
6种野生鸢尾种皮、种苗特征与进化的关系1)
2012-06-13 06:20陈士惠卓丽环
东北林业大学学报 2012年1期
孙 颖 陈士惠 卓丽环 王 玲
(东北林业大学,哈尔滨,150040) (上海农林职业技术学院) (东北林业大学)
鸢尾属是单子叶植物鸢尾科中最大的属,也是最进化的属。野生的鸢尾属植物生命力强,抗逆性强,绿色期长,观赏价值高,有大面积推广应用的前景。目前世界鸢尾属的研究主要集中在杂交育种、创制新品种上[1-5]。有关系统发育理论方面的研究,在国外的鸢尾属植物相关文章中有一定涉及[6-8],国内研究的较少[9-10]。对于我国东北野生鸢尾的种子相关结构的研究,将为更广阔地区乃至全国的鸢尾资源的系统研究和生产应用提供部分理论依据。
1 试验材料
试验材料于东北林业大学帽儿山实验林场、哈尔滨师范大学家属区院内、沈阳市植物园的鸢尾园采集(表1)。根据文献[11]所做的对60种鸢尾的数量分类学研究,将中国鸢尾属植物划分为11类,从中可以看出,本研究所选出的东北鸢尾均属于第一类无附属物亚属12个种中的6种。这是鸢尾属中最原始的类群,彼此间的亲缘关系较近,是一个很自然的类群。
2 试验方法
2.1 种皮结构观察
采用日本大学JEOL公司生产的JSM—5610LV型扫描电子显微镜进行观察,每个种观察3粒,并选择图像清晰、具代表性区域的画面拍照。拍照部位均为种子背面,种皮雕纹纹饰分别放大300倍及1 000倍。

表1 材料来源
2.2 种苗发育过程观测
采用常规发芽试验。出苗期观察萌发的种子,每种30粒,记录其发育过程,记录胚根、子叶鞘、真叶出现的时间。于萌发后7 d测量胚根、子叶鞘、子叶联结的长度。
3 结果与分析
3.1 种皮超显微结构
观察结果表明,6种鸢尾种皮超显微形态特征差异明显。玉蝉花初级雕纹为条网状纹饰。网眼凹陷较深,由长条状的四边形或五边形组成。网脊凸起,较粗,高度一致。各网眼的网脊界限清晰。次雕纹由次生壁加厚的不规则波状乳突构成(图1(a)、图1(b))。溪荪初级雕纹为条网状纹饰。网眼较平整,由不规则的长条状五边形或六边形组成,近似鱼鳞状。网脊为单条纹,狭窄,隆起程度差。各网眼的网脊界限清晰。次雕纹较光滑(图1(c)、图1(d))。北陵鸢尾初级雕纹为网状纹饰。网眼凹陷,由近规则的正五边形和正六边形组成。网脊突出,但没有玉蝉花的网脊明显。网脊较粗,高度不一致。各网眼的网脊界限较清晰,个别地方模糊。次雕纹由次生壁加厚的不规则条状网纹构成(图1(i)、图1(j))。马蔺初级雕纹为蜂窝状纹饰。网眼凹陷,由较为规则的正五边形或六边形组成。网脊凸出,较粗,宽度一致,高度近一致。各网眼的网脊界限十分清晰。次雕纹较光滑,个别为由次生壁加厚的不规则疣状乳突构成(图1(k)、图1(l))。黄菖蒲初级雕纹为条网状纹饰。网眼凹陷,由长条状的四边形或五边形组成。网脊凸起,较粗,高度一致。各网眼的网脊界限清晰。次雕纹由次生壁加厚的均匀分布的疣状乳突构成(图1(e)、图1(f))。燕子花初级雕纹为条网状纹饰。网眼较平整,由不规则的长条状五边形或六边形组成,近似鱼鳞状。网脊较窄,隆起程度差,高度一致。各网眼的网脊界限清晰。次雕纹由次生壁加厚的不规则条状网纹构成(图1(g)、图1(h))。
3.2 出苗期种苗结构特征
种子萌发时,下胚轴不发育或不伸长,只有上胚轴和胚芽迅速向上生长,形成幼苗的主茎,而子叶始终留在土壤中,这种方式形成的幼苗,称为子叶留土幼苗。本文所研究的6种鸢尾科植物均为子叶留土型。这种类型植物的子叶明显区别出子叶鞘、子叶联结和吸器。子叶鞘起着保护胚芽的作用,吸器是吸收胚乳的营养物质,子叶联结起着前两者之间联系的作用。更进化的种类在此基础上发育出另一些鞘状结构,称为舌状鞘、假舌状鞘。
解剖镜下观测发现,鸢尾属植物种子萌发时胚突破种皮的一端为圆钝形,当露白达到2~3 mm时,胚逐渐变尖,呈锥状,这是形成的胚根。这种现象与种胚在种子内部的状态有关。在种子内,种胚靠近发芽口的一侧即为圆钝状,远离发芽口的一侧呈锥状。大部分玉蝉花、黄菖蒲的胚根密生根毛,溪荪和北陵的根部具有较密根毛的比例很低,只有28%的马蔺根部生有根毛。在对鸢尾科射干属射干的同类研究中发现,旱生的射干萌发后,只有16%的胚根生有较显著的根毛。由此认为:旱生的鸢尾趋向于根毛稀疏,而湿生种类则趋向于密生根毛。
观察种苗可以看出(表2):溪荪、北陵、黄菖蒲、燕子花子叶只有基鞘,没有舌状鞘,这4种植物的子叶分为子叶鞘(即基鞘)、细长的子叶联结和棒状的吸器3个部分,细长的子叶联结下弯,真叶披针形而具弧形脉或剑形而具平行脉;玉蝉花幼苗子叶的基鞘和舌状鞘均发达,子叶联结下弯与子叶鞘部分或完全贴合,使原来的子状鞘变成了假舌状鞘(包括基鞘与舌状鞘),真叶披针形;马蔺的幼苗只有舌状鞘而基鞘完全退化,子叶联结不但和子叶鞘完全贴合,而且还继续伸延和下胚轴完全贴合,形成了中胚轴。以上6个种的子叶、真叶皆两侧压扁,鞘部套褶。

表2 6种鸢尾种苗子叶联结及子叶鞘的长度 mm
文献[12]将单子叶植物的幼苗类型分为21种。据此归类,溪荪、北陵、黄菖蒲、燕子花属于鸭跖草型,这个类型的最大特点是细长的子叶联结下弯;玉蝉花属于蓝耳草型,这个类型的最大特点是子叶联结下弯和子叶鞘部分或完全贴合,使原来的子叶鞘变成了假舌状鞘,由于贴合程度有异,子叶联结的游离部分可位于假舌状鞘的不同位置上;马蔺属于迦通尼型,因为马蔺的幼苗只有舌状鞘而基鞘完全退化,子叶联结不但与子叶鞘完全贴合,而且还继续伸延和下胚轴完全贴合,形成中胚轴。
4 结论与讨论
从演化趋势看,Corner[13]认为单子叶植物的种子较大并且种皮纹饰简单的是原始特征,种子较小并且种皮纹饰复杂的为次生特征。因此可认为,鸢尾属种皮微形态特征的网脊龙骨状、网眼较浅的是较原始特征,表面有凹点或网脊较宽、网眼较深的是较进化的特征。由此初步推断,鸢尾属6种植物的进化关系是溪荪→北陵→燕子花→黄菖蒲→玉蝉花→马蔺。
马蔺种皮表面为蜂窝状纹饰。从形态结构上推测,这种微形态有利于它们在干旱环境下吸收和保留水分,保水力为蜂窝状>网状。这与前人[14-16]在其他种类上对种皮微形态的研究结论是一致的,显示了旱生植物结构与功能的高度统一。同时也说明生态环境深刻地影响着植物形态结构的形成和差异。
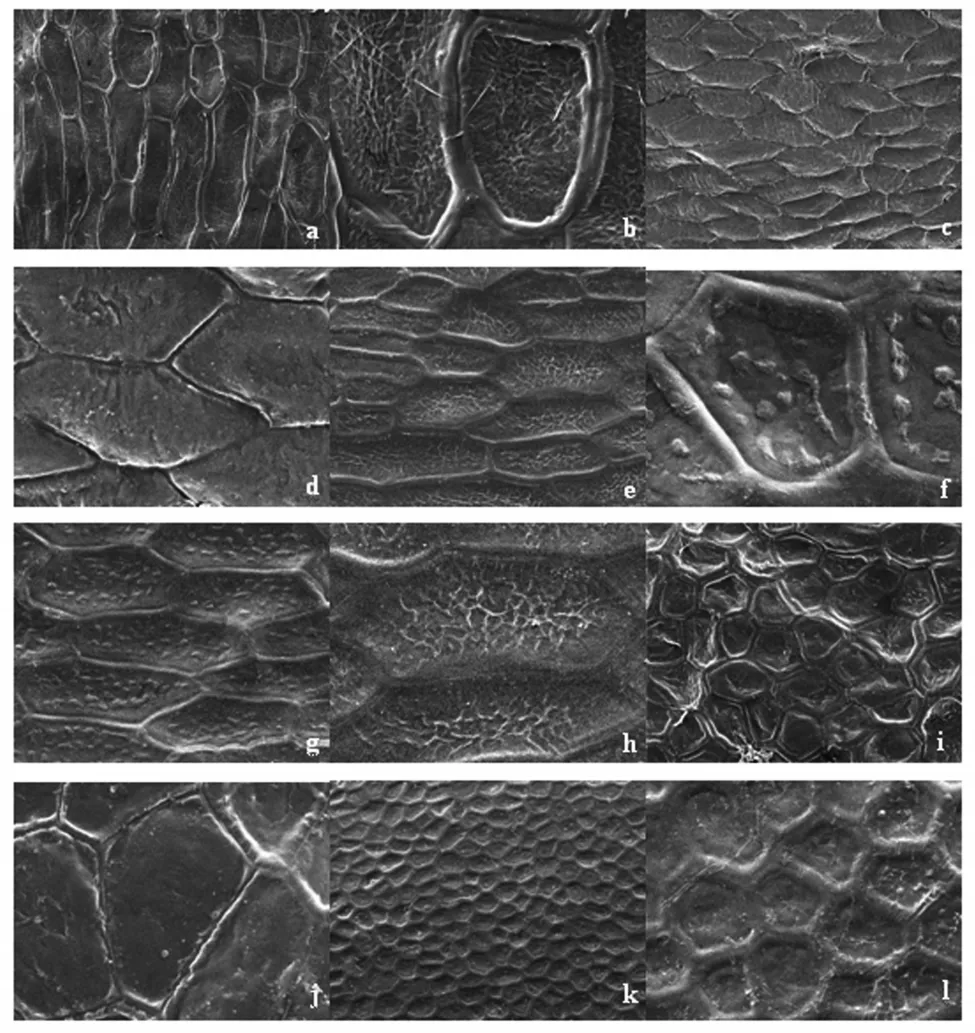
图1 扫描电镜下拍摄的种皮超显微结构(300倍及1 000倍)
由表2可以看出,溪荪、北陵、黄菖蒲、燕子花的子叶联结较长,而玉蝉花、马蔺的子叶联结较短。子叶鞘长度与子叶联结长度成正比,两者与舌状鞘长度则成反比。Boyd[17]指出,由于舌状鞘的存在,子叶联结常较短,也常伴随着子叶鞘(基鞘)的抑制。舌状鞘比子叶鞘能更好地保护胚芽。有舌状鞘的种类与无舌状鞘的种类相比,由于子叶联结短,子叶的维管束从吸器到达胚芽、胚根的联系更加直接而经济。因此可以推断,有舌状鞘的种类是较进化的类型。由此得出进化趋势为:6种鸢尾中马蔺是最进化的种类,其次是具有舌状鞘和基鞘的玉蝉花、黄菖蒲和燕子花,而溪荪、北陵则是相对原始的种类。这个结论与种皮电镜所得出的结论是一致的。
[1]Riley H P.A character analysis of colonies ofIris fulva,Iris HexagonaVar.Giganticaeruleaand natural hybrids[J].American Journal of Botany,1938,25(10):712-724.
[2]Mitra J.Karyotype analysis of bearded Iris[J].Botanical Gazette,1956,117(4):265-293.
[3]黄苏珍,顾姻,贺善安.鸢尾属(IrisL.)植物的杂交育种及其同工酶分析[J].植物资源与环境,1996,5(4):38-41.
[4]江明,谢文申.香根鸢尾的组织培养和快速繁殖[J].园艺学报,1995,22(3):301-302.
[5]Balkwill K,Campbell-Young G.Taxonomic studies in Acanthaceae:testa microsculpturing in southern African species ofThunbergia[J].Botanical Journal of the Linnean Society,1999,131(3):301-325.
[6]Goldblatt P.Systematics,phylogeny and evolution ofDietes(Iridaceae)[J].Annals of the Missouri Botanical Garden,1981,68(1):132-153.
[7]Manning J C,Goldblatt P.Endothecium in iridaceae and its systematic implications[J].American Journal of Botany,1990,77(4):527-532.
[8]Kron P,Stewen S C.Variability in the expession of a rhizome architecture model in a natural population ofIris versicolor(Iridaceae)[J].American Journal of Botany,1994,81(9):1128-1138.
[9]董晓东,赵毓棠,谢航.国产鸢尾属尼泊尔鸢尾亚属的研究[J].植物研究,1998,18(2):149-151.
[10]赵毓棠.国产鸢尾属的研究[J].植物分类学报,1980,18(1):53-62.
[11]谢航.中国鸢尾属有关分类群的讨论及属下分类系统的修订[D].长春:东北师范大学,1996.
[12]叶能干,季强彪,廖海民,等.种子植物幼苗形态学[M].贵阳:贵州科学技术出版社,2002:69-87.
[13]Corner E J H.The seeds of dicotyledons[M].Cambridge:Cambridge Press,1976:161-166.
[14]孔红.甘肃萱草属种子微形态及其分类学意义[J].西北植物学报,2001,21(2):373-376.
[15]陈世龙,何廷农.双蝴蝶属和蔓龙胆属(龙胆科)种子表面特征的研究[J].西北植物学报,2002,22(1):37-42.
[16]马骥,王勋陵,赵松岭.骆驼蓬属种子微形态及其生态学与分类学意义[J].武汉植物学研究,1997,15(4):323-327.
[17]Boyd L.Monocotyledonous seeding:Morphological studies in the post seminal development of the embryo Trans[M].Proc Roy:Soc Edinb,1932:30-35.
猜你喜欢
花卉(2021年10期)2021-12-06
中老年保健(2021年7期)2021-08-22
甘肃科技(2020年20期)2020-04-13
文萃报·周五版(2019年44期)2019-09-10
浙江中医杂志(2019年5期)2019-01-06
农机化研究(2018年2期)2018-06-05
中成药(2017年10期)2017-11-16
测绘学报(2017年6期)2017-07-01
植物保护(2016年1期)2016-09-14
中国棉花加工(2015年3期)2015-01-25
