
刺槐开花传粉及交配方式1)
2012-06-13 06:20:32胡瑞阳李允菲袁存权
东北林业大学学报 2012年1期
孙 鹏 戴 丽 胡瑞阳 习 洋 李允菲 袁存权 孙 妍 李 云
(林木育种国家工程实验室(北京林业大学),北京,100083)
刺槐(Robinia pseudoacacia)属于雌雄同花植物,原生于北美洲,后被广泛引种到欧洲、亚洲等地。刺槐叶片含有大量粗蛋白,可作饲料。刺槐木质坚硬且具有发达的根系。刺槐根部有根瘤,有提高地力之效;刺槐具有很强的抗旱能力,能够适应石质山地的干旱环境。刺槐的花属于蝶形花,开花时香气四溢,并且可供食用[1]。此外,刺槐还是一种重要的蜜源植物。综上所述,刺槐具有很好的生态和经济价值。但刺槐的应用还存在一些亟待解决的问题,例如,有些刺槐无性系速生性好,但抗性差;有些抗性强,但干型差或生长慢;有些叶片粗蛋白含量高,但生物量低等[2]。目前,刺槐改良以选种为主,但其选种是建立在实生刺槐林自由授粉的基础上,变异程度低,选择偶然性大,人为控制能力差。如果能够采用控制授粉实现刺槐杂交育种,必将提高刺槐改良程度。然而,本实验室通过多年研究发现刺槐杂交时去雄操作技术要求高、难度大,杂交结实率非常低,特别是去雄和套袋操作对刺槐的授粉受精和坐果影响严重。Dini-papanastasi et al.[3]对刺槐杂交育种的报道也发现杂交结实率低的现象。杂交工作在开花期进行,包括去雄、套袋、授粉等对花展开的操作。另外,杂交的成功还有赖于外源花粉的亲和能力。鉴于此,详细了解刺槐的开花传粉及交配方式对于提高刺槐的杂交育种工作水平具有重要的意义。近年来,国内外对刺槐开花特性的研究还比较少,主要有郝晨等[4]和姜金仲等[5]对四倍体刺槐花器官变异的报道,而二倍体刺槐的有关研究还未见报道。本研究关于刺槐花部外形、花粉萌发、柱头可授性的研究对于杂交时去雄时机的选择,花粉采集时间和储存方式以及授粉时机的选择具有指导意义。此外,关于刺槐天然群体的交配方式目前看法仍不一致,Surles et al.[6]利用等位酶技术研究认为刺槐天然群体异交率高达80%以上,而原法宪[7]认为刺槐是高度自交的树种。另外,Lian et al[8]从9个刺槐无性系采集种子,发现这些种子是高度自交的。针对上述疑问,本研究从刺槐的开花特性和传粉生物学入手,以延庆实生刺槐林为试材,以期阐明刺槐天然状态下自交及异交的亲和能力。该研究结果将有助于刺槐杂交时亲本的选择,以及提高刺槐杂交结实率。
1 试验区概况
试验地选在北京市延庆县米家堡苗圃(N40°29.748'~ 40°30.276',E115°59.192'~ 115°59.883'),延庆位于北京城区西北70 km处,属大陆性季风气候,处于温带与中温带、半干旱与半湿润带的过渡地带。气候冬冷夏凉,年平均气温8℃,刺槐多在5月底6月初开花。延庆米家堡苗圃的刺槐林为人工刺槐实生林。林区面积约6 hm2。所选样树高6~7 m,地径12~15 cm,胸径8~10 cm,为开花结实盛期的成年植株。
2 材料与方法
开花生物学特性观察:选择生长正常、树体健壮的3株刺槐成年植株,于开花后1 d在选定的每株树上随机选取3个正常花序,利用游标卡尺测量花序长和花序直径,然后在每个花序上选择10朵花,测量小花长、小花直径、花柱长、柱头与花药的最长和最短距离。
花粉形态观察:采集开花前2 d、开花当天、开花后24 h、开花后48 h及开花后72 h的花,用FAA固定。然后经叔丁醇干燥,镀金,利用扫描电子显微镜观察花粉形态并照相。
花粉体外萌发试验:开花当天采集花粉,放在培养皿中,加硅胶使其干燥,并于常温保存。用TTC法[9]测定花粉保存 24、48、72 h 后的活力。
花粉体内萌发试验:首先在开花散粉前人工去雄套袋,然后在适授期授粉,于授粉后2、6、10、24、48 h采集花朵,用FAA固定,根据胡适宜[10]报道的方法染色压片后在荧光显微镜下观察花粉萌发及花粉管生长情况。本试验材料取自3棵生长状况良好的单株,每个单株设3次重复。
柱头可授性试验:于幼花期、将开花期(该时期为本文叫法,特指开花前1~2 d的时期,图1A,下同)、开花当天、开花后24 h、开花后48 h,以及开花后 72 h 采集花朵,参照 Suwan et al.[11]的方法,利用扫描电子显微镜观察柱头形态,判断其可授性。试验材料取自3棵生长状况良好的单株,每个单株设3次重复。
传粉方式检测:在盛花期选取3株间隔50 m的刺槐单株观察其传粉媒介。在每个单株4周8个方位布置涂布凡士林的载玻片各2枚,凡士林涂布面积为2.5 cm×5.0 cm,露置1 d后收回,镜检以风为媒介散布花粉的数量。刺槐花粉的鉴定以其花粉的大小和形状与单独采制的标准花粉对比计数。
在野外选择间隔50 m以上的3株生长状况良好的刺槐单株,每个单株选择10个花序,于开花后连续3 d,每天分6个时段:8:00—8:30、10:00—10:30、12:00— 12:30、14:00—14:30、16:00—16:30、18:00—18:30进行观察,分别记为时段1至时段6。同时,对刺槐植株上的访花昆虫进行观察并拍照,凡发现有昆虫访花并在花上停留者,就用乙烯瓶捕捉、杀死并鉴定。同时记录每植株上所有来访昆虫的访花次数,并计算访花频率,访花频率的计量方法为10个花序在30 min内被访问的次数。
杂交指数估算:按照李昆等[12]研究小桐子的标准进行花序直径、花朵大小和开花行为的测量及交配系统的评判。具体方法:花朵直径(D)D<1 mm记为0,1 mm≤D<2 mm 记为1,2 mm≤D<6 mm 记为2,D>6 mm记为3;花药开裂时间与柱头可授期之间的时间间隔,同时或雌蕊先熟记为0,雄蕊先熟记为1;柱头与花药的空间位置,同一高度记为0,空间分离记为1。
上述三者之和为OC,I值。评判标准为:OC,I为 0时,交配系统为闭花受精;OC,I=1时,交配系统为专性自交;OC,I为 2 时,交配系统为兼性自交;OC,I为 3时,交配系统为自交亲和,有时需要传粉者;OC,I为4时,交配系统为部分自交亲和,异交需要传粉者。
花粉量及P/O值(花粉数/胚珠数)的测定:随机选取10棵树,每棵树随机选取3朵花药尚未开裂的小花,取小花的全部花药,用 0.1 mol·L-1HCl溶液在60℃下水解去除药壁,制成2 mL花粉粒悬浮液,吸取5 μL于显微镜下观察统计花粉数量,重复取样观察5次,取均值再乘以400(2 mL=5 μL×400)则为单花花粉数。取子房在解剖镜下用解剖针划开心皮,观测并记录胚珠数。最后计算花粉与胚珠的比值。按 Cruden[13]的标准,P/O为 2.7 ~5.4时,其交配系统为闭花受精;P/O为18.1~39.0时,交配系统为专性自交;P/O为31.9~396.0时,交配系统为兼性自交;P/O为244.7~2588.0时,交配系统为兼性异交;P/O为2108.0~195525.0时,交配系统为专性异交。
授粉试验:选择4株生长状态良好、间隔50 m以上、上一年天然状态下结实率较高的刺槐个体,进行完全双列杂交。参考李昆等[12]描述的方法进行下述处理:天然授粉,是不套袋,不去雄,自由传粉,用于检测自然条件下的传粉情况;自交,对花进行套袋但不去雄,检测自交能力;杂交,套袋并且去雄,用不同植株的花粉进行异花授粉;去雄不套袋,自然条件下异花传粉,对花进行去雄但不套袋,使去雄后的花在天然状态下异花授粉;孤雌生殖,对花进行去雄并套袋,但不授粉,使子房在没有花粉的情况下生长发育;竞争授粉,对花进行套袋但不去雄,人工授以异株花粉,在排除自交不亲和以及去雄影响的前提下,观察套袋对坐果率的影响。每种交配组合选5个花序,每花序保留10朵小花。在授粉后1周、1个月和3个月分别统计各交配类型的坐果率。其中坐果率=(结实数/开花数)×100%。
试验所用的花除自然授粉和自交外,均在将开花期去雄。这一时期花瓣闭合,花药未开裂,柱头表面干净无花粉。去雄后套袋隔离。授粉4 d(4 d时间足够花粉管生长到子房完成受精;4 d后柱头可授性大大下降,新的花粉难以萌发;去袋太晚会影响花的正常发育)后将硫酸纸袋去掉。人工授粉时间应在去雄后2 d内,否则柱头干燥,且可授性差。在试验期间,对套袋的花朵经常进行检查,并揭袋换气,防止花朵霉坏,换气时严格防止昆虫在花序附近活动。在授粉后1周(6月9日)、授粉后1个月(7月15日)以及授粉后4个月(10月20日)分别统计各交配组合的坐果率。由于4株树中的1株在授粉后遭到了破坏,因此,只统计其余3株的坐果率。
3 结果与分析
3.1 花朵表型性状
刺槐花为总状花序、蝶形花、两性花、雌雄同花,雄蕊10枚围绕在雌蕊周围。各雄蕊与柱头距离并不一致,距离最短的1.5 mm左右,距离最长的为4.5 mm左右。有些单株花萼为绿色,有些单株花萼略带红色(图1A)。花冠为白色,开花时香气浓郁。花的各项表型性状,即:花序长、花序直径、小花长、小花直径、花柱长、柱头与花药的最长距离、柱头与花药的最短距离分别为(112.53±6.71)、(33.67±1.95)、(24.37±0.20)、(11.49±0.30)、(7.05±0.06)、(4.53±0.11)、(1.48±0.08)mm。

图1 刺槐花形态、访花昆虫以及花粉管在花柱中生长
3.2 花粉形态观察
刺槐花粉近椭圆形,直径15~30 μm,具3个萌发沟,萌发沟处着色较深(图2A)。花粉外壁粗糙具有深刻而清晰的一级网状纹饰和微小的颗粒状次级雕纹且富有黏性(图2B),是虫媒花花粉粒的主要形态特征。因此,刺槐的花粉在形状、大小和外壁表面纹饰等方面的特征符合虫媒花的主要特征,花粉的形态特征与传粉的方式是相适应的,刺槐花粉的形态特征有利于花粉粘附在昆虫身体上传播。开花当天落在柱头上的花粉有些已经开始长出花粉管,花粉外形依然饱满;开花24 h后柱头上的花粉大量长出花粉管,其外形开始皱缩(图2F);开花48 h后柱头上的花粉进一步皱缩,大部分花粉已完成萌发。
3.3 花粉萌发
TTC法表明花粉在收集后24 h具有活力的占87.67%,收集后48 h具有活力的占23.78%,收集后72 h具有活力的占16.83%。
花粉管体内生长试验是将花粉授到提前去雄(去雄到授粉不超过2 d)的花的柱头上,然后每隔一定时间观察花粉管生长情况。结果表明,授粉后2 h花粉已经开始萌发(图1C),授粉后6 h已有少量花粉管生长到花柱基部(图1D),授粉后24 h花粉管完全生长到子房内(图1E),授粉后48 h花柱内的花粉管依然清晰可见(图1F)。
3.4 柱头可授性
幼花期的柱头干燥且缺乏黏液,可授性较差(图2C);将开花期的柱头上黏液较多,而且几乎没有附着花粉,这一时期是去雄的好时机(图2D);开花当天柱头黏液很多,表现出最佳的可授性,柱头上已沾染大量的花粉粒,这一时期去雄已晚,但却是授粉的最佳时期(图2E);开花后24 h柱头上依然有大量黏液,附着的花粉粒已经萌发出花粉管(图2F);开花后48 h柱头上黏液明显减少,有些柱头开始解体,形状变得不规则,说明柱头可授性开始下降(图2G);开花后72 h柱头上黏液已经很少,柱头进一步降解,形状更加不规则,此时柱头可授性已经很差(图2H)。综上所述,去雄的时机应选在将开花期,然后,必须在去雄后2d内完成授粉,这样既可以防止柱头被非目标花粉污染,又可以使得柱头与目标花粉亲和性最佳。另外,需要指出的是,利用扫描电镜观察时,柱头已经过干燥处理,但是黏液中含有的糖、氨基酸类物质会使柱头表面显得模糊,观察时看不清柱头表面乳突结构,据此可以判断柱头表面是否具有黏液。

图2 花粉及柱头表型的扫描电镜观察
3.5 刺槐传粉媒介分析
3.5.1 风媒传粉观测
用重力载玻片法检测到刺槐花粉风力传播能力极其有限,每个单株载玻片上粘黏花粉数如表1。利用显微镜观察载玻片,几乎所有视野都观察不到花粉,只有极个别视野内分布有高度集中的花粉。如果把柱头看成半球形,则其半径最长0.05 cm,表面积最大0.015 7 cm2(S=4πr2)。如表1,每平方厘米载玻片露置1 d后平均粘黏花粉量为7.50粒,则柱头露置1 d平均粘黏花粉为0.1粒。这样的效率说明风媒对刺槐传粉的贡献微乎其微。
3.5.2 虫媒传粉观测
刺槐的花萼有的呈红色,有的呈绿色,花冠呈白色,在绿叶衬托下非常显眼。刺槐花香气浓郁,对访花者极具吸引力。刺槐种群记录到的访花者可分为3类:分别为膜翅目的蜂类和蚁类,以及双翅目的蝇类。其中蜂类的蜜蜂为刺槐的最主要访花昆虫。蜜蜂访花时身体位于旗瓣和龙骨瓣之间,头部伸向花萼基部蜜腺处,后面的触手蹬在龙骨瓣上。雌雄蕊此时失去龙骨瓣包裹,与蜜蜂腹部产生接触,这使得花粉粘附在蜜蜂腹部及触手上,而柱头则接触到蜜蜂携带的异源花粉,从而完成虫媒传粉过程(图1B)。从表2可以看出晴好天气有利于蜜蜂访花,阴雨和大风天会妨碍昆虫的访花行为。蝇类和蚁类访花频率极低,且很少直接与花药柱头接触,因此,与蜜蜂相比,其传粉作用非常微小。综上所述,刺槐的传粉方式主要是虫媒传粉。
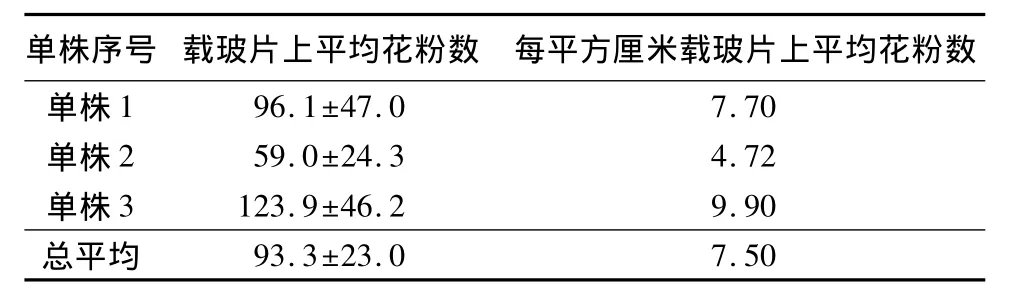
表1 载玻片露置1 d后粘黏花粉粒数 粒
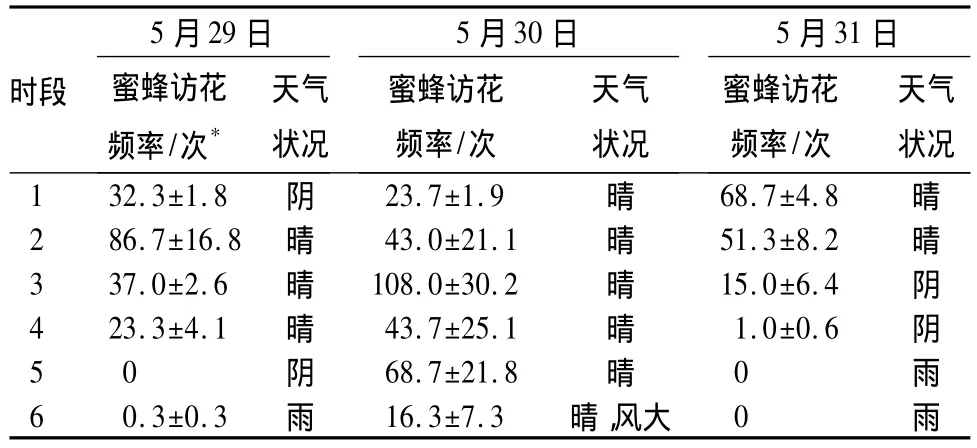
表2 蜜蜂访花频率统计
3.6 交配方式评判
3.6.1 刺槐杂交指数
刺槐花朵直径(11.49±0.30)mm。刺槐柱头可授性从开花前2 d延续到开花后2 d,这一时期也是花粉成熟散粉的时期,因此,可以认为雌雄蕊成熟时间相同。雌雄蕊在空间上有所分离,雌蕊比雄蕊高1~5 mm。以上几点使得刺槐杂交指数达到4。根据 Dafni[14]和李昆等[12]的标准,该树种的交配系统为部分自交亲和,异交需要传粉者(表3)。

表3 刺槐杂交指数
3.6.2 刺槐的花粉—胚珠比(P/O)
刺槐单花的平均胚珠数为(20.37±0.32),单花的平均花粉数为(28 071.67±879.47),花粉—胚珠比(P/O)约为 1 378.09。按 Cruden[13]的标准,刺槐的交配方式为兼性异交。
3.6.3 控制授粉试验坐果率统计
各交配组合在授粉1周、1个月、4个月后结实率统计如表4。
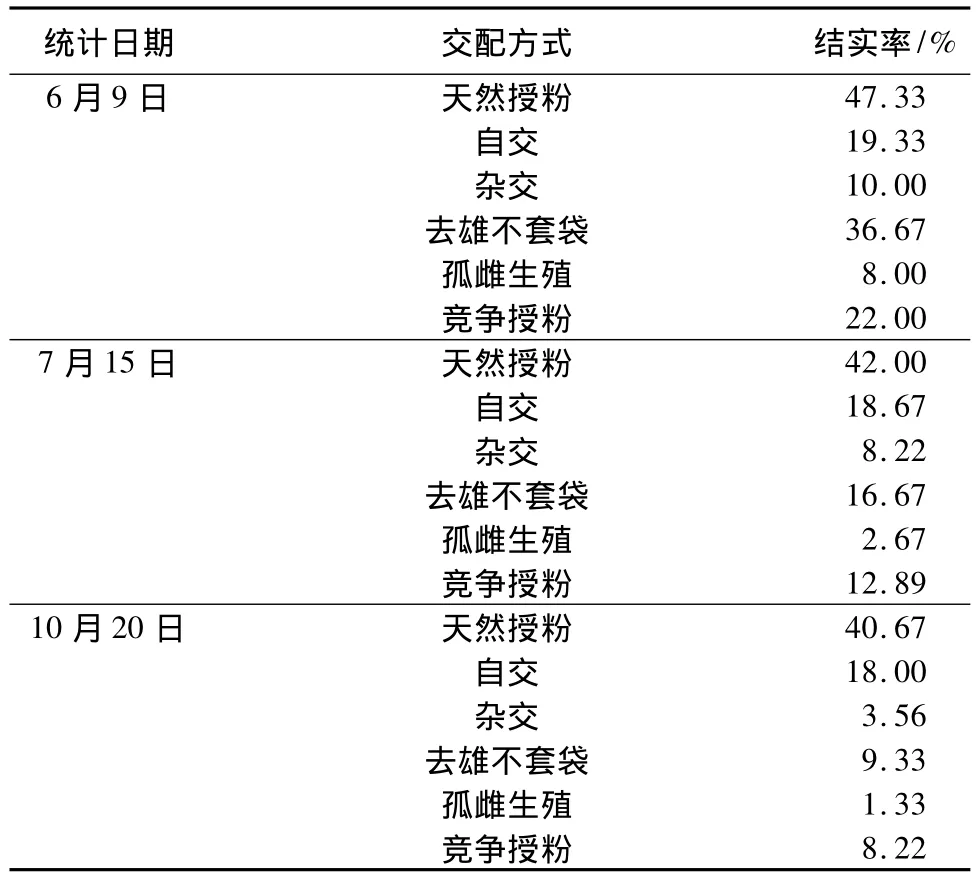
表4 控制授粉结实率统计
刺槐的果实到九十月份才能完全成熟。从10月20日的统计结果看,天然授粉结实率(40.67%)远远高于其他交配方式,自交结实率为18.00%,而杂交结实率为3.56%,去雄不套袋结实率为9.33%,孤雌生殖结实率为1.33%,竞争授粉结实率为8.22%。从上述3个时期的统计结果看,各交配组合在授粉后第1周内落花率最高,此后直到果实成熟的几个月时间内,落花率大大下降,坐果率变化相对较小。
4 结论与讨论
何亚平等[15]认为自然界绝对自交或异交的植物种类很少,大多数为两者兼而有之,即为混合交配的交配模式。植物近交会导致后代衰退,远交易受环境影响。自然选择使刺槐形成了以异交为主,部分自交的混合交配系统。Surles et al.[6]以多地刺槐天然大群体为研究对象,利用等位酶标记研究发现,天然状态下刺槐80%以上为异交,这与本文观点一致。而原法宪[7]认为刺槐主要是以自交为主的树种,其主要理由是刺槐在开花前,花药就已经开裂,从而使得柱头上沾满了花粉,这些花粉必然是自花花粉。本试验通过3年观察也发现刺槐在开花前柱头上就已经沾满了花粉这一现象。既然如此,那么为何 Surles et al.[6]和本文的研究结果都认为刺槐是以异交为主的树种呢?Hasegawa et al.[16]报道指出板栗在传粉阶段,自花传粉占到90.2%,而到种子成熟时,自花传粉所得种子仅仅占0.3%,因此,他指出自交不亲和性强烈抑制了自花传粉种子的生长和发育。Kristina et al.[17]在研究Platypodium elegans时也发现,从授粉结束到种子成熟,再到幼苗长出的过程中,自交后代个体所占的比例不断下降,尤其从种子成熟以后到幼苗长出这段时间下降更快。此外,他还指出自交所得种子的质量明显比异交得到的种子轻。据此,猜测刺槐也存在这一机制——从授粉到种子成熟再到幼苗长出的过程,自交后代所占比例逐渐下降,最后存活的个体就主要是异交的后代。关于这一点还需要后续相关的种子成熟、萌发及幼苗生长的对比试验进行验证。Lian et al.[8]利用SSR分子标记对从9棵单株上采集的种子进行亲本鉴定后认为,这些种子大部分是自交个体。但该试验样本取自无性系林分,样本量小,仅有9个无性系,不足以说明刺槐天然群体的交配类型。在无性系林分中,无性系之间的授粉会受其周围的无性系个体影响,虽然是异株传粉,但会有一定程度的相同基因型之间的杂交。因此,该试验结果不足以代表刺槐天然大群体的异交率。
载玻片粘黏花粉试验计算得出每平方厘米载玻片上平均黏着花粉数为7.50粒,这一结果只有假设花粉平均分布在载玻片上才可靠。事实上,花粉在载玻片上的绝大部分区域并没有被观察到,只有极个别的区域分布有高度集中的花粉粒。如果是风媒传粉,那么分布在载玻片上的花粉就应该趋于分散。高度集中的花粉粒可能是因为载玻片与雄蕊在风的作用下直接接触造成的,具体原因还需进一步观察。根据以上推测,认为风媒对刺槐的传粉贡献可以忽略不计,刺槐属于虫媒传粉植物。观察发现,蜜蜂采蜜时经常就近访问同一株树的不同花,这种同株异花授粉依然属于自交,因为同株花粉基因型相同。此外,由于蜜蜂就近采蜜,杂交的花粉来源也以邻近的单株为主,这会使得杂交后代基因型与邻近植株相近,变异幅度有限。
花粉萌发和柱头可授性试验表明开花当天散粉量最大,花粉收集后放入加硅胶的培养皿,常温下保存24 h后萌发率可达80%以上,常温下保存48 h后萌发率不到30%。野外杂交试验中,花粉采集后要尽快授粉,否则花粉活力会大大下降。柱头可授性试验表明,柱头适授期从开花前2 d持续到开花后2 d。早于这一时期,柱头干燥,不易粘黏花粉。晚于这一时期,柱头开始降解,形状不规则,缺乏粘液,不利于花粉萌发。杂交试验去雄时机应尽可能地晚,但是一定要在散粉前,这样既可以把去雄对花发育的影响降到最小,又能保证柱头不被自身花粉沾染。去雄后应尽快(2 d内)完成授粉,否则花粉活力和柱头可授性都将大大下降。
文中的授粉试验显示,刺槐天然生长结实率为4 0.66%,自交结实率为18%,杂交结实率为3.56%,孤雌生殖结实率为1.33%。本试验去雄套袋操作虽然严格防止自身和外界花粉污染,但是因为没有对后代种子进行染色体倍性分析,所以还不能肯定刺槐存在孤雌生殖的现象。
另外,从以上数据看出,刺槐天然生长也存在落花落果现象,这和很多果树的生理落花落果现象一致,但去雄杂交后刺槐落花落果率急剧增加,这就不是普通意义的生理落花落果,而一定是与去雄或套袋的人工操作有关。杂交过程需要去雄、套袋、授粉(本试验杂交所用的花粉已被证明是有活性的)三步人工操作,而自交只需要套袋一步人工操作。现假设自交和杂交坐果率低的原因只是套袋(套袋使花处于一个相对封闭的环境,花生长环境中的温度会升高,光照会减少,风吹时袋对花有可能造成机械损伤作用)引起的,而与去雄无关,那么根据Surles et al.[6]的试验以及文中观点,刺槐天然状态下属于高度异交树种,外来花粉更有利于结实,由此推断刺槐杂交结实率应高于自交,但是从文中杂交试验结果看,杂交结实率远远低于自交,那么在保证杂交所授花粉有活力的前提下,可以确定去雄是造成杂交结实率极低的最主要原因,此外,去雄不套袋花的结实率也低于天然生长和自交,从而进一步断定去雄是影响花发育和结实率的主要因素。
按照这一推论,人工杂交所得结实率并不能反应自然状态下刺槐真实的异交结实率。从Dini-papanastasi et al.[3]的报道中也可发现刺槐杂交结实率低的现象。前人在兰属[18]和石竹属[19]以及豇豆[20]等植物中发现去雄会加速花衰老。在酸樱桃中已有去雄杂交会降低结实率的报道[21]。Hedhly et al.[22]研究甜樱桃发现,去雄杂交后落花落果率明显上升,并且去雄杂交后雌蕊在随后9d内重量增长比天然个体的慢,去雄杂交后10 d内胚珠退化率比天然个体高,并推测去雄后花器官乙烯含量上升是造成以上现象的重要原因之一。此外,从上述3个时期的统计结果看,各交配组合在授粉后第1周内落花率最高,直到果实成熟的三四个月时间内,落花率大大下降,坐果率变化相对较小。因此,研究去雄授粉后1周内花器官生理生化变化,对于了解去雄如何影响花发育及结实率的机理至关重要。综上所述,去雄对刺槐花发育以及坐果率的影响是值得深入研究的问题。
通过本研究发现刺槐是以异交为主的树种,因此,杂交育种中使用外源花粉进行授粉是可行的。另外,还发现了刺槐人工杂交结实率低的现象,如果该问题能够得到解决,那么将极大地提升刺槐的杂交育种工作水平。
[1]Keeler H L.Our native trees and how to identify them[M].New York:Charles Scriber’s Sons,1900:97-102.
[2]张国君.饲料型刺槐优良无性系选育及其栽培利用研究[D].北京:北京林业大学,2010:15-23.
[3]Dini-papanastasi O,Filippos A.Aravanopoulosartificial hybridization betweenRobinia pseudoacaciaL.andR.pseudoacaciavar.monophyllaCarr[J].Forestry,2008,81(1):91-101.
[4]郝晨,李云,姜金仲,等.四倍体刺槐大小孢子发育时期与花器形态的相关性[J].核农学报,2006,20(4):292-295.
[5]姜金仲,郝晨,李云.四倍体刺槐花器原基分化及其成熟表型变异[J].林业科学,2008,44(6):34-41.
[6]Surles S E,Hamrick J L,Bongarten B C.Mating systems in openpollinated families of black locust(Robinia pseudoacacia)[J].Silvae Genetica,1990,29(1):35-40.
[7]原法宪.刺槐开花生物学特性观察[J].山西林业科技,1987(3):1-10.
[8]Lian C,Oishi R,Miyashita N,et al.High somatic instability of a microsatellite locus in a clonal tree,Robinia pseudoacacia[J].Theor Appl Genet,2004,108:836-841.
[9]胡适宜.植物胚胎学实验方法(一):花粉生活力的测定[J].植物学通报,1993,10(2):60-62.
[10]胡适宜.植物胚胎学实验方法(五):检查花粉在柱头上萌发和花粉管在花柱中生长的制片法[J].植物学通报,1994,11:58-60.
[11]Suwan T,John N O.Floral biology,pollination,pistil receptivity,and pollen tube growth of teak(Tectona grandisLinn)[J].Annals of Botany,1997,79:227-241.
[12]李昆,尹伟伦,罗长维.小桐子繁育系统与传粉生态学研究[J].林业科学研究,2007,20(6):775-781.
[13]Cruden R W.Pollen ovule ratios:A conservative indicator of breeding systems in flowering plants[J].Evolution,1977(31):32-36.
[14]Dafni A.Pollination Ecology[M].Oxford:Oxford University Press,1992:59-89.
[15]何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151-163.
[16]Hasegawa Y,Suyama Y,Seiwa K.Pollen donor composition during the early phases of reproduction revealed by DNA genotyping of pollen grains and seeds ofCastanea crenata[J].New Phytologist,2009,182:994-1002.
[17]Kristina M H,Hamrick J L.Viability selection at three early life stages of the tropical tree,platypodium elegans(Fabaceae,papilionoideae)[J].Evolution,2003,57(3):518-526.
[18]Woltering E J,Harren F.Role of rostellum desiccation in emasculation-induced phenomena in orchidfiowers[J].J Exp Bot,1989,40:907-912.
[19]Woltering E J,Somhorst D,de Beer C A.Roles of ethylene production and sensitivity in senescence of carnationfiower(Dianthus caryophyllus)cultivars white sim,chinera and epomeo[J].J Plant Physiol,1993,141:329-335.
[20]刘先斌,宿秀丽,温海霞.提高长豇豆“去雄套袋杂交法”结实率的关键技术[J].作物杂志,2009(2):101-103.
[21]Brown S K,Iezzoni A F,Fogle H W,et al.Fruit breeding:VolⅠ tree and tropical fruits[M].New York:John Wiley& Sons,1996:213-255.
[22]Hedhly A,Hormaza J I,Herrero M.Flower emasculation accelerates ovule degeneration and reduces fruit set in sweet cherry[J].Scientia Horticulturae,2009,119(4):455-457.
猜你喜欢
中国科学探险(2022年8期)2022-12-31 07:27:54
疯狂英语·新悦读(2022年12期)2022-03-08 02:37:12
现代园艺(2020年2期)2020-12-22 05:57:04
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25 01:40:26
小溪流(故事作文)(2018年10期)2018-12-28 10:45:08
铜仁学院学报(2018年7期)2018-09-08 08:50:40
现代园艺(2018年2期)2018-03-15 08:00:12
现代园艺(2017年22期)2018-01-19 05:06:57
中国稻米(2017年6期)2017-12-28 02:13:01
山东林业科技(2016年5期)2016-07-05 00:43:00
