
网球运动员击球线路预判能力及ERP特征研究
2012-05-23 09:04:54周成林
体育科学 2012年12期
张 怡,周成林
体育运动中的知觉预测是建立在背景知识基础上对可能发生事件的内隐期望,是快速、准确执行动作反应的重要依据,是运动决策的前提。网球作为高策略性的开放性运动项目,对运动员的知觉认知技能要求很高,是研究知觉预测的典型项目。在比赛的攻防对抗中快速搜索对手击球信息,对来球的力量、速度、线路、落点等做出准确预期对于网球运动员赢得每一分起决定性作用。
关于网球运动员的知觉认知技能,国内、外学者已开展了大量研究。Jones等(1978)运用时间阻断技术,要求被试在不同时间预测发球落点,结果发现,高水平网球运动员在1/8s时间点的预测能力显著优于新手[43]。Pherson(2000)研究女子网球运动员的执行反应能力发现,专家对动作的预期反应是基于精细的、复杂的动作和对现实情境的配置,而新手缺乏这种记忆结构[33]。Shim(2005)研究了专家和新手网球练习者利用对方运动信息预测击球类型和方向的能力,发现两组运动员的正确预期均大于随机水平,专家运动员对现场(馆)来球的反应速度明显快于应对发球机的来球[37]。Raoul Huys(2009)、Farrow(2012)等对不同网球运动员发球落点的预判能力进行了研究。结果发现,当遮蔽对手不同身体部位时,技术熟练的运动员能以全局性的方式提取信息,比低技能运动员提取的信息相关度更高,因此,判断准确率高[18]。不同年龄发展阶段的专业网球运动员对现场(馆)情境视频发球落点的预判能力的不同主要表现在年长的运动员更善于利用对手击球前的运动学信息,反应速度快,而年轻运动员则更多地从球飞行的过程中提取信息做出反应[22]。Smeeton(2011)和Loffing(2011)等研究了高水平网球运动员和新手运动员观看不同呈现方式的实验材料(如视频呈现、光点呈现)预测对手发球方向的能力。结果发现,被试的预测绩效都受到了呈现方式的影响,虽然对光点呈现的判断结果差于视频,但高水平网球运动员的预测能力在各种条件下均优于新手,与运动场(馆)景相关的信息是不同水平运动员的反应行为出现差异的重要因素[23,38]。以上研究多从行为角度或辅助眼动记录技术揭示出不同水平运动员的差异。相关研究结果均表明,基于有限的先前信息预测对手行为的能力是区别高水平网球运动员的重要标志。
近些年,国内、外学者们试图运用时间分辨率很高的事件相关电位(ERP)技术检测知觉预测过程中大脑皮层的电生理活动。如Jin H(2011)比较了职业羽毛球运动员和非运动员观看不同难度羽毛球比赛视频剪辑并预测球落点的行为学和ERP特征,发现职业运动员的判断准确率显著高于非运动员,并表现出较好的行动预期。同时,职业运动员的P300波幅大于非运动员,且潜伏期短[19]。王小春等(2012)以时间序列遮蔽为手段,借助事件相关电位技术研究揭示出网球专家的预判准确性与反应速度具有显著优势,网球专家诱发的N1波幅显著大于经验组和新手组,且受时间序列影响显著,专家诱发PSW的时间显著早于经验组[8]等。以上研究揭示了运动专家知觉预测过程中各时间阶段的认知加工特征及认知优势的内在机制。
已有关于网球知觉预测的研究多以发球或某一拍球的图片或视频为实验材料,但在实际比赛过程中,每一拍的击球策略会因双方攻防对抗的表现而不同,因此,运动中知觉预测的研究不能脱离特定的运动情境。“相持阶段”(即第4拍以后的攻防对抗阶段)作为网球比赛中重要的竞技单元,是继“发球”和“接发球”之后得失分的关键环节,其最后一拍的得分依赖于运动员对相持过程中每一拍球的感知、判断、分析和比较。
因此,本研究以网球比赛相持阶段为背景,采用“专家-中间水平-新手”研究范式,结合事件相关电位技术,考查不同水平网球运动员对击球线路的预判绩效,探索优秀网球运动员击球线路预判过程的信息加工特征,为检测运动员知觉预测能力提供方法支持,为改进训练方法提供理论依据。研究假设为,优秀网球运动员预判击球线路时能较好地把握关键线索出现的时机,快速提取存储在大脑中的信息,表现出认知优势,形成既快又准的决策模式。
1 研究对象与方法
1.1 研究对象
43名被试按照运动等级分为专家组、二级组、新手组。专家组为江苏省网球队及中国网球学院运动员,运动等级均为一级以上(其中1人获全国网球大奖赛女子单打冠军,1人获两届全运会冠军,5人获城运会冠军),代表了国内最高水平;二级组为上海体育学院网球专项运动员和江苏省省队运动员,均达到国家二级水平;新手组为上海体育学院体育教育专业网球专项大一学生,尚无运动等级。
所有被试健康状况良好,无遗传病史,无脑部损伤、无精神和神经系统疾病,视力或矫正视力正常,无色盲色弱,均为右利手,自愿参加本实验(表1)。

表1 本研究被试基本信息一览表Table 1 Information of Participants
1.2 研究方法
1.2.1 实验设计
实验采用3(组别:专家组、二级组、新手组)×3(时间阻断点:Time1、Time2、Time3)两因素混合设计。自变量为组别、实验材料的时间阻断点,其中,组别为被试间变量,时间阻断点为被试内变量。因变量分别为反应时、正确率、ERP峰波幅和潜伏期。
1.2.2. 实验材料
实验刺激材料来自于2008—2010年温布尔登网球公开赛男子单打比赛高清视频。根据实验目的截取“相持段”得分球最后三拍的比赛片段,分别在对手最后一拍触球前 80ms(Time1)、击球瞬 间(Time2)、击球后 80ms(Time3)终止,片段长度统一为3 000ms。经部分教练员、运动员预评估后,最终筛选出144段作为正式实验材料(avi格式,25帧/s)。所有材料由E-prime软件编程,随机呈现。
1.2.3 实验流程
新手组和部分二级组被试的实验在上海体育学院运动心理研究中心脑电实验室进行,专家组和其余二级组被试的实验在南京体育学院心理实验中心进行,实验室设置标准一致,无噪音干扰,光线暗淡柔和。实验前告知被试全部实验流程,填写知情同意书和被试基本情况调查表,清洗头皮角质并吹干,关闭手机等通讯工具,防止电磁干扰。实验刺激材料呈现于19in.DELL液晶显示器,主机操作系统为 WindowsXP,CPU主频3.0Hz,刷新频率100 Hz,调整分辨率至1 280×720。被试端坐于实验台前,调整座椅高度,使之平视时视线汇聚于观测屏幕的中心,眼睛距屏幕60cm,下巴置于U型托上以稳定头部。双臂自然放置于实验台上,左右手食指分别置于数字键盘的1、3键。完成准备工作后,主试再次详细介绍实验流程后,由被试独立在操作间完成实验任务。全部流程由实验人员监控。具体实验流程如下(图1):

图1 本研究实验流程图Figure 1. Experimental Flow Graph
首先,呈现指导语:“下列视频将呈现比赛相持阶段的最后三拍,最后一拍为得分球,请仔细观察视频,并对对手最后一拍的击球线路做出预判,击向左边场(馆)地:按数字键1,击向右边场(馆)地:按数字键3,要求判断既快又准”,练习10个试次,以熟悉实验流程和按键,每个试次后给予反应时和正误反馈,练习结束后静息1min,按任意键开始正式实验,正式实验过程中不提供反馈信息。
被试在视频播放的3 000ms及之后的2 000ms时间内均可做出按键反应,反应时以视频开始播放为零点计时,反应时小于2 000ms或在规定时间内未按键均视为错误。要求被试在实验过程中集中注意力并尽量减少眨眼。
1.2.4 数据采集与处理
行为数据由E-prime 2.0软件采集,并通过E-DataAid对数据进行筛选、合并及预处理,剔除平均值加减3倍标准差之外的异常值,分析各组在不同时间定格点的判断正确率和反应时。
ERP数据由德国Brain Product公司生产的ERP记录分析系统进行记录与离线(off-line)分析,采用 Ag/AgCL电极,按国际10~20系统扩展的64导电极帽记录EEG,以双侧乳突为参考电极,水平眼电(HEOG)置于右眼外侧1cm处,垂直眼电(VEOG)安置于左眼框上方1cm处,实验时在各电极注入导电膏,保证头皮与电极之间阻抗降至5KΩ以下。脑电信号(EEG)经放大器放大后连续记录,采样频率为1 000Hz/导。
对EEG数据进行离线分析时,以双耳乳突的平均电位为参考,去除眨眼伪迹,充分排除噪音干扰和其他伪迹,30Hz低通滤波(24dB/oct),去除50Hz市电干扰,波幅大于±100μV者视为伪迹自动剔除。根据本实验目的,观察所有被试判断正确的叠加平均后的波形,根据电极在头皮的分布关系及实验目的,最终确定对以下3段时程进行分析:
第1段:以视频开始播放(onset)为0点,分析视频开始后0~500ms的时程内的早期ERP成分,并对波峰和潜伏期进行统计分析(图1a,epoch1);
第2段:以视频开始(onset)为0点,将500~3 000ms划分为多个小的时间窗口,观察晚期成分,并对各时间窗口的平均波幅进行统计分析(图1a,epoch2);
第3段:以视频结束(offset)为0点,根据行为学数据以及EEG记录结果,选取offset后0~1 000ms,对其成分的峰值和潜伏期进行统计分析(图1b,epoch3)。
所有分段均以视频开始播放前的600ms(-600~0 ms)为基线进行基线校正。
所有数据预处理后,采用SPSS for Windows 18.0软件进行重复测量方差分析,对不满足球形检验的统计量采用Greenhouse-Geisser法矫正自由度和P值,事后比较采用LSD法,本研究只对有统计学意义的显著性差异结果进行讨论分析。
2 研究结果
2.1 行为结果
图3为各组运动员判断击球线路的反应时、正确率及差异分析。以时间阻断点作为被试内变量,组别作为被试间变量,分别以反应时和正确率为因变量进行重复测量。方差分析结果显示,被试反应时的组别主效应不显著[F(2,40)=2.294,P=0.114,ηp2=0.103],时间阻断点主效应 非 常 显 著 [F(1.585,63.398)=66.345,P <0.001,ηp2=0.624],即被试反应时随时间阻断点的推后而缩短。组别×时间点交互作用不显著[F(3.170,63,396)=2.147,P=0.100,ηp2=0.097]。
事后分析发现,专家组判断击球线路的反应时显著快于二级组(P<0.05)。进一步通过单因素方差分析进行组内两两比较,结果显示,在对手击球前80ms的时间点(Time1),专家的反应时显著低于二级组(P<0.05);在击球瞬间和击球后80ms的时间点,专家预判的反应时与二级组和新手组相比无显著性差异(P>0.05)。
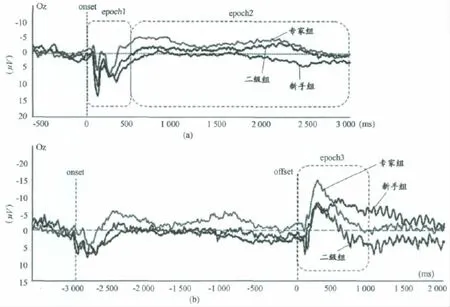
图2 本研究ERP数据分析分段示意图(以Oz为例)Figure 2. Diagram of ERP Epoch(Take Oz for example)
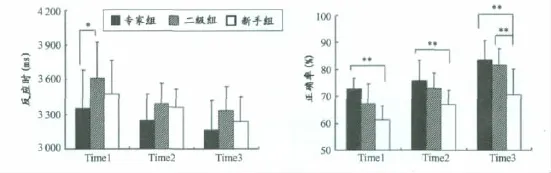
图3 本研究被试判断击球线路的反应时和正确率示意图Figure 3. The Reaction Time and Accuracy Rate in Ball Routes Anticipation
对被试在不同时间点判断击球线路的正确率进行重复测量方差分析,结果显示,时间阻断点主效应显著[F(2,80)=36.859,P<0.01,ηp2=0.480],组别主效应显著[F(2,40)=24.659,P<0.01,ηp2=0.552],时间和组别交互作用不显著[F(2,40)=1.030,P>0.05,ηp2=0.049]。进一步进行组间单因素方差分析显示,专家运动员在各时间点判断击球线路的正确率均显著高于新手(P<0.01),在对方球出手后80ms的时间点,专家和二级组的判断正确率均显著高于新手(P<0.01)。通过对专家组男、女运动员的反应时和判断准确率进行统计分析,未发现明显的性别差异。
以上结果提示,优秀运动员只在最早的时间阻断点表现出明显的反应速度优势,并在对手击球信息呈现尚不完整时能有效地利用击球前的关键信息,预判准确率明显高于新手。
2.2 脑电记录结果
脑电数据采集过程中,新手组和二级组各有2名被试由于记录过程中噪音等伪迹干扰过多,视为无效数据剔除。实际记录到有效ERP数据的样本量为:新手组13人、二级组11人、专家组15人。统计分析的3段时程反映了被试执行任务时认知加工的动态过程。Epoch1早期成分揭示了被试接受刺激并对任务进行初步识别的心理过程,Epoch2晚期慢波表示被试对实验材料的感觉、知觉、记忆、分析、判断等信息加工过程,Epoch3是基于对实验材料全过程的感知分析做出决策的主要阶段。各段统计结果如下:
2.2.1 早期 ERP成分
观察ERP总平均图显示,3组被试在实验材料呈现早期,大脑皮层各部位有不同程度的激活水平,诱发出明显的P1、N1、N2、P300成分,其特点如下:
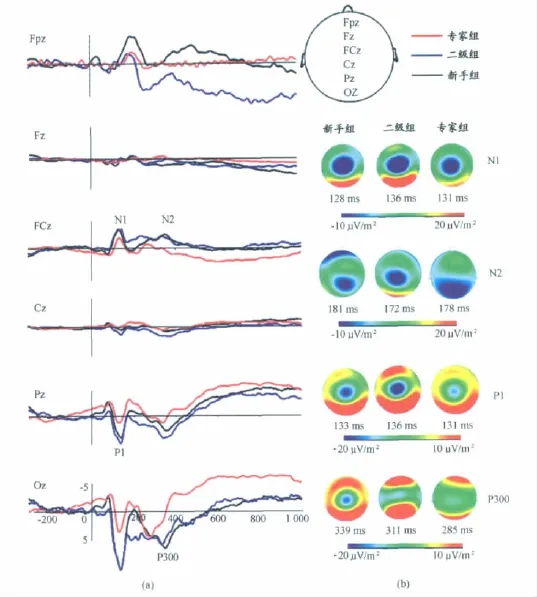
图4 代表性电极点ERP波形图及电流密度图Figure 4. ERP Oscillogram and CSD Maps of Several Representative Electrode
2.2.1.1 P1成分
本实验刺激呈现后在各脑区出现的第一个正走向的成分,潜伏期在80~140ms之间,我们认定其为P1,在枕区波幅达到最大(图4a可见Oz点P1波幅最大,图4b红色区域表示枕区激活最明显)。观察所有被试叠加后的ERP波形,根据头皮分布与电极位置之间的关系选取顶枕区中线及两侧共9个代表性电极(Pz、Oz、POz、PO3、PO4、P5、P6、PO7、PO8),分别以波峰和潜伏期为因变量,进行3(组别)×9(电极点)两因素重复测量方差分析,结果显示:波 峰在 各 电 极 点 主 效 应 显 著 [F(4.106,147.803)=27.22,P<0.01,ηp2=0.431],其中,枕区(PO8、Oz、PO7)峰值最大。组别主效应显 著[F(2,36)=4.542,P<0.05,ηp2=0.201],电 极点 和 组 别 之 间 无 交 互 作 用 [F(8.211,147.803)=1.146,P=0.113,ηp2=0.075]。事后检验显示,各组 P1波的峰值大小顺序为:二级组>新手组>专家组,其中,二级组(11.701±1.563μV)的 P1波幅显著高于专家组(5.705±1.338μV,P<0.05)。
P1成 分 的 潜 伏 期在 各 电 极 点主 效 应 显 著 [F(4.420,159.105)=3.176,P<0.05,ηp2=0.081],电极点和组别之间无交互作用[F(8.839,147.803)=1.817,P=0.070,ηp2=0.092],组别主效应不显著[F(2,36)=0.144,P=0.886,ηp2=0.008]。
分别以P5、PO3、PO7和P6,PO4,PO8的平均值代表左右半球两侧枕区,对波幅和潜伏期进行3(组别)×2(脑区:左、右)两因素重复测量方差分析。结果发现,P1波幅仅在组别上主效应显著[F(2,36)=3.994,P=0.027,ηp2=0.182],P1潜伏期在左右半球和组别上均无主效应和交互效应。
以上结果显示,顶枕区诱发出明显的P1成分,其中,在枕区的波幅最大,且二级组波幅最大,专家组波幅最小,各组P1成分潜伏期无显著差异,左右半球无显著差异。
2.2.1.2 N1成分
总平均图显示,在60~140ms诱发出N1成分,选取额中央区、中央区、顶区、枕区中线的FCz、Cz、Pz、Oz 4个代表性电极,分别以波幅和潜伏期为因变量,进行3(组别)×4(脑区)两因素重复测量方差分析。结果显示:波幅在 各 电 极 点 主 效 应 显 著 [F(1.822,65.594)=11.987,P<0.01,ηp2=0.250],FCz点N1波幅最大。各脑区电极点和组别的 交 互 作 用 显 著 [F(3.644,65.594)=3.448,P<0.05,ηp2=0.161],组别的主效应不显著。进一步简单效应分析显示,在额中央区和顶区,组别主效应边缘显著[F(2,36)=3.10,P=0.057;F(2,36)=3.15,P=0.055],专家运动员在额中央区诱发的N1波幅最小,二级组在顶区诱发的N1波幅最小(图4b显示专家组蓝色区域最小)。峰潜伏期在各电极点主效 应 显 著 [F(2.515,65.594)=217.014,P<0.01,ηp2=0.858],各脑区电极点和组别的交互作用不显著,组别的主效应不显著。N1被证明与选择性注意有关。以上结果显示,优秀网球运动员在实验刺激材料呈现初期的注意和大脑激活水平低于新手。
2.2.1.3 N200成分
三组被试在-300~3 000ms的ERP总平均图显示(图2a),视频开始播放后150~300ms在前额区诱发出明显的负偏转波N200(图4),本研究从前至后选取各脑区中线5个电极点(Fpz、Pz、Oz、Cz、Fz)对 N2波幅和潜伏期进行重复测量方差分析。结果显示,波幅在各脑区主效应显著[F(1.979,71.242)=6.341,P<0.01,ηp2=0.150],各 脑 区和 组 别 无 交 互 作 用 [F(8,71.242)=1.616,P=0.327,ηp2=0.061],组别主效应不显著[F(2,36)=1.449,P=0.248,ηp2=0.074],进一步多重比较发现,各脑区诱发的N200波幅在前额达到最大,其次为顶区,即Fpz(-4.469±0.759μV)>Pz(-3.167±0.719μV)>Oz(-1.395±1.032μV)>Cz(-1.212±0.233μV)>Fz(-0.850±0.213μV)。潜伏期在组内和组间均无主效应和交互作用。
进一步选取前额Fp1、Fpz、Fp2 3个电极点,分别以波幅和潜伏期为因变量做重复测量方差,结果显示,组别主效应边缘显著[F(2,36)=3.213,P=0.052,ηp2=0.151]。多重比较证实,专家组在前额诱发的N200波幅显著低于新手组(P<0.05)(图4b蓝色区域显示,新手在前额区激活 最 明 显 )。 潜 伏 期 的 组 别 主 效 应 边 缘 显 著 [F(2,36)=3.221,P=0.052,ηp2=0.152],专家组 N200潜伏期最长。额区N200波幅在左右半球无显著差异。
2.2.1.4 P300成分
根据总平均波形图,选取中央区、顶区、枕区中线的Cz、Pz、Oz 3个代表性电极,分别以波幅和潜伏期为因变量,进行3(组别)×3(脑区)两因素重复测量方差分析。结 果 显 示 ,波 幅 在 各 脑 区 主 效 应 显 著 [F(1.460,52.558)=57.857,26 P<0.001,ηp2=0.616],各脑区波幅大小为:枕区(8.493±0.952μV)>顶区(3.977±0.649μV)>中央区(1.411±0.270μV),且两两比较差异显著(P<0.01)(图4b显示,枕区电流密度最大)。脑区×组别的交互作用不显 著 [F(2.920,52.558)=0.316,P=0.808,ηp2=0.017],组 别 主 效 应 不 显 著 [F(2,36)=1.019,P=0.371,ηp2=0.054]。
潜 伏 期 在 各 脑 区 主 效 应 显 著 [F(2,72)=3.374,P<0.05,ηp2=0.086],组别主效应不显著[F(2,36)=1.711,P=0.195,ηp2=0.087]。脑 区 × 组 别 的 交 互 作 用 显 著[F(4,72)=3.722,P<0.01,ηp2=0.171]。进一步简单效应检验发现,潜伏期在枕区(Oz)的组别主效应显著[F(2,36)=7.24,P<0.01],各组P300潜伏期大小为:新手(339.846±38.914ms)> 二 级 组 (311.455±42.681ms)> 专 家(285.867±31.713ms)。P300的潜伏期代表了大脑对外部刺激进行分类、编码、识别的速度。
以上结果表明,在刺激任务出现的感知阶段,专家运动员的识别速度快,且经过统计检验发现无明显的性别差异。
2.2.2 晚期慢波
500~3 000ms的晚期慢波反映了被试接受刺激信息后在大脑中进行编码和信息加工的过程。根据相关文献及对总平均图波形走势的观察,本研究将Epoch 2晚期成分划分为4个阶段(Segment1~4),对各组被试在各阶段ERP成分的平均波幅进行统计分析(图5):1)500~1 100 ms,本段波形显示出明显的晚期负走向成分LNC(late negative component);2)1 100~1 700ms,显示出波幅为负的正走向波形,我们认为是LNC的延续;3)1 700~2 500 ms,显示出明显的负走向的波形LNC;4)2 500~3 000 ms,显示出正走向的波形LPC(late positive component,或称为 LPP,late positive potential)。
为了更细致地比较各组ERP成分在前、后脑区及大脑左右半球分布随时间变化的趋势及差异,将各段进一步划分为14个时间窗口(Time Window,表2)进行统计分析。将大脑皮层从前至后划分为前额、额区、中央区、顶区、枕区5个区域,并分别取每个区域内左右半球共10个代表电极(左前额区Fp1、右前额区Fp2,左侧额区F3、右侧额区F4,左中央区C3、右中央区C4,左侧顶区P3、右侧顶区P4,左侧枕区O1、右侧枕区O2),对各时间窗口的ERP平均波幅进行3(组别)×5(前后脑区)×2(左右半球)的三因素重复测量方差分析,统计结果的F值和差异显著性(表2),对主效应和交互作用有显著差异的结果进一步进行多重比较和简单效应分析。
左、右半球差异:各时间窗口统计结果显示,各组在800~1 900ms的6个时间窗口内平均波幅在大脑左右半球的主效应显著(P<0.05),电流密度图可见明显的左半球优势,且专家组左右半球均有激活(图5c~g)。在500~600ms、2 100~2 500ms时间窗内表现出左右半球×组别的显著交互作用(P<0.05)。
前后脑区差异:不同组别在各时窗内各脑区的激活有显著差异(图5可见不同脑区电流密度差异明显,P<0.05)。脑区×组别在1 900~2 500ms交互作用呈边缘显著,在2 500~3 000ms存在脑区×组别的显著交互作用(P<0.05),在1 400~1 900ms存在显著的脑区×半球交互作用(P<0.05)。
熵权法[9]作为一种客观的赋权方法,能够在权重信息未知且以评价值区间数形式给出的情况下求出指标权重.通过信息熵公式可以看出:熵值越大表明指标值的无序性越低,提供的有用信息越少,权重越小;反之指标值的无序性越高,权重越大.在衡量智慧城市建设过程中的公众参与水平时,可以根据指标的离异程度,在计算信息熵的基础上得到各评价指标的熵权,最终得到相对客观的指标权重.
组别差异:在500~600ms,1 700~3 000ms时间窗内,组别主效应显著(P<0.05)。事后比较发现,在500~700ms,900~1 000ms,1 400~3 000ms时窗内,专家组和新手组在各脑区诱发LNC和LPP的平均波幅均有显著差异(P<0.05)。
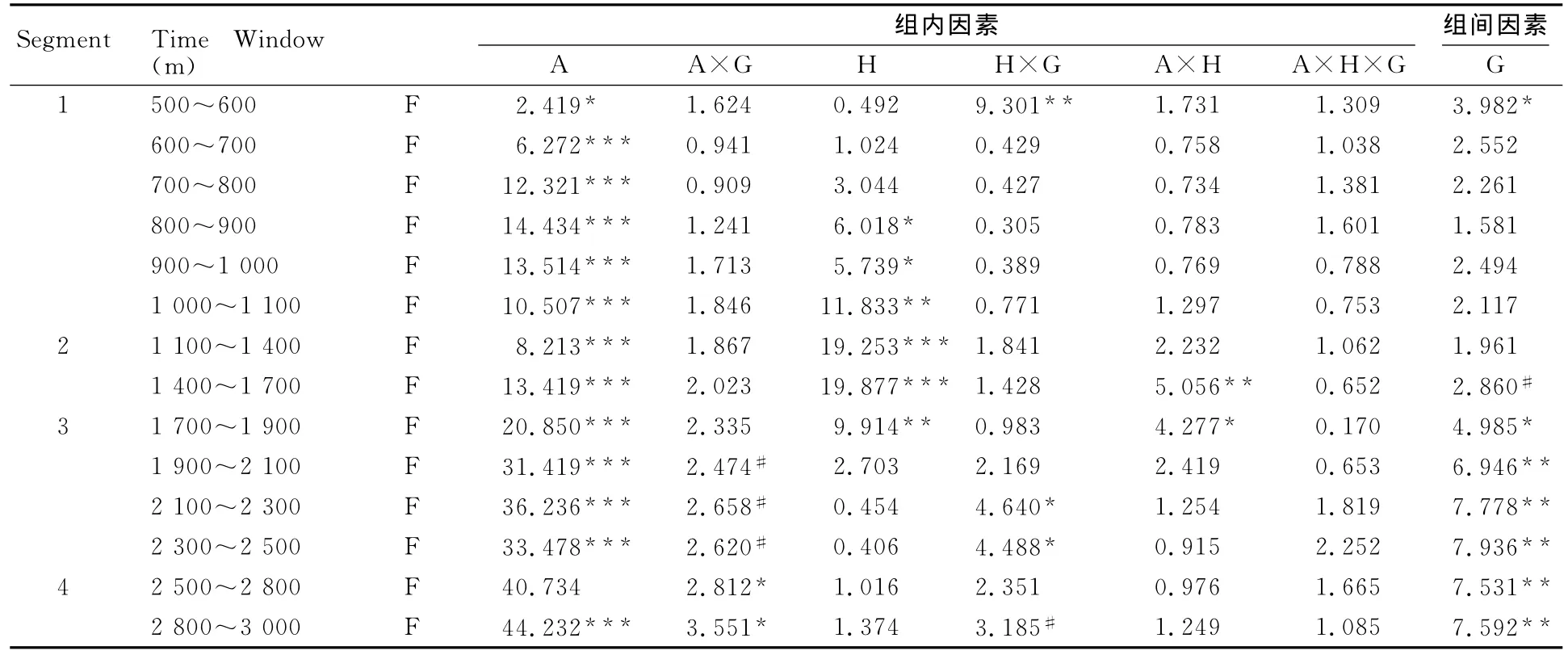
表2 本研究500~3 000ms晚期成分各时程平均波幅显著性效果分析一览表Table 2 Significant Effects of Mean Amplitude from the Analyses of Late ERP during 500~3 000ms
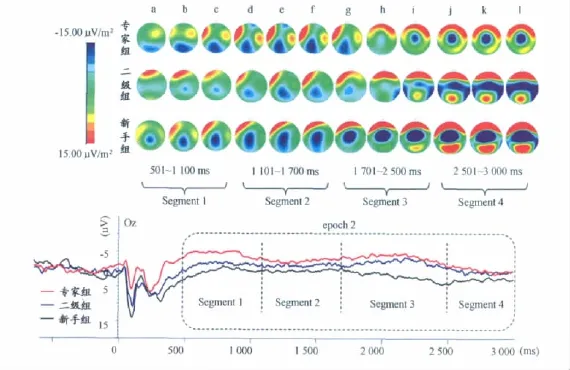
图5 Epoch2各段波形图及脑电流密度图Figure 5. Oscillogram and CSD Maps of Epoch2
对以上有交互作用的变量进一步简单效应分析发现:在额区,新手组左、右半球的主效应显著[F(1,36)=8.04,P<0.01];在 左 侧 额 区 (F3)组 别 的 主 效 应 显 著 [F(2,36)=3.75,P<0.05],新手的平均波幅最大,专家最小。在中央区,新手左右半球的主效应显著[F(1,36)=4.39,P<0.05],左半球(C3)组别的主效应显著[F(2,36)=3.91,P<0.05]。顶区专家组左右半球(P3、P4)主效应显著[F(1,36)=4.26,P<0.05],左半球组别主效应显著[F(2,36)=7.14,P<0.01]。
同时,通过对专家组男、女运动员的数据进行了统计分析发现,在500~3 000ms的信息加工过程中的认知特征并未表现出明显的性别差异。
2.2.3 决策阶段ERP成分
以被试对击球线路做出决策的反应时为依据,观察所有被试叠加后的ERP波形发现,在视频结束后的200~400ms时间段内诱发出明显的负偏转波,计算得出峰值平均潜伏期为273ms,因此,我们将其命名为N250。选择额区Fz、中央区Cz、顶区Pz和枕区Oz 4个电极点对3组被试的ERPs进行叠加处理后分别以N250的峰值波幅和潜伏期为因变量进行3(组别)×3(视频定格时间)×4(不同脑区电极点)的三因素重复测量方差分析。结果显示,N250峰值在不同电极点主效应显著[F(2.094,67.017)=3.482,P<0.001,ηp2=0.398],电极点×组别交互作用显著[F(4.189,67.017)=5.406,P<0.01,ηp2=0.253];视 频 定 格 时间 × 电 极 点 × 组 别 的 交 互 作 用 显 著 [F(5.473,88.751)=2.350,P<0.05,ηp2=0.128],组 别 主 效 应 非 常 显 著 [F(2,32)=11.239,P<0.001,ηp2=0.413]。进一步简单效应检验发现,被试在对击球前80ms的视频做出预判时,在顶枕区(Pz、Oz)诱发出明显的负偏转波,且组别主效应显著[Pz:F(2,32)=3.48,P<0.05;Oz:F(2,32)=3.78,P<0.05],专 家组诱发的N250波幅最大。
潜伏期的视频定格时间×电极点交互作用显著[F(4.614,147.643)=3.392,P<0.01,ηp2=0.096],视 频 定 格 时间 × 电 极 点 × 组 别 交 互 作 用 边 缘 显 著 [F(9.228,147.643)=1.920,P=0.052,ηp2=0.107],组别主效应显著[F(2,32)=4.419,P<0.05,ηp2=0.216]。进一步简单效应显示,被试在对手将球打出手80ms以后做出判断时,在枕区的组别主效应显著[F(2,32)=4.99,P<0.05],新手在枕区诱发的N250潜伏期最长。
3 讨论分析
3.1 网球运动员判断击球线路的行为优势
本研究中,高水平网球运动员在比赛相持阶段判断击球线路时仅在对手触球前80ms表现出明显的反应速度优势,且在各时间点的判断准确率显著高于新手,表明专家运动员能在信息量的呈现尚不完整时快速有效利用击球前的信息。
实验中的专家组运动员均为国家一级和健将级运动员,训练年限超过10年,多次参加国内外各级水平比赛,临场(馆)经验丰富,基本代表了同年龄段我国网球的最高水平。长期的专项技能训练和理论知识学习以及丰富的比赛经验使运动员对专项技能的知觉分化水平获得提高,也使储存在长时记忆中的内容更加丰富,提高了知觉的敏感性,从而在知觉过程中能够快速对大脑中的经验信息做出归类分析和判断,属于自上而下的加工方式,专家运动员决策过程中的策略是在保证准确率的前提下以快速取胜。而新手组运动员为网球专项大一新生,由于学习时间短,大脑中储存的相关信息少,还不具备监控对手及制定战术的能力。在比赛情境下,新手长时记忆中缺乏高效编码和处理环境线索的必要的知识结构[28],因此,无需经过模板匹配与比较的过程,刺激材料中呈现的不完整信息并未给他们提供有效的线索,其信息加工完全依赖于现实场(馆)景的表面特征,属于自下而上的加工方式。在实验要求“既快又准”的前提下,新手运动员更倾向于采用加快反应速度的直觉性决策策略,表现出“初生牛犊不怕虎”的状态。因此,无论从内部的信息加工方式还是外在的选择性注意,新手都区别于经验丰富的运动员。这一结果与已有关 于 网 球 运 动 员 预 期 能 力 的 研 究 相 一 致[18,23,29,37]。 同 时 ,在 乒 乓 球[4,10,12]、羽 毛 球[7,19]等 技 能 主 导 类 隔 网 对 抗 项 目中,专家运动员的行为优势也同样得到证实。
实验结果还发现,二级运动员判断准确率高于新手,但预判的速度最慢。二级运动员作为从新手到专家发展过程的过度阶段,其运动技能已达到较高水平,但由于大赛经验不足,在决策绩效上仍与专家存在明显差异。在观看刺激材料时,二级运动员虽然能有意识的将运动场(馆)景与大脑中的经验模板相比较进行模式识别,但在任务加工时,为了保证准确率,检索和提取相关信息的耗时较长,延长了决策时间,也即我们所说的“速度准确性权衡”。
3.2 网球运动员早期注意阶段的ERP特征
本实验目的是检验不同水平网球运动员对网球比赛相持阶段对手最后一拍击球线路预判的绩效及神经生理特征,因此,实验刺激材料呈现的前两拍球作为前摄信息提供给被试主要是考察被试接受刺激并根据特定运动情境快速有效提取信息的能力。
对ERP的分析结果显示,在实验材料呈现早期,顶枕区诱发出明显的P1成分,且二级组在枕区波幅最大,专家组波幅最小,潜伏期在组间无显著差异,左、右半球激活无显著差异。视觉P1代表了知觉加工的早期阶段,是视觉诱发电位(visual evoked potentials,VEP)的一个分支。最初对视觉P1的描述来自于Spehlmann(1965),他将P1定义为“出现在80~120ms之间的表面正成分(surface positive component at 80-120ms)”。自20世纪末,P1开始在选择性注意领域中开展研究。Van Voorhis等(1977)通过实验证明,注意可以调节早期的视觉诱发电位P1[40]。另有学者研究指出,视觉P1成分代表了对刺激空间方位的辨别,且主要与心理资源的利用有关,其效果反映了注意的消耗(cost of attention)[24,26]。 当 大 脑 在 对 刺 激 进 行 加 工 和 提 取信息时需要占用认知资源,刺激越复杂或加工任务难度越大,占用的认知资源越多。通常在认知系统内专门负责注意执行的脑功能网络会受到我们的控制,以便将认知资源分配到重要刺激上。
专家网球运动员诱发的P1波幅较小,表明投入认知资源少,可能是源于长期训练及丰富的比赛经验的积累,使其能将有限的脑资源进行合理分配,在不断变化的比赛情境中快速扑捉到对手的关键信息,对于次要信息则无需投入更多的脑资源,表现出能量节省化。而二级组运动员在从任务目标出现、视觉搜索的早期阶段就投入了较多心理资源,努力搜寻大脑中储存的记忆片段,因此,顶枕区诱发出P1成分的峰值显著高于专家组运动员。
另一个成分N1作为注意条件下辨别加工的有效指标,主要反映了注意的认知识别。在视觉工作记忆编码和保持的不同阶段对材料的加工程度不同,加工时间越长越精细,其波幅大小与注意程度密切相关[1]。N1的波幅在注意条件下明显大于非注意条件,反映了注意焦点内(刺激自身特征信息)的识别过程,或是对特定区域的辨别过程[41]。本研究中,专家组被试N1的激活程度最小,进一步证实了网球专家运动员在实验材料呈现时对早期刺激信息的注意和激活水平并不高,表现出认知资源的节省化。
N200(N2)成分主要反映的是识别靶刺激特征的心理加工过程[17]。早期很多研究将N2和P3联系起来,称为“N2-P3混合体(N2-P3complex)”,已有研究将大脑额-中央部的N2称为前部N2s(anterior N2)。近年来,出现了很多关于前部N2s在行为前对信号的检测、策略制定,行为中对策略的监控,对动作的及时控制,行为后对结果反馈信息的加工,对策略的调整等一系列认知控制加工中的作用[21]。而大脑后部N2反映视觉皮质加工刺激时的注意程度,且N2波幅随注意范围的扩大而增大。本研究中,N200波幅在前额达到最大,且专家组显著小于新手组,表明专家运动员在行为前能很快将注意力集中于关键信息,在短时间内完成对手信息的识别与检测并在动态变化的攻防过程中及时调整策略,在识别靶刺激特征方面表现出优势。新手由于经验不足,不能对关键信息进行有效检测,而是通过场(馆)上运动员及周围环境等多方面信息寻求策略,在行为监控和决策过程中耗费了较多的注意资源。
P300是神经中枢在感受信息刺激过程中产生的晚成分,反映感知、注意、判断、鉴别、传递、期待等复杂的多层次心理认知活动。在此过程中,注意起着决定性的作用。以往的研究发现,P300与信息量提取数量有关,其波幅与所投入的心理资源和任务难度成正相关[22],亦或是反映了被试对反应的自信程度[14]。本研究结果显示,P300波幅在枕区最大,其次是顶区和中央区。头皮后部的P300被证明与目标事件的自上而下的、内源性的和主动的加工有关[1]。P300的潜伏期与视知觉、视觉记忆、注意、视觉运动速度和运动控制能力相关,代表了大脑对外部刺激进行分类、编码、识别的速度,或者说反映了刺激评价所需要的时间[6]。本实验结果表明,被试在刺激材料出现后选择性注意与靶刺激相关的信息,把主要注意力放在感知材料和模式辨认上,把感觉、知觉、记忆过程结合起来,从长时记忆结构中搜寻、提取恰当的信息。在此过程中,专家组运动员诱发的P300成分潜伏期明显短于新手组。Jin H等(2011)通过实验研究职业和非职业羽毛球运动员对球落点的预期,也同样得出职业运动员的P300潜伏期比非职业组短[19],证明了高水平运动员对刺激材料的识别和编码速度快。
在以往的相关研究中,冯琰、周成林等(2008,2011)研究发现,高水平花剑运动员在静态运动情景中分辨识别技术动作以及在空间知觉实验中特定脑区诱发的P1、N1和N2峰波幅均低于普通大学生组,且P3出现的时间较早[5,14]。本研究与其结果一致。而赵洪朋(2010)[13]、王小春(2012)[8]在研究散打运动员特征搜索和网球运动员时间序列知觉的实验中均发现,专家组N1的波幅显著大于新手组。Jin H(2011)研究发现职业羽毛球运动员预判球落点时诱发的P300波幅大于非运动员[19]。对同一ERP成分在不同实验中结果不一致的原因可能是实验材料类型及要求被试完成任务的差异导致被试采取了不同的知觉策略。
本实验综合P1、N1、N2、P300成分的分析结果,进一步表明专家网球运动员在刺激材料呈现早期投入的认知资源少,对目标进行识别和编码的速度快。
3.3 网球运动员晚期类比加工过程的ERP特征
本研究中,通过划分时间窗口分析了视频播放全程的ERPs,对500~3 000ms时程的分析发现,500~1 100 ms、1 400~2 500ms两个时间段内出现明显的LNC(late negative component)成分。Nessler等(2006)曾在研究语义记忆推理过程中出现相似的ERP成分,认为在做出反应之前出现的负的ERP成分反映了执行过程。比如说,在记忆中寻找潜在的执行方案,注视相关信息,寻找策略等等[30]。事实上,选择性的检索依赖于抑制相互矛盾的记忆痕迹,其中,额区诱发晚期成分的波幅增大与检索过程中遗忘任务的过程相关联[20]。Morrison等(2004)提出,LNC成分应该与概念间的类比映射加工相关,反映了隐喻词对类推需要在工作记忆中对概念间关系进行比较、平行联接以及结构对齐等与类比映射相关的加工进程[29]。Alan(2008)在研究面孔识别的过程中发现,晚期的负成分反映了在意识后对刺激的识别过程[32]。除此以外,有学者认为,LNC成分在错误信念推理、条件推理等过程中均有诱发,并且当需要在工作记忆中进行再认、信息维持等操作任务时,也均会诱发类似的头皮慢负电位[27,35,42]。可见,LNC成分与信息加工过程中的识别、类比、推理和执行策略等密切相关。
本实验中,刺激材料呈现的时长是3 000ms,在500 ms左右的时间点,实验材料中对手的第一拍球已经出手,如果说,500ms以前是前注意阶段,那么,此时物体特征还处于“漂移”状态,加工方式是平行加工或自动加工,专家并未投入较多注意资源。而在500~700ms时间段,专家组诱发的LNC比二级组和新手组出现了更明显的负偏转,说明专家在视频开始500ms以后,才开始将注意资源分配于场(馆)上双方运动员在进攻和防守过程中的击球方式、球速、击球特点等关键信息,并对情境中的对抗和进攻形成表征,同时对其特征进行类比加工,抽取当前刺激各方面与记忆中的模式特征相匹配的成分进行特征整合。本阶段作为模式识别的第2阶段,加工方式是控制加工或系列加工。只有到了后期的特征整合阶段,对物体特征的知觉才能形成。专家在确定方案之前,往往会花费更多的时间和精力来建立对问题情境的表征,并对此进行评估和分类,因此,诱发的LNC波幅大,即专家组出现了更明显的负偏转成分。
有学者认为,LPP(或LPC)属于P300家族,与刺激的唤醒度、注意及动机的参与有关[3,9]。影响LPP的因素中,注意资源的投入和刺激的唤醒度高低是两个非常重要的部分,注意资源的投入也与被试的参与动机有关。另外,LPC更多的与外显记忆关联,其波幅大小受刺激的重复性和相似性影响,加工操作水平的程度影响LPC的波幅,加工程度越深波幅越大。因此,LPC的波幅与回忆和检索的成功率密切相关。Pastor(2008)研究不同情绪图片诱发的LPP揭示,LPP反映了最初对结果的评估、分类,以及注意的收集[31]。LPP波幅差异主要集中在额叶和额中央部位,这种持续的正电位反映了图片识别任务中个体投入的注意资源量和动机参与程度。本实验的波形图和电流密度图(图5)显示,晚期的持续正成分主要出现在前额和额区,且随着时间的推移激活水平逐渐提高。前额叶对工作记忆的信息暂存、编码、激活、控制和协调至关重要。这一结果符合实验过程中被试信息加工的过程和特点,也与以往多数研究相一致。
关于左、右半球激活的差异,本实验结果发现,各组在800~1 900ms的6个时间窗口内平均波幅在大脑左右半球的主效应显著,各组均表现为左半球波幅大于右半球,且新手组的平均波幅大于二级组和专家组,专家组的平均波幅最小。有学者通过研究提出大脑两半球功能单侧化理论,认为在大脑左、右半球之间存在信息加工的差异[15],左半球以零碎的、分析的和序列的方式加工信息,而右半球是以直觉的、完形的、综合的方式加工信息[11]。陈熙熙等(2004)的研究也进一步说明,大脑功能的单侧化依赖于自上而下的主动认知过程,是任务驱动而非材料驱动[2]。但两个半球在加工时是分工与合作不断变化的动态关系[25],个体倾向使用右半球或左半球的加工方式或策略,并不意味着另侧半球不发挥作用,半球之间脑电活动的相对差异比某个半球的活动水平更能说明个体的认知风格。本实验中,之所以在视频呈现初期的500ms内没有出现明显的左、右半球激活的差异,可能是因为此时任务的难度还未出现明显差异,半球不对称反映的是相对的效率不同而不是绝对功能的差异。电流密度图也显示(图5b),专家组运动员的信息加工方式不同于二级组和新手组,其大脑右半球也有不同程度的激活,说明专家组更善于将左半球分析的和序列的加工方式与右半球直觉的、综合的加工方式相整合,这可能是专家组表现出认知优势的重要原因之一。
3.4 网球运动员决策阶段的ERP特征
本实验中,各组被试在实验材料播放结束后诱发的N250成分峰值潜伏期在反应时范围内,且潜伏期短于RT,专家组在顶枕区的N250波幅显著高于二级组和新手组。
N250被认为是识别任务中识别靶刺激时诱发的典型ERP成分,常在大脑后部电极点记录到。Tanaka等(2006)在研究面孔识别时认为,N250成分可以反映特殊的、不熟悉的面孔在短期记忆痕迹中的激活,同时,也可以激活过去储存在长时记忆中的熟悉面孔[39],重复呈现一些熟悉政治家和名人的面孔比重复呈现新异的面孔诱发出了更大波幅的 N250[36]。Pierce(2011)研究人对熟悉面孔和新异刺激识别的ERP时发现,N250的波幅受到熟悉程度的影响,当看到熟悉的面孔时,会比看到新异刺激诱发出更负的 N250[34]。
以上研究结果均表明,N250成分代表了个人对已有知觉记忆的提取,当储存在长时记忆中熟悉的内容呈现于面前时,相比出现新异刺激能诱发出更大波幅的N250成分。因此,本研究中,被试在顶枕区诱发出明显的N250成分,且专家组诱发的N250波幅最大,推断其原因是专家组运动员经过多年的训练及比赛积累了丰富的经验,各种类型的攻防对抗情境已熟记于心,实验中呈现的刺激材料与其储存在长时记忆中的模板相匹配程度高。在观察实验材料的3 000ms过程中,当熟悉的比赛画面反复呈现于大脑中时,专家组能够快速提取、整合信息,及时调整策略,快速做出决策,因此,专家组运动员诱发出更负的N250成分。同时,在行为上表现出反应时间短,决策准确率高的优势特征。而实验中的新手由于学习时间短,大脑中储存的信息量少,大部分实验刺激材料是作为新异刺激呈现的,这种不熟悉的画面在短期记忆痕迹中的激活诱发出相对波幅较小的N250成分。同时,由于很难在短时间内搜寻到与任务相匹配的模板,导致新手决策反应速度慢,且准确率显著低于专家组运动员,尤其在对手击球前80ms的时间点,专家组运动员的N250波幅显著大于新手,进一步印证了在该时间点专家组反应 速度显著快于新手组的事实,揭示出专家组运动员在信息呈现不完整时更善于把握有效信息和有利时机,即在任务难度较大时表现出更明显的决策优势。
4 结论
本研究通过考查不同水平网球运动员对击球线路的预判绩效,探索优秀运动员在技战术发挥过程中信息加工的事件相关电位特征,研究结果支持了“专家组具有认知优势”这一假设,得出如下结论:
1.专家组运动员在比赛相持阶段预判击球线路时能较早把握时机,行为特征表现为既快又准,反映了最优化理论。
2.专家组运动员能够合理分配认知资源。在早期阶段投入的认知资源少,而在场(馆)上双方运动员在进攻和防守过程中投入认知资源多;同时,专家组运动员更善于将左半球分析的和序列的加工方式与右半球直觉的、综合的加工方式相结合,符合整合加工理论。
3.专家组运动员的决策绩效依赖于在其长时记忆中储存了高效编码和处理环境线索的必要的知识结构,这与长期训练与比赛的经验积累密切相关,得到了经验说的支持。
[1]白学军,尹莎莎,杨海波,等.视觉工作记忆内容对自上而下注意控制的影响:一项 ERP研究[J].心理学报,2011,43(10):1103-1113.
[2]陈熙熙,苏彦捷.大脑功能侧化的心理学研究及分子和细胞神经生物学依据[J].中国神经科学杂志,2004,20(2):174-179.
[3]丁妮,丁锦红,郭德俊.个体神经质水平对情绪加工的影响-事件相关电位研究[J].心理学报,2007,39(4):629-637.
[4]段宇防.兵乓球运动员预期判断攻球线路的眼动特征与反应时研究[D].北京:首都体育学院,2008:25-28.
[5]冯琰.高水平花剑运动员认知优势特征及神经机制研究[D].上海:上海体育学院,2008:22-28.
[6]欧小凡,潘速跃,黄英.事件相关电位P300与认知相关研究[J].中国老年学杂志,2010,30(3):833-835.
[7]王洪彪.羽毛球练习者知觉动作技能认知加工特征研究[D].上海:上海体育学院,2009:37-38.
[8]王小春,周成林.基于时间序列情境的网球专家预判优势—来自ERP的证据[J].中国运动医学杂志,2012,31(4):345-352.
[9]吴燕,周晓林.公平加工的情境依赖性:来自ERP的证据[J].心理学报,2012,44(6):797-806.
[10]徐璐,乒乓球运动员决策过程认知加工特征的研究[D].上海:上海体育学院,2010:22-24.
[11]张晓文,余嘉元.半球认知风格:一种研究个体差异的新视角[J].南京师范大学学报(社会科学版),2007,3(2):106-111.
[12]张玉慧.高水平乒乓球运动员发球旋转判断过程中神经心理机制的研究[D].上海:上海体育学院,2011:17-18.
[13]赵洪朋.优秀散打运动员知觉预测过程认知特征与神经机制研究[D].上海:上海体育学院,2010:25-26.
[14]周成林,冯琰,王小春.高水平男子击剑运动员空间知觉特性的事件相关电位研究[J].中国运动医学杂志,2011,30(2):121-127.
[15]BAYNE T.The unity of consciousness and the split-brain syndrome[J].J Philoso,2008,105(6):277-300.
[16]FARROW D,REID M.The contribution of situational proba-bility information to anticipatory skill[J].J Sci Med Sport,2012,(15):368-373.
[17]HILLYARD S A,ANLLO-VENTO L.Event-related brain potentials in the study of visual selective attention [J].Proceed National Academy Sci,1998,95:781-787.
[18]HUYS R,CANAL-BRULAND R,HAGEMANN N.Global Information Pickup Underpins Anticipation of Tennis Shot Direction[J].J Motor Behav,2009,41(2):158-170.
[19]JIN H,XU G,ZHANG J X,et al.Event-related potential effects of superior action anticipation in professional badminton players[J].Neurosci Letters,2011,492 (3):139-144.
[20]JOHANSSON M,ASLAN A,BAUML K H,et al.When remembering causes forgetting:electrophysic ecological correlates of retrieval-induced forgetting[J].Cerebral Cortex,2007,17:1335-1341.
[21]JONATHAN R.FOLST E,PETTEN C V.Influence of cognitive control and mismatch on the N 2component of the ERP:A review[J].Psychophysi,2008,45:152-170.
[22]KOK A.On the utility of P3amplitude as a measure of processing capacity[J].Psychophysi,2001,38(2):557-577.
[23]LOFFING F,WILKES T,HAGEMANN N.Skill level and graphical detail shape perceptual judgments in tennis[J].Perception,2011,40(12):1447-56.
[24]LUCK S J,HILLYARD S A,MOULOUA M,et al.Effect of spatial cueing on luminance detectability:Psychophysical and electrophysiological evidence for early selection[J].J Experi Psychol:Human Perception and Performance,1994,20(4):887-904.
[25]LUO Y J,WEI J H,et al.ERP effects of recognition of Chinese spoken and written words and neural mechanism of retrieval[J].Acta Psychol Sinica,2001,33(6):489-494.
[26]MANGUN G R,HILLYARD S A.Modulation of sensory-evoked brain potentials provide evidence for changes in perceptual processing during visual-spatial priming[J].J Experi Psychol,1991,17(4):1057-1074.
[27]MCEVOY L K,SMITH M E,GEVINS A.Dynamic cortical networks of verbal and spatial working memory:Effects of memory load and task practice[J].Cerebral Cortex,1998,(8):563-574.
[28]MCPHERSON S L.Expert-novice differences in performance skills and problem representations of youth and adults during tennis competition[J].Res Q Exe Sport,1999,(70):233-251.
[29]MORRISON R G,KRAWCZYK D C,HOLYOAK K J,et al.A neurocomputational model of analogical reasoning and its breakdown in frontotemporal lobar degeneration[J].J Cognit Neurosci,2004,(16):260-271.
[30]NESSLER D,JOHNSON J R,BERSICK M,et al.On why the elderly have normal semantic retrieval but deficient episodic encoding:a study of left inferior frontal ERP activity[J].Neuroimage,2006,(30):299-312.
[31]PASTOR M C,BRADLEY M M,LOW A,et al.Affective picture perception:Emotion,context,and the late positive potential[J].Brain Res,2008,(1189):145-151.
[32]PEGNA A J,LANDIS T,KHATEB A.Electrophysiological evidence for early non-conscious processing of fearful facial expression[J].Int J Psychophysio,2008,70:127-136.
[33]PHERSON M C,SUE L.Expert-novice differences in planning strategies during collegiate singles tennis competition[J].J Sport Exe Psycho,2000,22(1):39-62.
[34]PIERCE L J,SCOTT L S,BODDINGTON S,et al.The N250 brain potential to personally familiar and newly learned faces and objects[J].Frontiers Human Neurosci,2011,10(31):1-13.
[35]QIU J,LI H,HUANG X T,et al.The neural basis of conditional reasoning:An event-related potential study[J].Neuropsycho,2007,(45):1533-1539.
[36]SCHWEINBERGER S R,HUDD Y V,BURTON A M.N250r:a face-selective brain response to stimulus repetitions[J].Neuroreport,2004,(15):1501-1505.
[37]SHIM J,CARLTON L G,CHOW JW,et al.The Use of Anticipatory Visual Cues by Highly Skilled Tennis Players[J].J Mo-tor Beha,2005,37(2):164-175.
[38]SMEETON N J,HUYS R.Anticipation of tennis-shot direction from whole-body movement:The role of movement amplitude and dynamics[J].Human Movement Sci,2011,(10):957-965.
[39]TANAKA J W,CURRAN T,PORTERFIELD A L,et al.Activation of pre-existing and acquired face representations:the N250ERP as an index of face familiarity[J].J Cogni Neurosc,2006,(18):1488-1497.
[40]VANVOORHIS S,HILLYARD S A.Visual evoked potentials and selective attention to points in space[J].Percep Psycho,1977,22(1):54-62.
[41]VOGEL E K,LUCK S J.The visual N1component as an index of a discrimination process[J].Psychophysio,2000,37:190-203.
[42]WANG Y W,LIU Y,GAO Y X,et al.False belief reasoning in the brain:An ERP study[J].Sci China Series C:Life Sci,2008,(51):72-79.
[43]WILLIAMS A M,WARD P,JOHN M.Anticipation Skill in a Real-World Task:Measurement,Training,and Transfer in Tennis[J].J Experi Psychol,2002,8(4):259-270.
猜你喜欢
癫痫与神经电生理学杂志(2022年6期)2022-02-10 13:55:46
口腔护理用品工业(2021年4期)2021-11-02 08:23:02
世界热带农业信息(2019年7期)2019-08-30 01:02:58
海峡姐妹(2018年12期)2018-12-23 02:39:20
价值工程(2018年25期)2018-09-26 10:14:36
作文通讯·高中版(2017年6期)2017-07-10 03:21:34
饲料与畜牧(规模养猪)(2017年1期)2017-02-28 03:10:15
西南交通大学学报(2016年4期)2016-06-15 20:29:36
中国医疗保险(2016年11期)2016-05-17 07:47:31
华北电力大学学报(自然科学版)(2016年3期)2016-04-25 05:06:56
