
2010年西藏小麦条锈菌生理小种群体结构与分析
2012-02-28 07:48:00王保通李佼佼胡小平康振生杨敏娜刘太国陈万权
植物保护 2012年2期
王保通, 李佼佼, 胡小平*, 李 强,康振生, 杨敏娜, 刘太国, 陈万权
(1.旱区作物逆境生物学国家重点实验室/西北农林科技大学,杨凌 712100;2.西藏农牧学院植物科学技术学院,林芝 860000;3.中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193)
西藏地区位于我国西南边陲,青藏高原的西南部,平均海拔4 000 m以上,南隔喜马拉雅山脉与印度、尼泊尔、锡金、不丹、缅甸等国接壤。北部和东部与青海、四川、云南等省区为邻。由于地理位置等原因,长期以来,人们认为该地为一个相对独立的农业生态区域。1990-1993年期间,王宗华、李晓忠等[1-4]对西藏小麦条锈病进行了阶段性研究,基本弄清了西藏小麦条锈病区域流行规律,越夏越冬规律、品种抗病性变异原因,开展了综合防治技术等方面的研究。但长期以来,由于西藏地区地理位置和交通等方面的原因,我国小麦条锈病协作组一直未对该地区的小麦条锈菌生理小种进行系统监测[6-10]。截至目前,对该地区小麦条锈菌生理小种群体结构组成的研究报道甚少,仅有的研究也由于采集标样数少而代表性可能不足[5]。小麦条锈病是一种气流性远距离传播的病害,监测西藏地区小麦条锈菌生理小种现状,研究西藏地区与内地小麦条锈菌生理小种种群之间的关系,对于完善我国小麦条锈病流行区域划分和条锈病大区流行规律,指导西藏地区小麦抗病育种和小麦条锈病防治具有重要意义。
1 材料和方法
1.1 采样地点
2010年8月,作者与西藏地区农牧学院在西藏自治区小麦条锈病发生最适流行区的林芝地区共采集小麦条锈病标样528份,其中繁殖成活了316份,鉴定并得出结果的标样为261份。采样点主要分布在西藏主要小麦种植区的林芝地区境内,其他地区由于没有发病均未采到标样。由于林芝地区是西藏自治区小麦条锈病发生最重,并可周年循环的地区,这次采样地点囊括了林芝地区不同海拔和生态条件下的条锈病标样,标样采集量是历史上最大的一次,因此,此次采集的标样基本能够代表西藏地区小麦条锈菌种群结构。
1.2 鉴别寄主
参试的鉴别寄主有‘Trigo Eureka’、‘Ful har d’、‘Lutescenes 128’、‘南大2419’、‘维尔’、‘阿勃’、‘早洋’、‘阿夫’、‘丹麦1号’、‘尤皮Ⅱ号’、‘丰产3号’、‘洛夫林13’、‘抗引655’、‘水源11’、‘中四’、‘洛夫林 10’、‘Hybrid 46’、‘Triticu m spelta albu m’和‘贵农22’。均来自西北农林科技大学植保学院植物免疫室。
1.3 菌种繁殖及小种鉴定
采用中华人民共和国农业行业标准(NY/T 1443.1-2007)小麦抗病虫性评价技术规范:第一部分小麦抗条锈病评价技术规范中“小麦条锈病病原菌和生理小种鉴定”方法。
1.4 反应型分级
反应型记载依据0、0;、1、2、3、4共6级。0~2级为抗病(R),3~4级为感病(S)。
2 结果与分析
2.1 西藏地区小麦种植基本情况及小麦条锈病发生状况
小麦是西藏的主要粮食作物之一,面积和产量仅次于青稞,居第2位。2010年,西藏小麦播种面积为36 770 h m2,其中冬小麦种植面积28 270 h m2,占小麦播种面积的76.88%。主要分布在拉萨市、山南地区、林芝地区、日喀则地区和昌都地区,冬小麦播种面积分别为8 780 h m2(占80%)、8 140 h m2(占 96%)、7 410 h m2(占 97%)、6 250 h m2(占42%)和6 280 h m2(占53%)。阿里地区和那曲地区只有极少量春小麦,不种植冬小麦。据调查,2010年西藏自治区小麦条锈病在全区属中度偏轻发生,主要发生在林芝地区冬小麦上,发生面积2 620 h m2,其余地区没有发生或零星发生,未采到条锈病标样。
2.2 西藏地区小麦条锈菌生理小种监测结果
鉴定结果表明(见表1):在261份标样中,共鉴定出19个已知小种类型,分别为CYR17、CYR20、CYR21、CYR22、CYR23、CYR29、CYR31、CYR32、CYR33、Lov13-6、Su11-1、Su11-2、Su11-3、Su11-4、Su11-5、Su11-6、Su11-7、Su11-8、Su11-13,它们出现的频率分别为 4.46%、4.46%、0.77%、0.38%、3.83%、1.15%、5.36%、31.80%、24.14%、0.38%、4.60%、0.38%、1.92%、2.30%、6.90%、0.38%、0.38%和0.77%;另外还有13份不能归类的标样,频率为4.98%。从鉴定结果来看,2010年我国西藏地区小麦条锈菌生理小种类型丰富,在鉴定并明确的生理小种(致病类型)中,出现频率最高的小种为CYR32,其出现频率为31.80%,其次为CYR33,出现频率为24.14%,而CYR31的频率仅为5.36%,CYR31号之前的小种有5个,分别为:CYR17、CYR20、CYR21、CYR22、CYR23和CYR29,其频率分别为4.60%、4.60%、0.77%、0.38%、3.83%和1.15%,处于相对较低的水平,洛13类群中除CYR29之外,仅监测到Lov13-6类型,出现频率为0.38%;Hybrid46类群中,除CYR32外,没有其他类型出现;而水源11类群中,除CYR33外,还有9个类型,它们分别为 Su11-1、Su11-2、Su11-3、Su11-4、Su11-5、Su11-6、Su11-7、Su11-8和 Su11-13,出现频率为 4.60%、0.38%、0.38%、1.92%、2.30%、6.90%、0.38%、0.38%和0.77%,其频率总和达42.15%,占有较大比例。表明我国西藏地区小麦条锈菌生理小种种群结构复杂,CYR32以前小种数较多,但主要优势小种以CYR32和CYR33为主,两者频率之和达到55.94%,超过了鉴定标样的一半以上,占绝对优势;水源11类群为优势类群,其频率之和为42.15%。
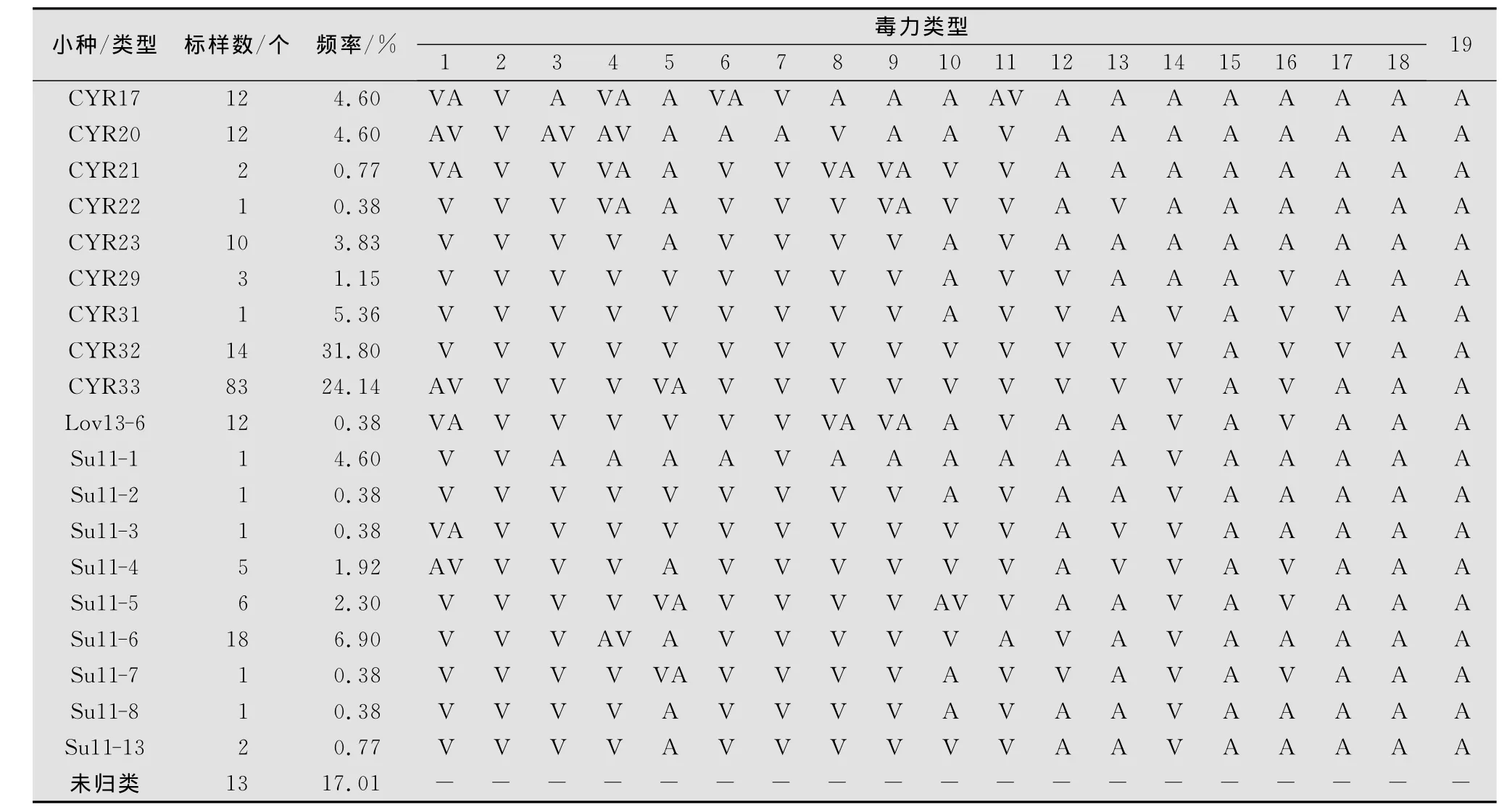
表1 2010年西藏地区小麦条锈菌生理小种监测结果1)
2.3 西藏地区小麦条锈菌毒性频率分析
从鉴定的261份标样结果看,西藏小麦条锈菌对小麦条锈病主要抗性基因有毒力的频率分别为:Yr1(‘丰 产 3 号’)为 95.16%、Yr3(‘丹 麦 1 号’)为85.48%、Yr3b/4b(‘Hybrid 46’)为 39.11%、Yr6(‘Trigo Eureka’)为95.16%,Yr9(‘洛夫林10’和‘洛夫林13’)为70.97%、Yr A(‘阿夫’)为90.32%。和全国平均水平相比(见表2),西藏条锈菌对以上已知条锈菌基因的毒力频率中,除Yr3、Yr9、Yr A略低于全国水平,其余对Yr1、Yr6和Yr3b/4b有毒力的频率与全国水平相当。

表2 西藏地区小麦条锈菌毒性基因频率与全国平均水平比较 %
3 讨论
小麦条锈病是一个可远距离传播的大区性流行病害,我国历次小麦条锈病的大流行都是由于条锈菌新的生理小种产生并成为优势小种,从而导致主栽品种抗性“丧失”。因此,做好小麦条锈菌生理小种监测是一项长期的基础性研究工作,对小麦条锈病的预测预报、抗病育种和病害的持久治理均有重要作用。我国自1957年小麦条锈病协作组成立以来,一直不间断地开展了中国小麦条锈菌生理小种的监测和预报工作,成功地预测了我国多次条锈菌群体变异引起的条锈病大流行。然而,我国小麦条锈菌生理小种监测工作至今尚未涉及西藏自治区境内的条锈菌标样,对西藏小麦条锈病的研究,仅限于1990-1993年期间王宗华等的阶段性研究[1-5],之后则只有极少数报道[5]。
小麦条锈病是西藏地区小麦上的重要病害[6]。西藏地区由于地处远离内地省份的高海拔地区,与内地省份生态条件差异大,形成了相对独立的农业生态区。王宗华、李晓忠等[1-4]研究认为,条锈菌在西藏可以顺利越夏、越冬,独立完成周年循环,构成了一个以本地菌源为主的流行区域;而该区与其他流行区域的关系研究甚少。2010年邝文静[5]对采自西藏的59份条锈菌标样进行鉴定并与四川、云南两省的标样进行了比较,认为西藏地区小麦条锈菌生理小种主要为2种类型:水源11类群类型和CYR28以前的类型,其中以水源11致病类群类型为优势小种,出现频率较高的类型为Su11-5和Su11-2,而CYR28以前的小种出现频率之和高达32.35%,认为西藏地区小麦条锈菌与四川省和云南省有着菌源交流,但西藏地区小种演化的速度慢于内地。考虑到该研究采集标样较少,代表性可能不足。作者于2010年从西藏采集并鉴定了小麦条锈菌标样261份,且标样采集地点主要分布在西藏小麦条锈菌可周年循环的最适流行区—林芝地区。因此,本研究无论从标样采集数量和采样地点来看更具有代表性,研究结果表明,西藏地区小麦条锈菌群体结构复杂,生理小种丰富,其优势小种为CYR32和CYR33,优势小种类群为水源11类群,没有监测到Hybrid46类群中的其他类型;另外CYR32之前小种数多且频率较高,主要以CYR17、CYR20、CYR31为主。与周边省份比较,西藏地区小麦条锈菌与四川省、云南省的群体结构有很大的相似性,优势小种组成也基本相似,而与青海省的结果存在较大差异。该结果与邝文静研究结果基本一致,只是在一些小种类型的频率上有所差异,本次鉴定出的小种类型数更复杂,这可能是本次研究的标样数和采集地点更具代表性。与周边省份比较(2010年全国条锈菌监测结果,内部资料,尚未发表)认为,西藏地区小麦条锈菌与四川省和云南省有着密切的菌源交流,而与青海省菌源交流较少。但西藏地区小麦条锈菌组成也有其自身的独特性,表现在生理小种群体结构更复杂,小种总数明显多于四川和云南省,CYR32之前的小种数和出现频率也明显高于四川和云南省。表明西藏地区的小麦条锈菌生理小种变异相对滞后。原因可能是:一方面西藏地区小麦条锈病可在当地完成周年循环,外来菌源传播至西藏后,可在当地保存、繁殖,加之西藏地区种植的小麦品种使用的抗源也是从周边省份引进的,从抗源使用上滞后于周边省份;另一方面西藏地区由于自然原因,新的小麦品种推广缓慢,而很多过去的老品种至今仍在种植,这就造成了西藏地区小麦品种和抗源的多样性比内地更复杂,为CYR32以前的小种存活提供了适合的寄主,这也是西藏地区条锈菌生理小种群体复杂的重要原因。
从西藏地区小麦条锈菌毒性频率分析结果来看,西藏地区小麦条锈菌对小麦条锈病主要抗性基因的毒力频率与全国总体水平相当,对一些抗锈基因的毒力频率略低于全国水平,这从另一个角度也证明了西藏地区小麦条锈菌生理小种变异滞后于周边地区的结论。小麦条锈病是一个大区流行的病害,研究西藏地区小麦条锈菌群体结构,弄清西藏小麦条锈病发生与我国周边地区条锈病发生的关系,对于完善中国小麦条锈病流行体系和持续控制全国小麦条锈病的发生和危害具有重要的指导意义。
[1] 王宗华.条锈菌在西藏越夏越冬规律的初步研究[J].西南农业学报,1992,5(1):64-68.
[2] 王宗华.西藏大、小麦条锈病区域流行规律[J].西南农业大学,1992,5(2):79-84.
[3] 王宗华.西藏麦类条锈病发生规律及综合防治研究[J].西藏农业科技,1993,15(2):6-11.
[4] 王宗华,旺姆.小麦品种在西藏抗条锈性变异分析[J].西南农业学报,1992,5(4):88-92.
[5] 邝文静.西藏小麦条锈病菌群体多样性研究[D].雅安:四川农业大学,2010.
[6] 吴立人,杨华安,袁文焕,等.1985-1990年我国小麦条锈菌生理专化研究[J].植物病理学报,1993,23(3):269-274.
[7] 万安民,牛永春,吴立人,等.1991-1996年我国小麦条锈菌生理专化研究[J].植物病理学报,1999,29(1):15-21.
[8] 万安民,吴立人,贾秋珍,等.1997-2001年我国小麦条锈菌生理小种变化动态[J].植物病理学报,2003,33(3):261-266.
[9] Wan A M,Zhao Z H,Chen X M,et al.Wheat stripe rust epidemic and vir ulence of Puccinia striifor mis f.sp tritici in China in 2002[J].Plant Disease,2004,88(8):896-904.
[10]Chen W Q,Wu L R,Liu T G,et al.Race dynamics,diversity and virulence evolution in Puccinia striifor mis f.sp.tritici,the causal agentof wheat stripe rust in china from 2003 to 2007[J].Plant Disease,2009,93(11):1093-1101.
猜你喜欢
南方农业学报(2022年11期)2022-03-18 05:55:26
农业技术与装备(2021年12期)2021-12-02 17:24:28
成都信息工程大学学报(2017年5期)2018-01-23 02:45:52
林业科学(2017年7期)2017-08-30 14:24:16
植物保护(2017年4期)2017-08-09 01:18:38
中国马铃薯(2017年1期)2017-03-02 09:15:51
作物学报(2016年10期)2016-10-19 04:14:09
西藏科技(2016年10期)2016-09-26 09:02:06
陕西林业科技(2016年1期)2016-08-04 00:35:49
中国学术期刊文摘(2015年2期)2015-10-27 08:12:31
