
陕北野生山丹丹染色体数目与核型分析研究
2012-01-25 05:07丁媛媛齐向英陈宗礼
中国野生植物资源 2012年2期
丁媛媛,齐向英,陈宗礼,刘 维,杨 鹏
(延安大学生命科学学院陕西省红枣重点实验室(延安大学)陕西省区域生物资源保育与利用工程技术研究中心,陕西延安716000)
山丹丹(Lilium pumilum DC.)为百合科,百合属多年生草本植物[1]。主要分布在陕西、宁夏、山西、甘肃、青海等地。在陕北延安地区分布的山丹丹一般株高30~40 cm,叶狭长如松叶,花具清香,春末夏初开放。其花朵下垂,花瓣向外反卷,通常无斑点有光泽,十分美丽,是极好的园艺观赏花卉和切花材料。其生命力顽强,花色火红,是陕北地区的代表性花卉,在陕北地区常被视为中国红色革命的象征。山丹花具有极高的药用价值。《本草纲目》中记载山丹丹花味微苦,性平,有养阴润肺、清心安神功能。现代医学研究表明山丹丹不仅用于阴虚久咳,虚烦惊悸,而且是很好的药食同源植物。国外对野生百合资源的研究较早,英国在16世纪末期就开始了野生百合的调查、收集和评价[2],国内也对部分百合属 植 物 如 对 毛 叶 百 合(L.dauricum Ker.gewl.)[3-15]、卷丹(L.lancifolium Thunb.)[4-5]、条叶百合(L.callosum Sieb.et Zucc.)[6]、紫斑百合(L.nepalense D.Don.)[7]等进行了核型研究,但关于山丹丹,尤其是陕北野生山丹丹的核型至今尚未见报道。本文利用酸解去壁低渗法首次对陕北地区野生山丹丹染色体数目和核型进行报道,旨在了解其特性,为山丹丹的品种选育及栽培利用提供细胞学基础。
1 材料与方法
1.1 材料与试剂
1.1.1 供试材料 采自陕西省区域生物保育与利用工程技术研究中心苗木品种园示范园保存的野生山丹丹,原种采自延川县小程村。
1.1.2 试剂 饱和对二氯苯,无水乙醇,冰醋酸,氯化钾,改良苯酚品红染液,盐酸。
采用Olympus BX41 DP 71全自动显微照相系统。
1.2 方法
1.2.1 染色体制片方法 取材:于上午8:30~9:30进行。从山丹丹根部取长约1~2 cm幼嫩根尖。预处理:饱和对二氯苯溶液前处理4 h。固定:卡诺氏固定液(无水乙醇:冰醋酸体积比为3∶1)中固定12 h。前低渗:0.075 mol/L氯化钾溶液中前低渗1h。酸解:放入预热好的0.1mol/L盐酸中60℃恒温水浴解离7 min。后低渗:0.075 mol/L氯化钾溶液中后低渗30 min。染色:改良石碳酸品红染液染色30 min。压片:取分生区的材料置于载玻片上,滴一滴染液压片。镜检拍照:将制好的染色体临时装片置于Olympus BX41 DP71全自动显微镜下观察,捕捉图像,寻找染色清晰、分散而不重叠的分裂中期细胞进行拍照。
1.2.2 染色体数目确定与核型分析方法 随机选取30个染色体清晰且分散良好的中期分裂相细胞进行染色数目统计。分别对染色体数目及含有不同染色体数目的细胞数进行统计,计算其在观察细胞的总数中的百分比和95%置信区间。
冲洗5张放大的照片,剪下染色体测量长度,将同源染色体进行配对。核型分析按李懋学(1985)的分类标准进行[8],核型分类根据Stebbins(1971)的分类标准划分[9],核型不对称系数(As.K/%)按Arano的方法计算[10]。染色体的相对长度、臂长及类型参照Levan等的命名系统命名[11]。
染色体形态测量公式:
相对长度=(染色体长度/染色体组总长度)×100%
着丝粒指数=(短臂/染色全长)×100%
臂比=长臂/短臂
核型不对称系数(As.K%)=(长臂总长/全组染色体总长)×100%
2 结果与分析
2.1 染色体数目
通过对30个细胞分裂相的观察,结果见表1。含24条染色体的细胞共计27个,占观察细胞总数的90.00%,95%置信区间在79.26% ~100.74%,大于24条染色体的细胞有3个,占观察细胞总数的10%,其中有1个细胞含有25条染色体,占总数的3.33%,2个细胞含有26条染色体,占观察总数的6.67%。
2.2 山丹丹染色体长度与核型
山丹丹的中期染色体形态见图1,山丹丹染色体及核型图见图2,核型模式图见图3。据表2可以看出,山丹丹有24条染色体,相对长度变化范围在5.84%~12.75%,全组染色体总长度为100.01 μm,平均长度为8.334 μm,属于大型染色体。最长染色体与最短染色体之比为2.183,臂比大于2∶1的染色体有3、4、5、6、7、9、11、12 ,8 组,占全组染色体的66.67%。按Stebbins的核型不对称标准,属于“3B”型。核型不对系数为72.16%,核型对称性差,为较不对称类型。根据Stebbins有花植物核型进化中对称—原始,不对称—进化的观点[12],陕北野生山丹丹属于较进化类型。配对的12组染色体中,第1、2、8、10 组为中部着丝点染色体(m),第6组为近中部着丝点染色体(sm),第 3、4、5、7、9、12 组为近端部着丝点染色体(st),11组为端部着丝点染色体(t)。其中在第10组,第11组染色体短臂上发现具有随体。根据以上数据,可以确定野生山丹丹的染色体核型公式为2n=2x=8m+2sm+12st+2t。

表2 山丹丹染色体相对长度、臂比与类型

图1 山丹丹的中期染色体形态
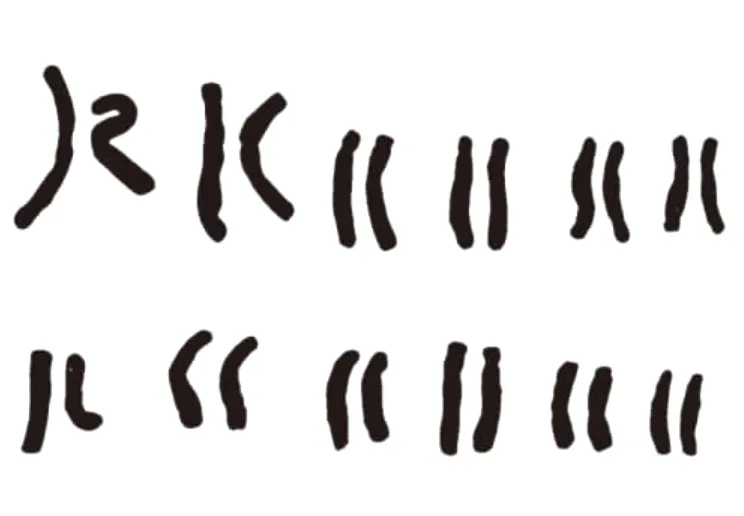
图2 山丹丹染色体核型图

图3 山丹丹染色体核型模式图
3 讨论
每一种生物染色体数目是恒定的,我们通过对30个细胞分裂相的观察得到,含24条染色体的细胞为27个,占观察总细胞数的90%,95%的置信区间结果为79.26~100.74。根据陈瑞阳[13]等植物染色核型分析标准的要求,最少统计30个细胞,其中85%以上细胞具有恒定一致的染色体数,我们的研究结果正好在该标准要求的范围内,所以认为山丹丹染色体是24条。
核型是指染色体组在有丝分裂中期的表现,是染色体数目、大小、形态特征的总和。在对染色体进行测量计算的基础上进行分组、排队、配对并进行形态分析的过程叫核型分析。百合属植物的染色体多为大型,染色体核型一般为“3B”型[14],染色体基数为12,以二倍体居多,目前仅在卷丹[15-16]、川百合[17]等少数种中发现三倍体。图立古尔等[18]对吉林省产的5种百合品种进行了核型分析,结果表明5种百合的核型为“3A”和“3B”型,戴小红等[19]对观赏百合3个品种的核型分析表明,其核型类型也为“3A”和“3B”型,本研究得到山丹丹的核型属于“3B”型,与他们的研究结果一致。在植物体细胞染色体鉴别中,主要是以其全长、臂长、着丝点位置和特殊的结构如随体为依据的。我们的研究在野生山丹丹的染色体中发现具有居间随体,分别位于第10对,11对染色体的短臂上。这与刘华敏等[20]报道的山丹(L.pumilum DC.)第1、6、8 对染色体上有居间随体,第2对染色体的短臂上有随体,第11对染色体的长臂上有随体,范小峰等[21]报道的细叶百合无随体出现,岳玲等[16]报道的细叶百合有1对染色体有随体的结果有所不同。
本研究对山丹丹的核型分析结论与刘华敏等[20]报道的山丹(L.pumilum DC.)核型为 2n=2x=2m+6sm+4st+12t,属3A型结论不一致,这与百合属各种间植物的生长环境有关。如果排除试验误差因素的干扰,可能是因为山丹产地的不同,为了适应不同的环境条件,植物染色体及其外部形态发生了一定程度的变异。刘敏华等所用材料来源于甘肃天水的野生山丹,其生长环境与陕北延安地区相比差异较大,生境不同将会引起不同生态类型,这将会造成其核型的差异。
[1] 汪劲武.种子植物分类学[M].北京:高等教育出版社,2009:253-254.
[2] Hubert B,Drysdale W,Wllliam T S.Lilies of theWorld[M].London:Country Life Limited,1950:17-19.
[3] 唐翠.毛百合的核型分析[J].哈尔滨师范大学自然科学学报,2008,24(3):85-88.
[4] 杨利平,刘雪梅,张敩芳.卷丹的细胞学研究[J].植物研究,1997,17(1):85-87.
[5] 李卫民,孟宪纡,高英,等.中药百合的核型分析[J].中国中药杂志,1991,16(5):268-270.
[6] 邵建章,张定成,杨连珍,等.安徽百合属细胞学研究[J].安徽师范大学学报:自然科学版,1994,17(2):39-43.
[7] 李标,虞泓,唐坤.紫斑百合居群核型变异式样[J].重庆邮电学院学报:自然科学版,2004,16(1):98-102.
[8] 李懋学,张赞平.作物染色体及其研究技术[M].北京:中国农业出版社,1996:1-37.
[9] Stebbins G L.Chromosomal Evolution in Higher Plants[M].London:Edward Amold Ltd,1971:85-104.
[10] Arano H.Cytological studies in subfamily Carduoides(Compositae)of Japan[J].Bot Mag(Takyo),1963,76:32 -39.
[11] Levan A,Fregdga K,Sandberg A A.Nomenclature for centromeric position on chromosomes[J].Hereditas,1964,52:201 -220.
[12] Stebbins G L.植物的变异和进化[M].复旦大学遗传研究所译.上海:上海科学技术出版社,1950.
[13] 李懋学,陈瑞阳.关于核型分析的标准化问题[J].武汉植物学研究,1985,3(4):297 -302.
[14] 洪德元.植物细胞学分类[M].北京:科学出版社,1990:91-96.
[15] 杨利平,刘雪梅,张敩芳.卷丹的细胞学研究[J].植物研究,1997,17(1):85-87.
[16] 岳玲,雷家军,王欣.辽宁的四种野生百合(Lilumu spp.)的核型研究[J].辽宁农业科学,2006(4):5-8.
[17] 谢晓阳,武全安.三倍体川百合的核型与酯酶同功酶鉴定[J].云南植物研究,1993,15(1):57-60.
[18] 图立古尔,刘立波.吉林省产的5种百合的核型研究[J].武汉植物学研究,1996,14(1):6 -12.
[19] 戴小红,牛立新,张延龙.百合三品系代表品种的核型分析[J].西北林学院学报,2006,21(4):58 -61.
[20] 刘华敏,智丽,赵丽华,等.四种野生百合核型分析[J].植物遗传资源学报,2010,11(4):469 -473.
[21] 范小峰,郭小强,李师翁.子午岭产4种百合科植物的核型多样性研究[J].西北植物学报,2000,20(5):882-888.
猜你喜欢
临床检验杂志(2021年10期)2021-11-24
川北医学院学报(2021年6期)2021-07-13
学生天地(2020年19期)2020-06-01
科学之谜(2019年3期)2019-03-28
人大建设(2018年6期)2018-11-17
科学之谜(2018年8期)2018-09-29
传记文学(2017年4期)2017-04-25
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
中国当代医药(2015年9期)2015-03-01
