种非常有效的增加变异的手段[8]。搞好刺槐的杂交育种工作,对于我国刺槐育种和新品种选育至关重要。授粉受精是杂交工作的基础,花粉的萌发和生长是授粉受精的关键问题之一。特别是生长在不同区域,不同气候条件下的刺槐,以及不同单株、无性系,甚至不同种的刺槐,常常表现花期不遇。有的还表现花粉活力低、不萌发、生长差、难以实现授粉受精,从而致使杂交失败。同时花粉资源保存、跨地域开展杂交,以及不同优株、不同种之间的杂交,都离不开花粉的保存和活力鉴定技术,综上所述,能够快速准确测定刺槐花粉活力对于刺槐的杂交育种具有重要意义。
植物花粉生活力检测的方法较多,如花粉染色测定法、氯化三苯基四氮唑染色法(TTC法)、I-KI染色法、联苯胺法、荧光染色法、花粉离体萌发测定法,以及花粉授粉结实检测法等,其中花粉离体萌发测定方法简单、迅速、合理[9]。前人对于刺槐花粉活力的研究并不多,多数使用染色法,如田志和等[10]在研究刺槐开花结实生物学特性时使用亚甲基蓝和蔗糖的混合溶液测定花粉的生活力,原法宪[11]使用的联苯胺—甲萘酚染色的方法。在香花槐[12]和四倍体刺槐[13]花粉生活力的研究中使用TTC染色法。王念等[14]是唯一使用培养基法测定金叶刺槐和荷刺Ⅰ号花粉生活力的人,培养基使用15%蔗糖+0.001%硼酸。至于蔗糖、硼酸和Ca2+对于刺槐花粉萌发的影响还未见报道。蔗糖、硼酸和Ca2+作为花粉离体培养的基本组成成分在木槿[15]、百合[16]、小麦[17]、杨树[18]等很多植物中均有应用。但不同植物所需的质量浓度范围有一定的差异。本试验以刺槐、红花刺槐(R.pseudoacaciavar.decaisneanaCarr.)和四倍体刺槐(TetraploidR.pseudoacacia.L)的新鲜花粉为材料,采用正交设计,研究花粉的离体萌发最适培养基成分,其结果可为刺槐花粉生殖生物学研究、种质资源保存、杂交育种提供技术基础。
1 材料与方法
刺槐、红花刺槐和四倍体刺槐的花药和花粉材料分别采自北京市延庆县米家堡林场的刺槐林基地、北京林业大学校园的红花刺槐观赏绿化树,以及北京市延庆县北张庄的四倍体刺槐基地。
在材料来源地的刺槐盛花期分别收集3种刺槐的花药(b)和花粉(k)。花药采集于L(花冠)∶L(花萼)=5 ∶2(长度比),且未散粉的时期(图 1a、b、c),花粉采集于小花刚刚开放时期。将收集到的花药与花粉自然晾干一昼夜。将刺槐、红花刺槐、四倍体刺槐开花散粉后直接采集的花粉分别记为刺槐k、红花刺槐k、四倍体刺槐k;采集的花药开裂后收集的花粉记为刺槐b、红花刺槐b、四倍体刺槐b。将k和b两种不同方式下收集的花粉均匀涂抹在培养基上,在(25±1)℃条件下培养6 h后用13395H2X徕卡显微镜观察并照相,同时,测量不同培养基上花粉管生长长度,每个处理3次重复,每个重复观察3个视野。以花粉管长度超过花粉粒直径作为萌发标准,萌发率=(已萌发的花粉粒数/花粉粒总数)×100%。

图1 3种刺槐花外形及其花粉萌发
以2%琼脂为基本培养基。采用L16(45)正交设计,研究蔗糖、硼酸、硝酸钙3个因素的4个水平对刺槐花粉在不同花粉采集期的萌发和生长影响。试验结果采用DPS 7.05进行分析。
2 结果与分析
2.1 不同培养基对刺槐花粉萌发率的影响
蔗糖和硼酸是影响花粉萌发率的重要因素。蔗糖不仅为花粉萌发和花粉管生长提供营养,还有维持培养环境渗透压的作用[19]。硼酸可增加花粉对糖的吸收、运转及代谢,增加氧的吸收,并促进果胶物合成[19-20]。本试验观察发现,刺槐、红花刺槐以及四倍体刺槐的花粉均在低质量浓度(50 g·L-1)蔗糖的培养基上萌发率低,仅为1.6% ~23.5%,同时表现花粉壁和花粉管顶端破裂,释放出其内含物到培养基上(图1d、e、f),但当蔗糖质量浓度提高后(100~200 g·L-1),该不良现象消失。在低蔗糖质量浓度(50 g·L-1)不变的情况下,随着硼和钙质量浓度的增加,花粉管伸长生长明显,但其顶端膨大至破裂(图1g、h、i)。从而看出,硼和钙能够促进刺槐花粉管的伸长生长,但并不能提高培养基的渗透压,花粉管仍然不能正常生长。此外,在本试验所设置的蔗糖质量浓度梯度50~200 g·L-1中,始终没有发现质壁分离现象。
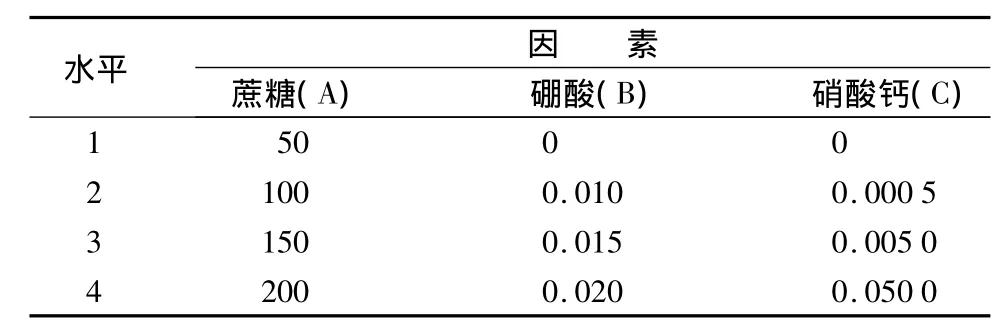
表1不同培养基对刺槐花粉离体萌发影响的正交设计g·L-1
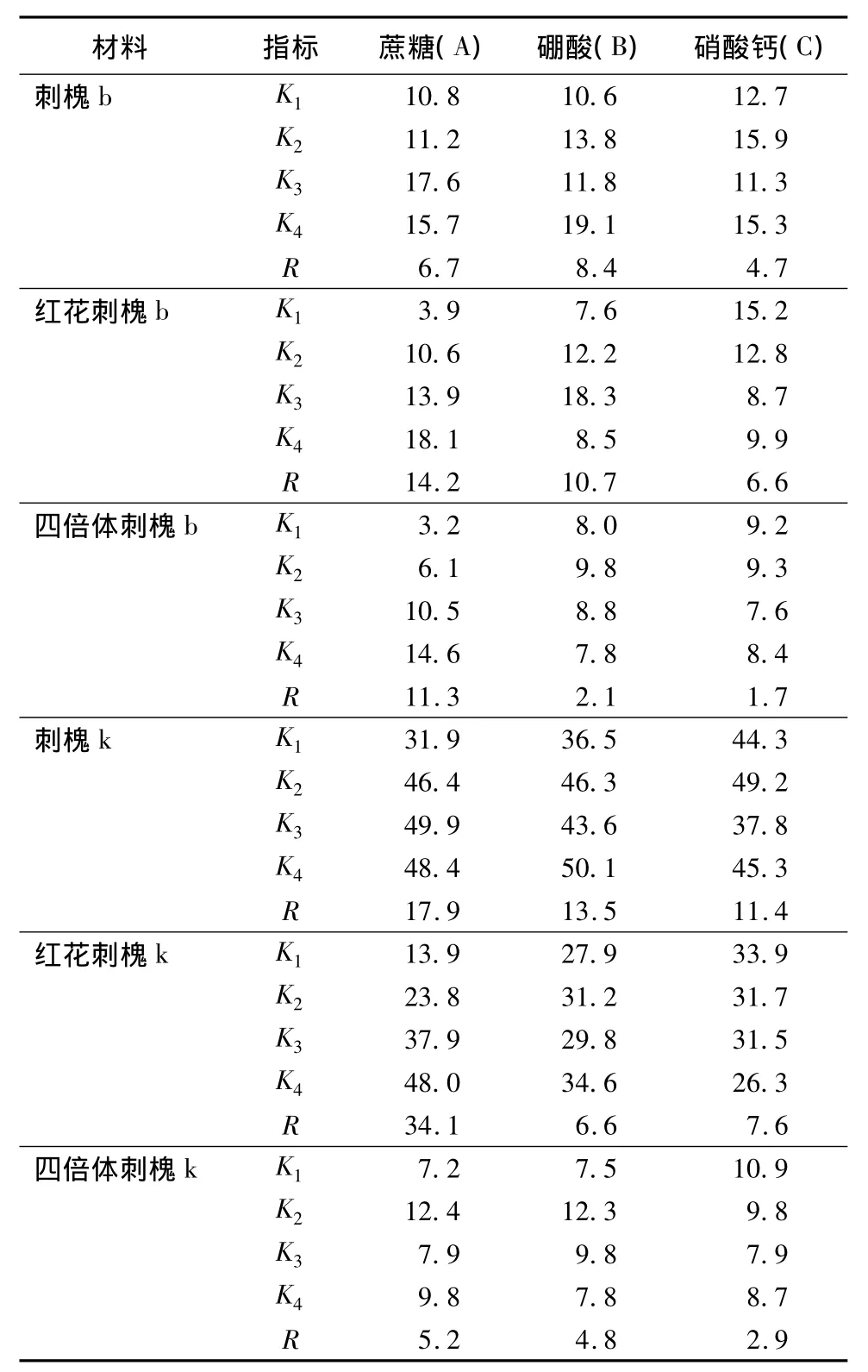
表2 不同培养基对刺槐萌发率影响的正交试验极差分析%
从表2的各因素不同水平的萌发率可以找出每种花粉的最适培养基质量浓度。刺槐b和k都为A3B4C2,萌发率分别为39.6%和68.2%(图1k、l);红花刺槐b:A4B3C1,红花刺槐k:A4B4C1(萌发率为54.5%,图1j);四倍体刺槐b:A4B2C2,四倍体刺槐k:A2B2C1。

表3 不同培养基对刺槐花粉花粉管长度影响的正交试验分析 μm
2.2 花粉的不同采集时期与刺槐花粉萌发培养基的关系
从表2和表4可以看出,除刺槐b之外,蔗糖的萌发率极差最大,说明蔗糖对3种刺槐不同时期的花粉萌发影响最大,方差分析也达到了显著水平。除红花刺槐k之外,硼酸对3种刺槐不同时期采集的花粉萌发影响次之,最后是硝酸钙。说明蔗糖、硼酸和硝酸钙对3种刺槐花粉萌发的影响大体相同,符合由大到小的顺序为:蔗糖、硼酸、硝酸钙的规律。对花粉萌发影响比较特殊的有:刺槐b由大到小的顺序为:硼酸、蔗糖、硝酸钙;红花刺槐k由大到小的顺序为:蔗糖、硝酸钙、硼酸,这说明硼对刺槐天然散粉前采集的花粉离体萌发的影响大于蔗糖和硝酸钙,而钙离子对红花刺槐天然散粉花粉体外萌发的影响大于硼离子。
刺槐和红花刺槐天然散粉后的平均花粉萌发率(平均萌发率为所有组合平均值)明显高于采集花药后收集的平均花粉萌发率(普通刺槐b、k分别为13.8% 和 44.2%,红花刺槐 b、k 分别为 11.6% 和30.9%)。说明二倍体刺槐在天然散粉前的一段时间内还在进行最后的成熟生长,这一段时间对于二倍体刺槐生殖生长很重要,影响花粉后期能否正常萌发。同二倍体刺槐相比,四倍体刺槐两种花粉的平均萌发率相差不多(四倍体刺槐b、k分别为:8.6%和9.3%),可见四倍体刺槐天然散粉前就已经达到成熟,但萌发能力远远低于二倍体,表现为花粉粒大,花粉活性差。
2.3 不同培养基对刺槐花粉管伸长生长的影响
花粉的萌发与花粉管的伸长生长是一个复杂的生理生化反应过程,许多研究表明硼和钙参与了花粉管的伸长生长,并且该体系已被越来越多地用作研究细胞生长的分子调控机理,近年来的许多研究结果证明细胞外的钙离子参与了这一过程[21-22]。离体生长的花粉管需要不断地从外界吸收Ca2+,以维持花粉管顶端的Ca2+梯度分布,为其极性生长创造条件[23]。木槿中也证实在有钙和无钙加入的情况下,有钙明显比无钙条件下花粉管伸长快,且增加的幅度也很明显[15]。硼与钙在花粉萌发过程中有着密不可分的关系,硼处理的百合花粉细胞内游离钙离子质量浓度明显高于无硼处理的质量浓度,并且细胞内游离钙离子变化属于剂量依赖型,即依赖于培养基中的硼酸质量浓度,随硼酸质量浓度升高而升高,但过高质量浓度的钙和硼又会抑制花粉的萌发和花粉管的伸长生长[16]。
为了找出最适硼酸和硝酸钙质量浓度,花粉培养6 h后测量花粉管长度,并进行分析。从表3和表4可以看出,蔗糖对红花刺槐和四倍体刺槐花粉管的伸长影响最大,对于刺槐花粉管的伸长影响不如硼酸大。硼酸对刺槐和红花刺槐的影响大于硝酸钙,而在四倍体刺槐花粉管生长过程中,硝酸钙的影响大于硼酸,这就说明即使同一树种,不同花色与不同倍性之间的生长过程中所需营养元素的量有所差异。从各水平的平均值得出3种刺槐的花粉管生长最适培养基,普通刺槐b、k:A2B4C3,花粉管长度分别达347 μm 和 330 μm(图 1m、n);红花刺槐 b:A2B3C3,红花刺槐 k:A2B3C2;四倍体刺槐 b:A4B3C2,花粉管长度达206 μm(图1o),四倍体刺槐k:A4B3C3。从最优组合可看出,刺槐所需硼酸质量浓度最高,四倍体刺槐所需蔗糖质量浓度最高,且每个因素对花粉管的影响都与对花粉萌发率的影响不同。但红花刺槐和四倍体刺槐各水平之间没有达到显著性差异,刺槐的各水平之间达到显著性差异。由多重比较可看出,刺槐b和k的蔗糖质量浓度A1、A3、A4之间无显著差异,A2水平显著高于前3个水平,且在显微镜下观察,其花粉管细长,无破裂扭曲现象。

表4 正交设计方差分析
3 结论与讨论
豆科植物具有闭花授粉的习性,在开花前已经散粉,所以研究散粉前的花粉活力对于选择去雄时间有一定的参考价值。在菜豆的开花习性研究中得出菜豆在花器居中时花粉生活力最大,结合菜豆闭花授粉的特性,应在花器更小时选择杂交亲本[24]。试验结果表明3种刺槐在天然散粉前的花粉活力低,最高平均萌发率为13.8%,此时花器发育达到最大且未散粉,便于操作,因此,可以考虑将此时作为去雄时间。
目前的许多研究认为,在植物的开花期内,盛花期采集的花粉生活力较好。如广东荔枝同一品种一般盛花期比初、末花期的花粉生活力强[25]。沙棘盛花期花粉的生活力明显高于初花期花粉[26]。葡萄4个不同发育时期中,开花盛期有生活力花粉的比率最高[27]。在本试验中,刺槐和红花刺槐的花粉活力表现同前人研究结果相同,开花后采集的花粉萌发率明显高于开花前采集的花粉萌发率,然而,四倍体刺槐开花前后2种花粉的平均萌发率相差不多,这可能是因为染色体组加倍以后加速了花粉成熟。
染色体组加倍以后大大降低了四倍体刺槐花粉的活力。四倍体刺槐属于同源多倍体,染色体成倍数性增加,4条同源染色体可能发生不联会和四价体提早解离的情况。这使得孢子母细胞在减数分裂时形成多种不同组合的分离方式,在后期Ⅰ分离时可能是2/2式均衡分离,也可能是3/1或2/1或1/1式不均衡分离。相对于未加倍的刺槐和红花刺槐而言,四倍体刺槐染色体的不正常分离造成花粉活力降低。花粉活力低对四倍体刺槐与刺槐之间的杂交造成了一定的困难。
本试验研究了蔗糖、硼酸、硝酸钙对花粉体外萌发及花粉管生长的影响。综上可知,3个因素在花粉萌发过程中起着不同的作用,在自然条件下,花粉在柱头上生长所需的营养物质及矿物质元素是由柱头和花柱来补偿的。在花粉离体培养中,不同植物种类所需的蔗糖、硼酸、硝酸钙的质量浓度有所不同。例如,金光杏梅[19]最适蔗糖质量浓度为20 g·L-1,最适硼酸质量浓度为 3 g·L-1;小黑杨[18]的最适蔗糖质量浓度为150 g·L-1,最适硼酸质量浓度为0.02 g·L-1。即使是同一个种的不同品种也有不同的最适蔗糖质量浓度,如菜豆[24]中的早油豆王、园丰907、油豆王等8个品种的最适质量浓度均在10%左右,油豆和白花菜豆为15%左右,红花菜豆则在5%左右。研究结果表明,刺槐和红花刺槐b的最适蔗糖质量浓度为15 g·L-1,红花刺槐k和四倍体刺槐b的最适蔗糖质量浓度为20 g·L-1,四倍体刺槐k的最适蔗糖质量浓度为10 g·L-1,从而可知刺槐散粉前后的花粉萌发所需蔗糖质量浓度不变,红花刺槐散粉后比散粉前的花粉萌发所需蔗糖质量浓度高,四倍体刺槐散粉后比散粉前的花粉萌发所需蔗糖质量浓度低。硼酸和硝酸钙的最适质量浓度也因不同种和不同的采集时间而有所不同,总结得出3种刺槐花粉萌发的适宜培养基,刺槐b和k 都为蔗糖0.15 kg·L-1+硼酸20.00 mg·L-1+硝酸钙 0.50m g·L-1;红花刺槐 b:蔗糖0.20k g·L-1+硼酸 15.00 mg·L-1,红花刺槐 k:蔗糖 0.20 kg·L-1+硼酸 20.00 mg·L-1;四倍体刺槐 b:蔗糖 0.20 kg·L-1+硼酸 10.00 mg·L-1+硝酸钙 0.50 mg·L-1,四倍体刺槐 k:蔗糖 0.10 kg·L-1+硼酸 10.00 mg·L-1。其中蔗糖对3种刺槐的花粉萌发的影响最大,其次是硼酸,最后是硝酸钙。从不同花粉最适培养基对比可以看出,红花刺槐花粉萌发可以没有外源钙的参与,刺槐和四倍体刺槐对外源钙的质量浓度要求也不高,说明刺槐花粉内部的钙库足以支持花粉的正常萌发,而在花粉管的生长过程中,有钙比无钙培养基的花粉管生长要快。
[1]茹桃勤,李吉跃,张克勇,等.国外刺槐(Robinia pseudoacacia)研究[J].西北林学院学报,2005,20(3):102-107.
[2]战臣祥,李观和,袁俊云,等.刺槐良种基因材料收集与选育研究[J].山东林业科技,2010(2):18-21.
[3]王文君,罗晓丹,张日清,等.刺槐种子萌发过程中的生理生化变化[J].河南林业科技,2009,29(2):1-3.
[4]荀守华,乔玉玲,张江涛,等.我国刺槐遗传育种现状及发展对策[J].山东林业科技,2009(1):92-96.
[5][匈]凯莱斯台舍.刺槐[M].王世绩,张敦伦,译.北京:中国科学技术出版社,1993.
[6]郑万钧.中国树木志:2册[M].北京:中国林业出版社,1985:1360-1361.
[7]解荷锋,于中奎,陈一山,等.种子园刺槐开花结实和控制授粉的初步研究[J].山东林业科技,1994(4):4-7.
[8]刘芳,周蕴薇.花粉的保存及生活力测定方法的探讨[J].南方农业,2007,1(3):70-71.
[9]王钦丽,卢龙斗,吴小琴,等.花粉的保存及其生活力测定[J].植物学通报,2002,19(3):365-373.
[10]田志和,董健,李继刚.刺槐开花结实生物学特性的观察初报[J].辽宁林业科技,1981(2):18-20.
[11]原法宪.刺槐开花生物学特性观察[J].山西林业科技,1987(3):1-10.
[12]刘德光.香花槐分类地位及不结实现象研究[D].合肥:安徽农业大学,2008.
[13]郝晨.饲料型四倍体刺槐生殖生物学特性的初步研究[D].北京:北京林业大学,2006.
[14]王念,王文君,赵蓬晖,等.温度和时间对储藏刺槐花粉生活力影响的研究[J].河南林业科技,2007,27(4):7-9.
[15]贾文庆,刘会超,郭丽娟.外源钙对木槿花粉萌发的影响[J].山西农业科学,2007,35(6):56-58.
[16]年玉欣,罗凤霞,张颖,等.测定百合花粉生命力的液体培养基研究[J].园艺学报,2005,32(5):922-925.
[17]朱展望,张改生,牛娜.小麦花粉的离体萌发研究[J].麦类作物学报,2007,27(1):12-15.
[18]赵丽娟,李淑娟,于金梅,等.小黑杨花粉离体萌发和细胞学分析[J].林业科学,2011,47(6):36-41.
[19]余义和,李桂荣,王新娟,等.蔗糖和硼酸浓度对金光杏梅花粉离体萌发的影响[J].山西果树,2006(4):7-8.
[20]郭光明,张福锁,尚忠林,等.硼对百合花粉萌发过程中细胞内游离钙离子的影响[J].中国农业大学学报,2002,7(5):32-37.
[21]Franklin-Tong V E.Signaling and the modulation of pollen tube growth[J].Plant Cell,1999,11:727-738.
[22]Reiss H D,Herth W,Nobiling R.Development of membraneand calcium-gradients during pollen germinat-ion ofLilium longiflorum[J].Planta,1985,163:84-90.
[23]田惠桥,远彤.钙在被子植物受精过程中的作用[J].植物生理学报,2000,26(5):369-380.
[24]王亚君,朱博.不同品种菜豆的开花习性和花粉生活力的研究[J].吉林蔬菜,2008(6):85-87.
[25]翁树章,李青英,陈育英,等.广东荔枝主要品种花粉生活力研究初报[J].果树科学,1990,7(2):91-96.
[26]郭荫槐,姜辉.沙棘粉生活力测定和贮藏试验[J].辽宁林业科技,1995(1):28-30.
[27]万怡震,贺普超,廖祥儒,等.葡萄花粉不同发育阶段、不同干燥及不同保存条件下生活力鉴定研究[J].陕西农业科学,1997(2):9-10.
[28]姜金仲,李云,贺佳玉,等.四倍体刺槐胚珠败育及其机制[J].林业科学,2011,47(5):40-46.
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:09:08
中国果业信息(2022年1期)2022-11-28 07:43:38
三农资讯半月报(2020年10期)2020-06-08 15:32:59
无机盐工业(2019年5期)2019-12-26 04:51:53
肥料与健康(2019年6期)2019-03-19 06:32:04
生物学杂志(2018年4期)2018-08-15 11:21:32
现代园艺(2017年13期)2018-01-19 02:28:17
中国果业信息(2017年8期)2017-01-13 20:06:36
中成药(2016年4期)2016-05-17 06:07:54
天津建设科技(2016年6期)2016-02-05 02:25:30
