人骨髓间充质干细胞与血管内皮细胞的基因表达差异分析
2012-05-17 05:13刘丹丹陈丽琼银国利沈香娣
医学研究杂志 2012年7期
刘丹丹 陈丽琼 仇 容 银国利 沈香娣
间充质干细胞(mesenchymal stem cells,MSCs)是一种多能成体干细胞,因具有取材方便、增殖能力强、免疫原性低和广泛的分化潜能而成为目前细胞治疗、基因治疗和组织工程方面的研究热点。不同组织来源的MSCs在血管内皮生长因子(VEGF)、表皮生长因子(EGF)、碱性成纤维生长因子(bFGF)、地塞米松、低血清或无血清培养基等多种诱导体系的作用下,可以成功地定向分化为血管内皮细胞,不同程度表达内皮细胞相关表型,甚至形成血管网络样结构,成为血管组织工程和缺血组织细胞移植的理想种子细胞[1~3]。但MSCs分化为内皮细胞的机制目前尚不清楚,其分化过程的调控靶标尚未确立。本实验采用基因芯片技术对人骨髓间充质干细胞(hMSCs)和脐静脉内皮细胞(hUVECs)进行了基因组规模的表达差异分析,结果报告如下。
材料与方法
1.材料:hMSCs和hUVECs分别从健康志愿成人骨髓、脐带静脉分离获得;淋巴细胞分离液percoll、低糖DMEM培养基购自GIBCO;优等胎牛血清购自HYCLONE;UNIQ—10柱式RNA抽提试剂盒购自上海生工;基因芯片BiostarH—40S为上海博星基因公司产品。
2.方法:(1)hMSCs的分离、培养:取健康成人骨髓5~10ml,离心沉淀血细胞,叠加到percoll分离液上,离心后吸取单个核细胞层,10%FBS的L-DMEM悬起,接种于24孔培养板中。3天后换液,hMSCs贴壁生长。以后每3天换液,细胞80%融合,传代扩大培养,收集P5~P6代细胞待用。(2)hUVECs的分离、培养:酶消化法收集hUVECs,接种于培养瓶中,加M200培养基,扩增培养传至P3代备用。(3)hMSCs和hUVECs RNA的提取、分析:收集hMSCs和hUVECs各约(1~2)×107细胞,UNIQ-10柱式RNA抽提试剂盒分离细胞总RNA,测定所提RNA的OD260、OD280,计算RNA含量及纯度。琼脂糖凝胶电泳确认总RNA中18S、28S、5S RNA的完整性并进行热稳定性验证。(4)hMSCs和hUVECs RNA的芯片检测及分析:以hMSCs、hUVECs为实验组和对照组,分别用Cy5、Cy3荧光素标记,依次进行预杂交、探针标记、杂交、洗片、扫描与分析操作。Scan Array 4000扫描仪扫描芯片,GenePix Pro 3.0软件进行图像处理,ImaGene 3.0软件分析Cy5、Cy3荧光信号强度和比值。实验重复3次。每张基因芯片BiostarH-40S含4096个点,其中包括控制系统(148个基因)和有效基因(3948个基因)。控制系统中设16个空白对照用以排除芯片扫描时的背景信号,判别不同样品间是否存在交叉污染;16个阴性对照检验杂交时是否存在外源性污染;20个内参校准荧光信号强度;96个管家基因均衡Cy5、Cy3值,保证杂交结果的可靠性。(5)原位杂交检测:将细胞固定在玻片上,用含5μg/L蛋白酶,37℃水浴消化10min。PBS洗涤后,梯度乙醇脱水,干燥。加入地高辛—RNA探针进行杂交,PBS洗涤后加入碱性磷酸酶标记的抗地高辛抗体,孵育4h加入显色液,室温避光显色,镜下控制,终止反应。黑色颗粒为阳性结果。
结 果
1.hMSCs和hUVECs的RNA鉴定:经纯化,hMSCs和hUVECs的RNA OD260/OD280比值均在2.0左右,纯度较高,符合基因芯片检测的要求。变性琼脂糖凝胶电泳结果见图1,常温下和70℃保温1h电泳图谱上都可观察到清晰的28S、18S条带,5S条带相对较弱,证明了所提RNA的完整性和热稳定性。
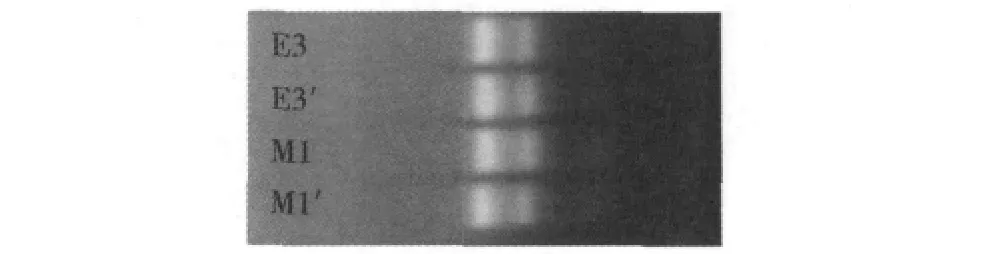
图1 hMSCs和hUVECs的RNA的琼脂糖凝胶电泳谱
2.hMSCs和hUVECs的基因芯片表达差异:根据差异表达筛选条件:①Cy5、Cy3值均>200或其一>800;②Cy5/Cy3比值 <0.5或者 >2。在所检测的3948条有效基因中,hMSCs和hUVECs的差异表达基因为627条,3321条基因在 hMSCs和hUVECs之间表达无明显差异,约占有效基因的84%,表1列出了非差异表达的前19条基因。差异表达的627条基因中,hMSCs高表达基因283条,约占7.2%,表2列出了显著高表达的前19条基因;hMSCs低表达基因共344条,约占8.7%,表3列出了显著低表达的前19条基因。
3.原位杂交分析:为了检验基因芯片检测结果的可靠性,我们采用了原位杂交方法来验证hMSCs和hUVECs细胞中部分差异表达的基因,包括hMSCs高表达的纤维连接蛋白(FN1)基因、Ⅳ型胶原基因、表皮生长因子受体(EGFR)基因和血小板源性生长因子受体(PDGFR)基因;hMSCs低表达的Ⅷ因子基因。杂交结果与基因芯片结果一致。
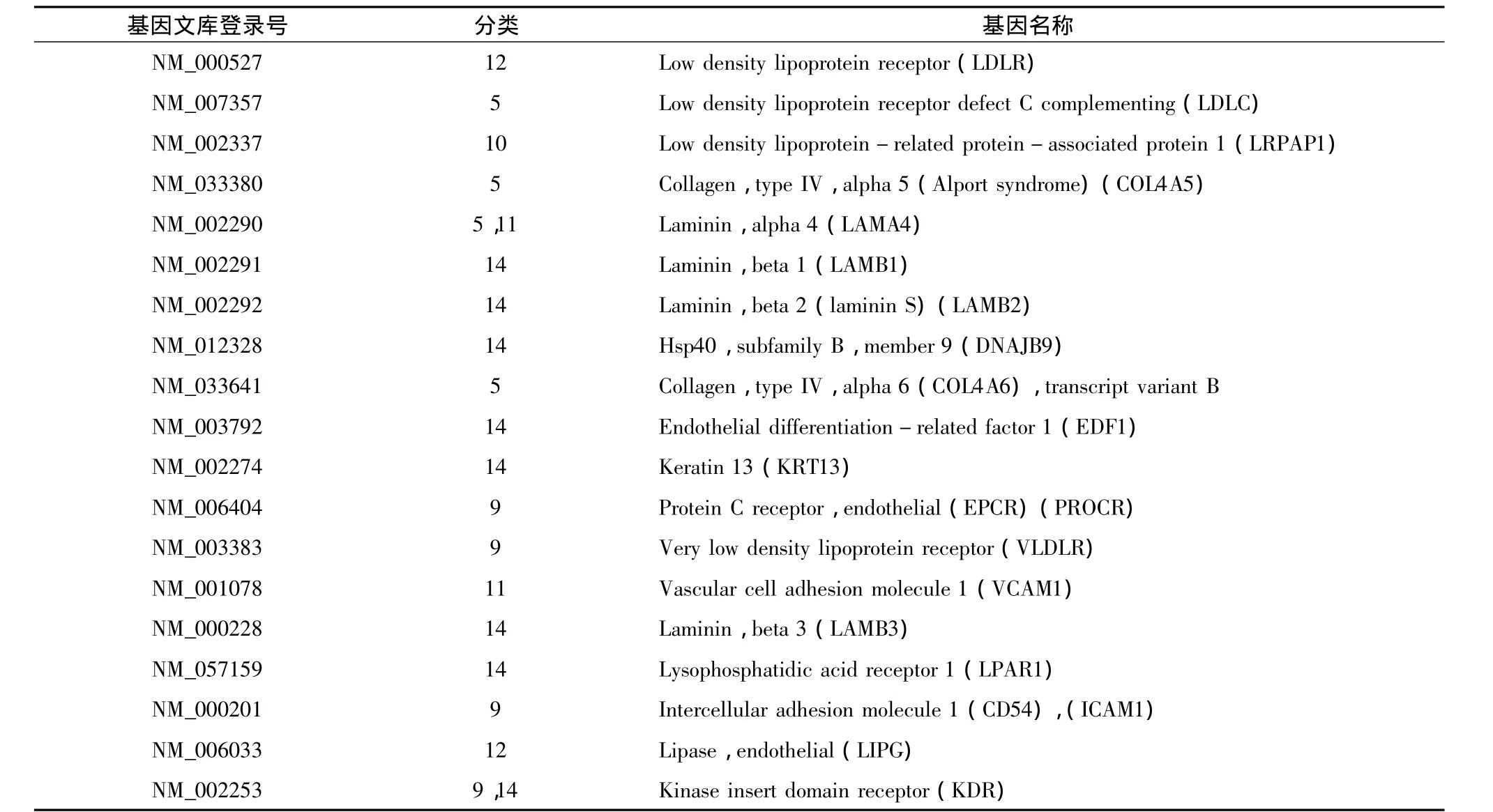
表1 hMSCs和hUVECs共表达的部分基因
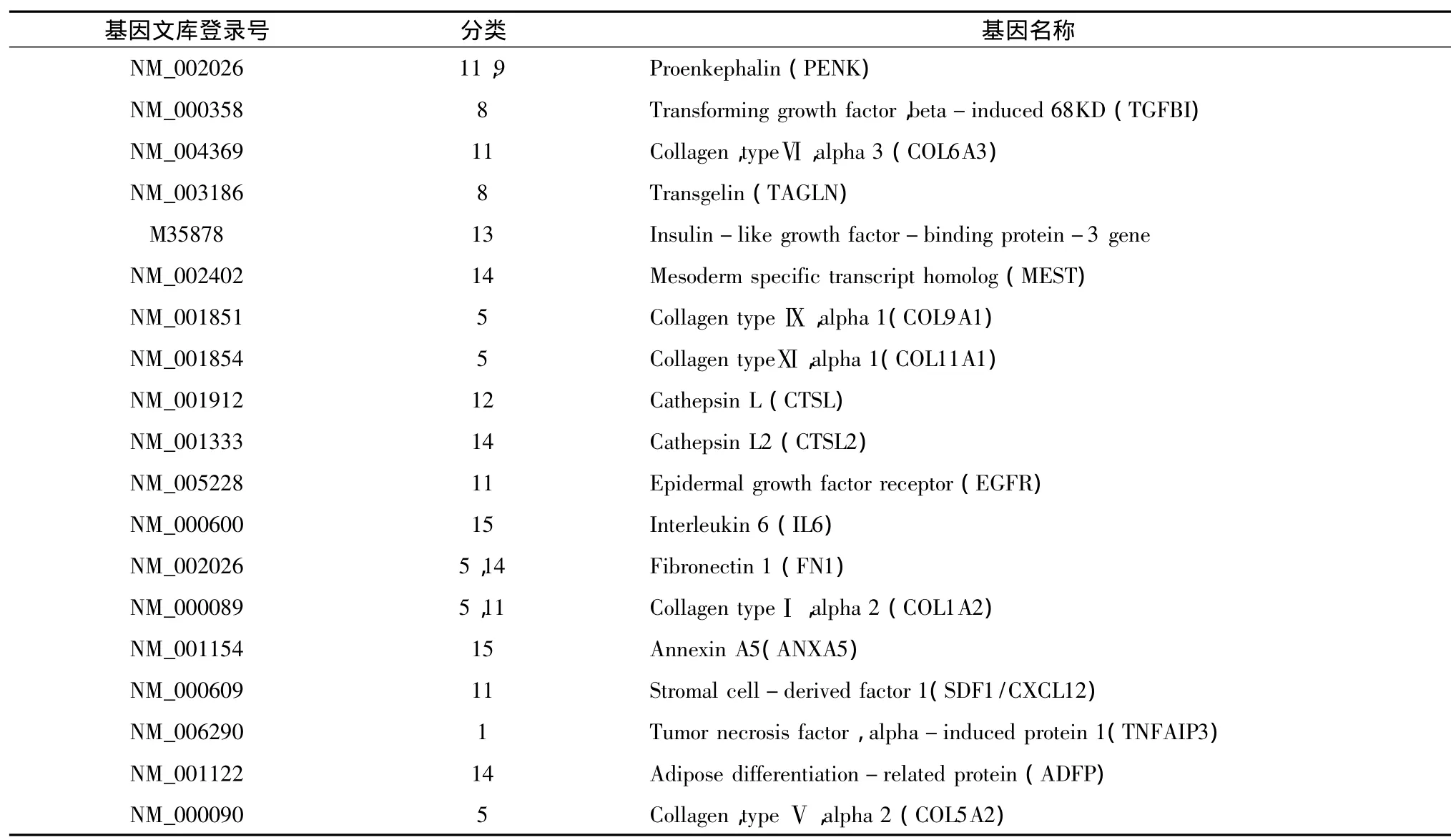
表2 hMSCs高表达的部分基因

表3 hMSCs低表达的部分基因
讨 论
从基因芯片检测的hMSCs和hUVECs表达的基因功能来看,主要包括以下5大类:①细胞骨架和运动蛋白基因;②细胞受体相关基因;③细胞信号和传递蛋白相关基因;④发育相关基因;⑤蛋白翻译、合成相关基因。
在非差异表达基因中,hMSCs除了显著高表达Ⅳ型胶原、层黏连蛋白、细胞间黏附分子1(ICAM-1)、血管细胞黏附分子1(VCAM-1)等和内皮细胞相关的细胞外基质和黏附分子家族基因外,还高表达内皮分化相关因子1(EDF1)、溶血磷脂酸受体1(LPAR 1)、DNAJB9、血管内皮生长因子受体2(KDR)等8个内皮细胞早期分化发育相关基因。
EDF1调控内皮细胞分化,在连接调控蛋白和基础转录机制上起桥接分子的作用,在通用转录因子TATA结合蛋白和基因特异性激活因子之间起转录辅助激活因子的功能,控制内皮分化相关基因的转录。溶血磷脂酸受体1(LPAR 1)是G-蛋白偶联的受体超家族成员,和溶血磷质酸结合后参与细胞信号传导和内皮分化鞘磷脂G-蛋白偶联受体介导的多种生物学功能,包括细胞增殖、迁移等。近来已有实验证明人脂肪组织来源的MSCs在人肺腺癌细胞A549的诱导下,通过LPA-LPAR1介导的旁分泌机制在肿瘤微环境的血管生成中有重要作用[4]。DNAJB9属于热休克蛋白Hsp40家族成员,调节70 kDa HSP的ATP酶活性,参与内皮细胞内质网应激反应中,组织相容性白细胞抗原HLA-B35上调内皮素1和下调NO合酶的过程[5]。因此,MSCs具有内皮分化的分子基础,为MSCs作为血管组织工程的种子细胞提供了细胞遗传学和免疫学依据。
以内皮细胞作对照,hMSCs主要高表达纤维连接蛋白1(FN1)和Ⅰ、Ⅴ、Ⅵ、Ⅸ、Ⅺ型胶原等细胞骨架蛋白基因;转化生长因子(TGF)、胰岛素样生长因子(IGF)、基质细胞源性因子1(SDF-1)、脂肪分化相关蛋白和表皮生长因子受体(EGFR)等与多种方向细胞分化发育相关的细胞因子/受体基因。FN与细胞膜受体整合素间的相互作用是细胞黏附与迁移的重要机制,其分子中的RGD(精氨酸-甘氨酸-天冬氨酸)位点是重要的结构基础,可与多种细胞外基质及细胞表面受体相结合,增强细胞间、细胞与基质间的紧密接触,影响细胞内骨架蛋白的组装,增强细胞的定向迁移,从而影响细胞的生长分化。SDF-1基因编码基质细胞来源的旁分泌家族成员,和受体CXCR4结合可活化血细胞,促进细胞的迁移和肿瘤的转移[6]。
相反,hMSCs低表达的344个基因中,细胞信号和传递蛋白基因所占比例最大,如1-磷酸-鞘氨醇受体1(S1PR1)、CTNNB1、PDGFB 等;其次是细胞发育相关基因,如趋化因子受体4(CXCR4)、血管生成素2(ANG-2)、血管内皮生长因子(VEGF)等;再次是蛋白翻译、合成基因,如vWF、CD34等。
趋化因子受体4(CXCR4)是CXC趋化因子SDF-1的特异性受体,两者在卵泡生长、发育和血管新生过程中有重要作用[7]。VEGF可刺激内皮细胞CXCR4的表达,增加对SDF-1的反应性,促进新生血管的形成,其机制可能和VEGF刺激转录因子与CXCR4基因启动子区域SP1位点的结合有关。β1连环蛋白(CTNNB1)是一重要的细胞黏附分子和信号传导因子,与E-Cad结合后形成细胞内的黏着复合体,起转录激活作用,启动细胞增殖相关基因转录,使细胞大量增殖。VEGF基因启动子包含7个CTNNB1的结合位点,CTNNB1可调节VEGF的表达,两者呈正相关关系[8]。PDGFB是PDGF家族成员之一,是间充质细胞的重要有丝分裂因子,在促进细胞增殖和血管生成方面有积极作用。因此,VEGF和PDGF成为近年来诱导MSCs内皮分化的首选诱导因子。
1-磷酸-鞘氨醇受体1(S1PR1)参与脂类分子信号转导通路,激活后可诱导细胞间的黏附,是细胞内的信号传导分子和促血管生成因子。S1PR1和配体SSP(鞘氨醇1-磷酸化酶)的结合具有高亲和性、高特异性,并可激活多种信号转导途径,诱导与血管生成有关的反应[9,10]。血管生成素 -2(Ang-2)是血管生成素1和内皮细胞酪氨酸激酶(TIE-2)拮抗剂,所编码的蛋白可阻止血管生成素1引起的血管重建,诱导内皮细胞凋亡。这些基因的功能虽然已有深浅不同的研究,但大多是在其他细胞或动物体内进行的。在骨髓间充质干细胞内,尤其是在间充质干细胞向内皮细胞分化过程中的作用和机制还有待进一步研究。
以上实验研究可见,由骨髓间充质干细胞诱导成血管内皮细胞是可行的,但分化机制是相当复杂的,涉及多基因的表达变化,包括一系列细胞骨架蛋白的重构;不同方向细胞分化相关基因表达的下调;细胞信号和传递蛋白相关基因的表达增强以及内皮相关发育基因的上调,还涉及一些DNA结合、转录、转录因子基因和蛋白翻译、合成相关基因的变化。这些差异表达的基因代表了一个与MSCs内皮分化相关的、特异的基因表达谱。正是这些基因的表达变化,使MSCs在各种内皮诱导体系的作用下表现出内皮细胞的抗原标志,甚至表现出内皮细胞的相关功能。
1 Oswald J,Boxberqer S,Jorqensen B,et al.Mesenchymal stem cells can be differentiated into endothelial cells in vitro[J].Stem cells,2004,22(3):377-384
2 Liang F,Wang YF,Nan X,et al.In vitro differentiation of human bone marrow-derived mesenchymal stem cells into blood vessel endothelial cells[J].Zhongguo Yi Xue Ke Xue Yuan Xue Bao,2005,27(6):665-669
3 Gang EJ,Jeong JA,Han S,et al.In vitro endothelial potential of human UC blood derived mesenchymal stem cells[J].Cytotherapy,2006,8(3):215-227
4 Jeon ES,Lee IH,Heo SC,et al.Mesenchymal stem cells stimulate angiogenesis in a murine xenograft model of A549 human adenocarcinoma through an LPA1 receptor-dependent mechanism[J].J Biochim Biophys Acta,2010,1801(11):1205-1213
5 Lenna S,Townsend DM,Tan FK,et al.HLA-B35 upregulates endothelin-1 and downregulates endothelial nitric oxide synthase via endoplasmic reticulum stress response in endothelial cells[J].J Immunol,2010,184(9):4654-4661
6 Kumar A,Kremer KN,Dominguez D,et al.Galpha13 and Rho mediate endosomal trafficking of CXCR4 into Rab11+ vesicles upon stromal cell-derived factor-1 stimulation[J].J Immunol,2011,186(2):951-958
7 Nishigaki A,Okada H,Okamoto R,et al.Concentrations of stromal cell-derived factor-1 and vascular endothelial growth factor in relation to the diameter of human follicles[J].J Fertil Steril,2011,95(2):742-746
8 Easwaran V,Lee SH,Inge L,et al.Beta-Catenin regulates vascular endothelial growth factor expression in colon cancer[J].Cancer Res.2003,63(12):3145-3153
9 Van Doorn R,Van Horssen J,Verzijl D,et al.Sphingosine 1-phosphate receptor 1 and 3 are upregulated in multiple sclerosis lesions[J].J Glia,2010,58(12):1465-1476
10 Sun X,Ma SF,Wade MS,et al.Functional variants of the sphingosine-1-phosphate receptor 1 gene associate with asthma susceptibility[J].J Allergy Clin Immunol,2010,126(2):241-249
猜你喜欢
中南药学(2022年2期)2022-11-14
体育科技文献通报(2022年4期)2022-10-21
世界科学技术-中医药现代化(2022年3期)2022-08-22
中国典型病例大全(2022年9期)2022-04-19
中国循证心血管医学杂志(2022年1期)2022-03-15
保健与生活(2021年24期)2021-12-12
现代临床医学(2021年5期)2021-11-02
昆明医科大学学报(2021年4期)2021-07-23
医学新知(2019年4期)2020-01-02
扣篮(2018年7期)2018-07-26