
牙龈卟啉单胞菌侵入口腔上皮细胞后基因表达变化的初步研究
2012-02-05 01:48孔令雪吴亚菲
牙体牙髓牙周病学杂志 2012年11期
成 伟,王 洁,孔令雪,齐 霞,吴亚菲,赵 蕾
(四川成都610041:1.四川大学口腔疾病研究国家重点实验室;2.四川大学华西口腔医院牙周科)
牙龈卟啉单胞菌(Porphyromonas gingivalis,Pg)是一种可破坏牙周组织的G-专性厌氧菌,它广泛存在于慢性牙周炎患者的龈下菌斑中,是公认的牙周主要致病菌[1]。Pg对龈沟内牙龈上皮细胞的粘附和侵入作用是细菌引发牙周炎的关键性起始步骤之一。在单纯培养状态下与感染宿主细胞过程中,Pg表达的基因存在显著差异。提示该菌与牙龈上皮细胞的感染接触作用可能是诱导其多种致病基因表达的重要调控信号,能帮助Pg完成粘附及侵入、逃逸宿主防御攻击、诱导宿主细胞炎症介质表达等致病过程。KB细胞株是目前国际通用的牙龈上皮细胞替代细胞模型之一,其增殖频率和活性稳定,与Pg相互作用的方式类似于牙龈上皮细胞[2-6],被广泛应用于牙周致病菌与宿主细胞的相互调控研究中。本研究利用 Pg ATCC 33277攻击KB细胞,采用差异显示反转录PCR技术(differential display reverse transcription PCR,DDRT-PCR),比较Pg侵入KB细胞后与纯培养状态下细菌全基因组的基因表达变化,初步筛选并分析与细菌侵入宿主细胞相关的毒力致病基因,以期为后续Pg特异性毒力因子的明确和功能分析提供依据。
1 材料和方法
1.1 菌株、细胞株和主要试剂、仪器
Pg ATCC 33277、KB细胞株ATCC CCL17(四川大学口腔疾病研究国家重点实验室);BHI培养基(OXIOD,英国);细菌RNA提取试剂盒Ribo-PureTM-Bacteria Kit(Ambion公司,美国);引物、cDNA逆转录试剂盒PrimeScriptTMRT reagent Kit(大连TaKaRa公司);2×Taq PCR MasterMix PCR试剂盒(北京全式金生物技术有限公司);梯度PCR仪(Eppendorf,德国);Gel-Doc2000凝胶成像观察系统(Bio-Rad,美国);透射电镜(H-600IV HITICHI,日本)。
1.2 牙龈卟啉单胞菌与KB细胞共培养
取Pg ATCC 33277常温复苏后,接种于BHI血琼脂培养基(含5%冻溶兔血、1%血红素和维生素K),37℃厌氧(800 mL/L N2、100 mL/L H2、100 mL/L CO2)培养48 h。革兰染色和生化鉴定为纯培养后,挑取菌落接种于BHI液体培养基继续厌氧培养24 h。培养物经6 000 r/min离心8 min去上清,用不含胎牛血清和双抗的低糖DMEM培养基重悬,麦氏比浊仪调整细菌悬液浓度至1×108CFU/mL备用。常规复苏KB细胞株ATCC CCL17并接种于低糖DMEM(100 mL/L FBS+1%青霉素/链霉素)中,饱和湿度、50 mL/L CO2、37℃标准环境中培养并传代1~2代,镜下观察细胞形态呈“铺路石”状,在长满瓶底80%时,用含EDTA的2.5 g/L胰蛋白酶消化,调整细胞密度为2×105/mL,以每孔2 mL接种于6孔板继续培养。24 h后每孔细胞数增殖为5×105/孔,将上述配置好的Pg菌悬液(1×108CFU/mL)1 mL加入6孔板各孔内,使细菌攻击细胞MOI值为200∶1,标准条件下共同孵育18 h并经透射电镜观察确认Pg成功侵入KB细胞后,PBS清洗3次以去除未粘附Pg,收集细胞备用。
1.3 Pg总RNA的提取和cDNA的合成
常规厌氧培养18 h的Pg ATCC 33277菌液调整浓度为1×108CFU/mL,取1mL菌液8 000 g 4℃离心2 min后弃上清,PBS清洗3次后备用;将已去除未粘附细菌的KB细胞用去离子冰水裂解20 min,加入含1 mmol/L MgCl2、0.5 mg/mL DNA酶和0.25 mg/mL RNA酶的PBS液浸泡30 min,去除KB细胞的DNA和RNA,收集样本于1 mL的EP管中。上述样本用RiboPureTM-Bacteria Kit细菌RNA提取试剂盒按照说明书操作步骤分别提取纯培养和侵入KB细胞的Pg的总RNA。10 g/L的琼脂糖电泳(90 V,80 mA)检测RNA质量。利用PrimeScriptTMRT reagent Kit试剂盒,根据说明书逆转录细菌RNA成cDNA,于-20°冻存备用。
1.4 差异显示反转录DDRT-PCR(表1)
选取不同的锚定引物和随机引物配置成不同组合(共12种)进行PCR反应。PCR总反应体系为:4μL 模板、锚定引物和随机引物(0.01 mmol/L)各1 μL、MIX酶12.5 μL、无菌三蒸水补足体积至25 μL。将细菌攻击细胞前后的cDNA作为一组进行反应。PCR反应在PCR热循环扩增仪完成,PCR程序为:94℃ 60 s,94℃ 60 s,40℃120 s,72℃60 s,1个循环;94℃45 s,60℃ 120 s,72℃120 s,35个循环;72℃7 min延伸。

表1 DDRT-PCR所用的锚定引物及随机引物序列
1.5 差异条带回收、二次扩增、测序和BLAST分析
取DDRT-PCR产物各15 μL,以100 bp为分子量标记,20 g/L的琼脂糖凝胶做电泳分析,显示差异表达片段。将差异片段按照胶回收试剂盒进行cDNA回收,将回收得到的cDNA应用相应引物再次进行DDRT-PCR扩增,扩增产物送至宝生物工程(大连)有限公司测序。所获得条带的cDNA序列数据,在GENEBANK中与Pg ATCC 33277和Pg W83的基因组已知核苷酸序列进行比对,行BLAST同源分析(http://www.ncbi.nlm.nih.gov/ sutils/genom_table.cgi,http://www.oralgen.lanl.gov/)。
2 结果
2.1 侵入KB细胞后Pg ATCC 33277 RNA提取结果(图1)
侵入KB细胞前后Pg ATCC 33277的RNA样品电泳条带清晰,泳道上无明显弥散痕迹,其中23S∶16S rRNA条带亮度比值约为2∶1,说明总RNA提取完整无降解,满足后续实验要求。
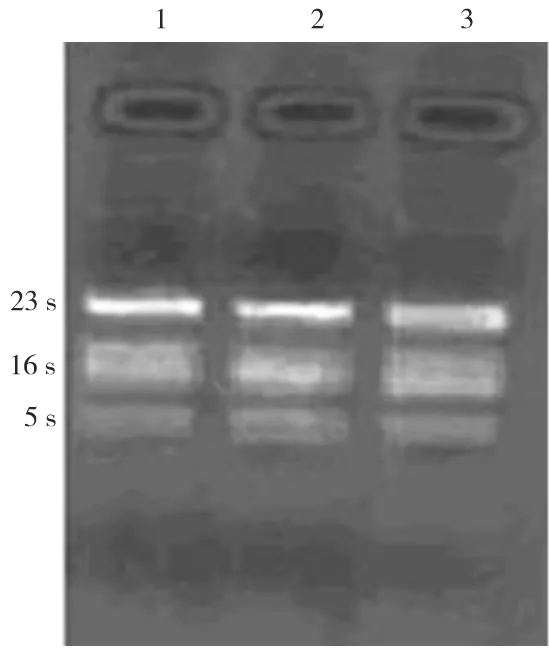
图1 侵入KB细胞前后Pg总RNA的电泳结果
2.2 差异显示反转录DDRT-PCR的电泳结果(图2)
侵入KB细胞后Pg ATCC 33277的cDNA经DDRT-PCR扩增后均得到清晰的扩增条带。其中第1、2泳道的扩增引物为A*ARP2;3、4泳道为C*ARP2;5、6泳道为C*ARP4。泳道1、3、5为攻击细胞后Pg cDNA扩增产物,2、4、6为纯培养Pg cDNA扩增产物。电泳结果显示:与2泳道相比较,1泳道存在差异条带HX 01;与4泳道比较,3泳道存在差异条带HX 02,与6泳道相比,5泳道存在差异条带HX 03。
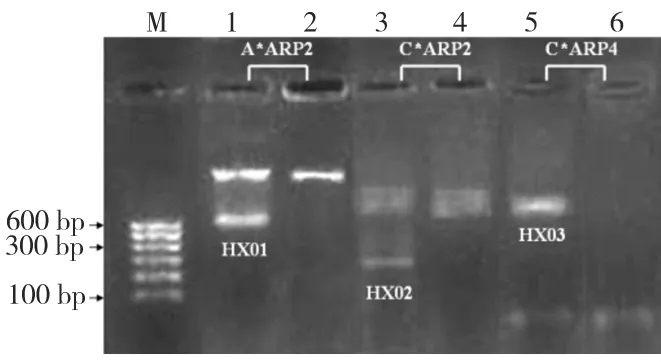
图2 Pg ATCC 33277侵入KB细胞前后DDRT-PCR结果
2.3 差异片段的BLAST序列分析
差异条带cDNA应用相应引物二次扩增后,扩增产物测序结果显示:PGHX 01序列共645 bp,经BLAST分析发现,PGHX01与Pg 33277的PG0683序列有96%的基因同源性,PG 0683编码ABC转运蛋白(ATP-binding cassette transporters);PGHX 02序列共351 bp,BLAST分析与PG 0159存在97%基因同源性,PG0159编码Pg PepO;PGHX03序列共738 bp,BLAST分析结果显示未发现类似基因序列片段(表2)。

表2 DDRT-PCR差异表达片段的BLAST同源性分析结果
3 讨论
牙龈卟啉单胞菌是慢性牙周炎的重要致病菌之一,具有多种毒力因子,细菌对宿主细胞的粘附和侵入能力是衡量其致病力的重要指标之一。研究显示[7-9]:Pg在与牙龈上皮屏障接触的15 min之内即可粘附并侵入宿主细胞;胞内细菌通过抑制细胞凋亡为自身创造有利于胞内生存的时间和空间条件,侵入24~48 h后达到胞内复制高峰;并通过调节细胞自吞噬循环通路逃离原驻细胞,转而攻击临近未感染细胞。Papapanou等[10]应用基因芯片分析了炎症位点牙周组织的基因表达变化与11种牙周可疑致病菌检出水平之间的相关关系,发现:8 537个芯片探针组相关基因的变化与牙龈卟啉单胞菌的侵入存在相关性。侵入细胞的Pg不仅能够调控基因的表达,还可通过影响信号传导通路和microRNA来控制细胞因子的表达和细胞凋亡。Mao等[11]的研究显示:Pg感染牙龈上皮细胞可导致细胞内JAK1和STAT3的磷酸化,控制细胞内的线粒体细胞死亡途径JAK/STAT通路,阻止宿主细胞的程序死亡,从而提高牙龈卟啉单胞菌在牙龈上皮细胞内生存。Moffatt等[12]发现:侵入牙龈上皮细胞的Pg可通过诱导microRNA-203来影响细胞因子信号3抑制子的表达。以上研究均提示:牙龈上皮细胞是Pg攻击的主要靶目标,是牙周病变发生发展的始发部位之一。本研究选用KB细胞替代牙龈上皮细胞与Pg共孵育,模拟体内Pg感染宿主牙龈上皮的初始状态。透射电镜观察显示:Pg可成功侵入KB细胞,为后续分析Pg侵入上皮细胞后毒力基因的表达奠定实验基础。
同宿主真核细胞的相互接触是诱导细菌特异性感染毒力因子表达的重要调控信号。Hendrickson等[13]的研究即显示:Pg与牙龈上皮细胞共培养后可特异性上调表达396条蛋白。Rodrigues等[14]也发现:Pg侵入冠状动脉内皮细胞后出现了62条编码能量代谢转移调控、细胞壁信封结构、核酸及蛋白修饰加工等蛋白的基因表达变化。以上结果均提示:Pg粘附和侵入上皮细胞后可能表达某些特异性毒力成分,从而干扰细胞正常结构和生理功能。基因的差异性表达是细菌、细胞形态和功能变化的根本原因,也是各种生理及病理过程发生的物质基础。从全基因组角度研究Pg粘附和侵入宿主上皮细胞过程中细菌基因表达谱的变化,对于宏观揭示Pg特异性致病机制具有重要意义。目前Pg W83和Pg ATCC 33277全基因组测序和部分基因功能鉴定已成功完成,为深入分析Pg差异表达基因的功能提供了依据。Tachibana等[15]已运用减杂交技术比较了纯培养状态下Pg ATCC 33277和Pg W83的差异表达基因,发现Pg W83具有5个不同于ATCC 33277的特异性毒力相关基因。本研究应用差异显示反转录技术(DDRT-PCR)从全基因组分析纯培养状态和侵入KB细胞后的Pg ATCC 33277的基因表达差异,通过3个锚定引物和4个M13随机引物的随机组合扩增cDNA后,成功获得3条差异基因条带。其基因序列编号为:PGHX 01、PGHX 02、PGHX 03。进行测序和BLAST同源性比对分析,结果显示:PGHX 01与Pg 33277的PG 0683基因序列有96%同源性。PGHX 02与Pg 33277的Pg 0159基因序列有97%同源性。PGHX 03未发现类似同源性基因序列。PGHX 01基因序列与编码 ABC转运蛋白(ATP-binding cassette transporters)基因序列 PG 0683[16-19]具有较高的同源性。ABC转运蛋白是一种高度保守性的跨膜蛋白,广泛存在于各个物种中。ABC转运蛋白与肿瘤耐药、囊肿性纤维化、高密度脂蛋白缺乏症等相关;并与微生物生物膜形成、粘附功能、细菌耐药性等存在相关性。Pg W83基因组中共有39个不同基因ABC转座子已被确定[20]。研究显示:ABC转运蛋白参与了Pg血红素的转运、结合与吸收;敲除某些编码ABC转运蛋白的基因可影响Pg生物膜的形成、对宿主细胞的侵入、并参与调控LPS和细菌分泌系统的合成。本研究中,与ABC转运蛋白编码基因PG 0683高度同源性的PGHX 01基因出现表达,再次证实ABC转运蛋白在Pg感染牙龈上皮细胞的过程中发挥重要的毒力作用。PGHX 02基因序列与编码Pg PepO基因序列PG 0159高度同源。Pg PepO为一种基质金属蛋白酶,与人内皮素转换酶(ECE)-1有显著同源性,人内皮素转换酶-1可以转换大内皮素 -1到内皮素 -1[21]。内皮素 -1水平升高,与动脉粥样硬化和心脏衰竭的发病有关。在Pg中出现PepO部分揭示了牙周炎与动脉粥样硬化相关性的微生物学机制[22-23]。Ansai等[24]研究发现缺乏Pep0基因的细菌变异型缺乏内肽酶活性,其对上皮的侵入效能仅为野生型的四分之一,提示Pep0在侵入细胞和细胞膜的溶解过程中发挥了一定作用。本研究所得PGHX 03基因经BLAST同源性比对,未发现类似基因。其可能为一种未知基因,编码未知功能蛋白,目前尚无报道,有待于进一步的研究。
[1] Holt SC,Ebersole JL.Porphyromonas gingivalis,Treponema denticola,and Tannerella forsythia:the“red complex,”a prototype polybacterial pathogenic consortium in periodontitis[J].Periodontol,2000,2005,38:72-122.
[2] Sugano N,Ikeda K,Oshikawa M,et al.Differential cytokine induction by two types of Porphyromonas gingivalis[J].Oral Microbial Immunol,2004,19(2):121-123.
[3] Yilmaz 0,Watanabe K,Lamont RJ.Involvement of integrins in fimbriae mediated binding and invasion by Porphyromonas gingivalis[J].Cell Microbiol,2002,4(5):305-314.
[4] Watanabe K,Yilmaz O,Nakhjiri SF,et al.Association of mitogen-activated protein kinase pathways with gingival epithelial cell responses to Porphyromonas gingivalis infection[J].Infect Immun,2001,69(11):6731-6737.
[5] Nakhjiri SF,Park Y,Yilmaz O,et al.Inhibition of epithelial cell apoptosis by Porphyromonas gingivalis[J].FEMS Microbiol Lett,2001,200(2):145-149.
[6] 郭永华,吴亚菲,刘天佳,等.两种fimA基因型牙龈卟啉单胞菌刺激下口腔上皮细胞ICAM-1的表达[J].北京口腔医学,2008,16(5):241-243.
[7] Yilmaz O,Verbeke P,Lamont RJ,et al.Intercellular spreading of Porphyromonas gingivalis infection in primary gingival epithelial cells[J].Infect Immun,2006,74(1):703-710.
[8] Yilmaz O,Jungas T,Verbeke P,et al.Activation of the phosphatidylinositol 3-kinase/Akt pathway contributes to survival of primary epithelial cells infected with the periodontal pathogen Porphyromonas gingivalis[J].Infect Immun,2004,72(7): 3743-3751.
[9] Takeuchi H,Furuta N,Morisaki I,et al.Exit of intracellular Porphyromonas gingivalis from gingival epithelial cells is mediated by endocytic recycling pathway[J].Cell Microbiol,2011,13(5):677-691.
[10] Papapanou PN,Behle JH,Kebschull M,et al.Subgingival bacterial colonization profiles correlate with gingival tissue gene expression[J].BMC Microbiol,2009,9:221.
[11] Mao S,Haseqawa Y,Tribble GD,et al.Intrinsic apoptotic pathways of gingival epithelial cells modulated by porphyromonas gingivalis[J].Cell Microbiol,2007,9(8):1997-2007.
[12] Moffatt CE,Lamont RJ.Porphyromonas gingivalis induction of MicroRNA-203 expression controls suppressor of cytokine signaling 3 in gingival epithelial cells[J].Infect Immun,2011,79 (7):2632-2637.
[13] Hendrickson EL,Xia Q,Wang T,et al.Pathway analysis for intracellular Porphyromonas gingivalis using a strain ATCC 33277 specific database[J].BMC Microbiol,2009,9:185.
[14] Rodrigues PH,Progulske-Fox A.Gene expression profile analysis of Porphyromonas gingivalis during invasion of human coronary artery endothelial cells[J].Infect Immun,2005,73(9): 6169-6173.
[15] Tachibana-Ono M,Yoshida A,Kataoka S,et al.Identification of the genes associated with a virulent strain of Porphyromonas gingivalis using the subtractive hybridization technique[J].Oral Microbiol Immunol,2008,23(1):84-87.
[16] Jones PM,George AM.The ABC transporter structure and mechanism:perspectives on recent research[J].Cell Mol Life Sci,2004,61(6):682-699.
[17] Joni T,Glenn ST.ABC transporters in microorganisms[J].Expert Rev,2010,8(4):375-377.
[18] Davidson AL,Dassa E,Orelle C,et al.Structure,function,and evolution of bacterial ATP-binding cassette systems[J].Microbiol Mol Biol Rev,2008,72(2),317-364.
[19] Goffeau A,Hertogh B,Baret PV.ABC transporters[M].In: Encyclopedia of Biological Chemistry,2004,1,1-5.
[20] Park Y,Yilmaz O,Jung IY,et al.Identification of porphyromonas gingivalis genes specifically expressed in human gingival epithelial cells by using differential display reverse transcription-PCR[J].Infect Immun,2004,72(7):3752-3758.
[21] Awano S,Ansai T,Mochizuki H,et al.Sequencing,expression and biochemical characterization of the Porphyromonas gingivalis pepO gene encoding a protein homologous to human endothelin-converting enzyme[J].FEBS Lett,1999,460(1):139-144.
[22] Beck J,Garcia R,Heiss G,et al.Periodontal disease and cardiovascular disease[J].J Periodontol,1996,67(10):1123-1137.
[23] Beck JD,Offenbacher S.Oral health and systemic disease:periodontitis and cardiovascular disease[J].J Dent Edu,1998,62(10):859-870.
[24] Ansai T,Yu WS,Urnowey S,et al.Construction of a pepO gene-deficient mutant of Porphyromonas gingivalis:potential role of endopeptidase in the invasion of host cells[J].Oral Microbiol Immunol,2003,18(6):398-400.
猜你喜欢
计算机应用与软件(2022年6期)2022-07-12
南京师大学报(自然科学版)(2022年2期)2022-06-20
油气地质与采收率(2022年3期)2022-05-20
自然灾害学报(2022年2期)2022-05-10
小哥白尼(趣味科学)(2021年10期)2022-01-17
中国学校体育(2021年10期)2021-04-26
学生天地(2020年10期)2020-08-25
大众科学(2018年1期)2018-03-28
科技知识动漫(2017年12期)2018-03-07
儿童故事画报·发现号趣味百科(2015年10期)2016-01-20
