
发芽蚕豆富集γ-氨基丁酸的培养液组分优化
2011-11-28 02:28:32杨润强韩永斌顾振新
中国粮油学报 2011年11期
陈 惠 杨润强 韩永斌 顾振新
发芽蚕豆富集γ-氨基丁酸的培养液组分优化
陈 惠 杨润强 韩永斌 顾振新
(南京农业大学农业部农畜产品加工与质量控制重点开放实验室,南京 210095)
以蚕豆为试材,研究谷氨酸钠(MSG)、CaCl2、和VB6对发芽蚕豆谷氨酸脱羧酶(GAD)及γ-氨基丁酸(GABA)的影响,采用Box-behnken设计对发芽蚕豆富集GABA的培养液组分进行了优化,并对发芽蚕豆富集GABA的二次回归模型进行分析。结果表明,低氧联合盐胁迫下,MSG、CaCl2和VB6对发芽蚕豆GAD及GABA的影响均达到显著水平(P<0.005)。经过回归分析建立了GABA含量对培养液组分的二次回归模型,回归方程的决定系数达到0.976,说明方程能很好的预测GABA富集含量的变化。蚕豆富集GABA的最适培养液组分为MSG 1.1 mg/mL、CaCl26.1 mmol/L、VB672 μmol/L,此时,GABA 富集量达到(1.98 ±0.09)mg/g DW,为对照[(1.08 ±0.01)mg/g DW]的1.83 倍。
发芽蚕豆 γ-氨基丁酸 富集 优化
蚕豆(Vicia faba L.)为豆科野豌豆属作物,其种子富含蛋白质、淀粉、氨基酸、维生素和钾、钙、磷等矿物质元素[1]。蚕豆种子中丰富的蛋白质和氨基酸可为γ-氨基丁酸(GABA)富集提供充足的底物。GABA具有镇静安神、抗焦虑、抗惊厥、降血压等作用[2]。通常植物组织中GABA含量较低,但受到热激、盐害和低氧等逆境胁迫时,其含量提高几倍至几十倍[3]。低氧胁迫导致植物细胞质酸化[4],使谷氨酸脱羧酶(GAD)活性提高,为GABA的合成提供了条件。盐胁迫使植物体胞内Ca2+浓度升高[5],CaM(钙调素)水平提高,GAD活性增加,从而促进GABA积累。因此,低氧胁迫和盐胁迫是植物源食品富集GABA 的有效方法[6]。Li等[7]研究了溶氧量对发芽蚕豆富集GABA的影响,但关于低氧胁迫联合盐胁迫对蚕豆富集GABA的研究未见报道。本研究考察了蚕豆发芽过程中,低氧联合盐胁迫处理条件下,MSG、CaCl2和VB6对GAD活性和对GABA富集效果的影响,旨在为蚕豆发芽富集GABA提供科学依据。
1 材料与方法
1.1 材料与仪器
1.1.1 试验材料
供试蚕豆由江苏沿江地区农业科学研究所提供,品种为启豆2号。
γ-氨基丁酸(GABA)标准品:购自Sigma公司,其他试剂均为国产分析纯。
1.1.2 试验仪器
Orion818型pH测试仪:Orion公司;Agilent 1200 Series高效液相色谱仪:Agilent公司;TDL-40B离心机:上海安亭科学仪器厂;HH-6型数显恒温水浴锅:常州国华电器有限公司;PYX-DHS-BS型隔水电热恒温培养箱:上海跃进医疗器械厂;Anke GL-20G-Ⅱ冷冻离心机:上海安亭仪器厂;JA2003型电子天平:上海精密科学仪器有限公司。
1.2 试验设计
1.2.1 单因素试验
称取50 g蚕豆,水清洗后,用1%的次氯酸钠水溶液消毒30 min,然后用蒸馏水洗去残留的次氯酸钠。消毒后的蚕豆于蒸馏水中30℃浸泡8 h,取出吸干表面水分,置于托盘中(33±1)℃避光培养1.5 d。取出后置于具塞培养瓶(φ6 cm×9 cm)中,加入60 mmol/L的NaCl溶液(用10 mmol/L柠檬酸-柠檬酸钠缓冲液配制,pH 3.5)200 mL,于(33±1)℃下避光培养4 d。期间,向培养液中连续通入空气,通气量为1.2 L/min,形成低氧联合盐胁迫条件。
培养液中谷氨酸钠(MSG)为0~2.5 mg/mL、CaCl2为0~15 mmol/L、VB6为0~150 μmol/L。
1.2.2 响应面试验
根据单因素试验结果,以MSG浓度(A)、CaCl2浓度(B)和VB6浓度(C)3个因素与GABA含量进行响应面试验设计(表1),优化培养液组分。通过Design Expert软件对试验数据进行回归分析,预测发芽蚕豆富集GABA的最优培养液组成。以pH 3.5柠檬酸-柠檬酸钠缓冲液培养为对照。

表1 响应面设计因素与水平
1.3 测定指标与方法
GABA含量:参照Bai[8]的方法测定;GAD活性:参照 Li[7]的方法测定。
2 结果与讨论
2.1 MSG对发芽蚕豆GABA含量和GAD活性的影响
由表2可见,当MSG质量浓度低于1.0 mg/mL时,发芽蚕豆GABA含量随着MSG质量浓度的增加而增加,MSG大于1.0 mg/mL时,GABA含量随MSG质量浓度升高而降低。MSG质量浓度在1.0~2.0 mg/mL时,GABA含量差异不显著。因此,本研究选择MSG质量浓度为1.0 mg/mL。GAD活性变化趋势与GABA含量变化相似。当MSG质量浓度为1.0 mg/mL时,GAD活性最高,为对照的1.43倍。谷氨酸和谷氨酸钠(MSG)在GAD作用下转化为GABA,其含量增多可使GABA支路中碳流量增加,并调节GAD 活性[9]。王玉萍等[10]研究表明,添加 MSG 对发芽糙米GABA的富积影响不显著。本研究与之报道不一致,原因尚有待探讨。
2.2 CaCl2对发芽蚕豆GABA含量和GAD活性的影响
植物GAD氨基酸序列中C末端存在CaM结合区,该区域与Ca2+/CaM结合后可提高GAD活性[3]。低氧条件下,添加Ca2+后,发芽粟谷中GAD活性得以提高,因而GABA含量提高[8]。由表3可知,随着CaCl2浓度的增加,发芽蚕豆GAD活性和GABA含量均呈先升高后降低的变化趋势。当CaCl2浓度为6 mmol/L时,GAD活性和GABA含量均达到最大值,分别为对照的1.88倍和1.09倍。这一研究结果也证实了培养液中添加适当浓度的CaCl2可提高蚕豆GAD活性和GABA含量。
2.3 VB6对发芽蚕豆 GABA含量合 GAD活性的影响
发芽蚕豆GABA含量随VB6浓度增加呈先升后降的趋势(表4)。

表2 MSG对发芽蚕豆GABA含量和GAD活性的影响

表3 CaCl2对发芽蚕豆GABA含量和GAD活性的影响

表4 VB6对发芽蚕豆GABA含量和GAD活性的影响
VB6浓度为60~120μmol/L时,GAD活性无显著差异,而VB6浓度为60μmol/L时,GABA含量达到最大值,是对照的1.35倍。VB6是磷酸吡哆醛(PLP)的前体物质,而PLP是GAD的辅基,适量添加PLP可促进其与脱辅基蛋白结合而激活GAD,促进MSG脱羧生产GABA,从而增加GABA含量[11]。张磊等[11]研究发现,在米糠孵育液中添加 VB6,可使GABA含量得到提高,本研究结果与此一致。
2.4 低氧联合盐胁迫下发芽蚕豆GABA富集的培养液组分优化
2.4.1 模型分析
对MSG浓度(A)、CaCl2浓度(B)和 VB6浓度(C)进行了三因素三水平响应面分析试验,试验设计与结果见表5。利用Design Expert软件对表5数据进行二次多元回归拟合,得到GABA含量对编码自变量A、B和C的二次多项回归方程:
y=1.181 15+0.525 00A+0.011 592C+1.875 000 AC-0.000 450BC-0.291 41A2-0.028 222B2-0.000 076 6C2
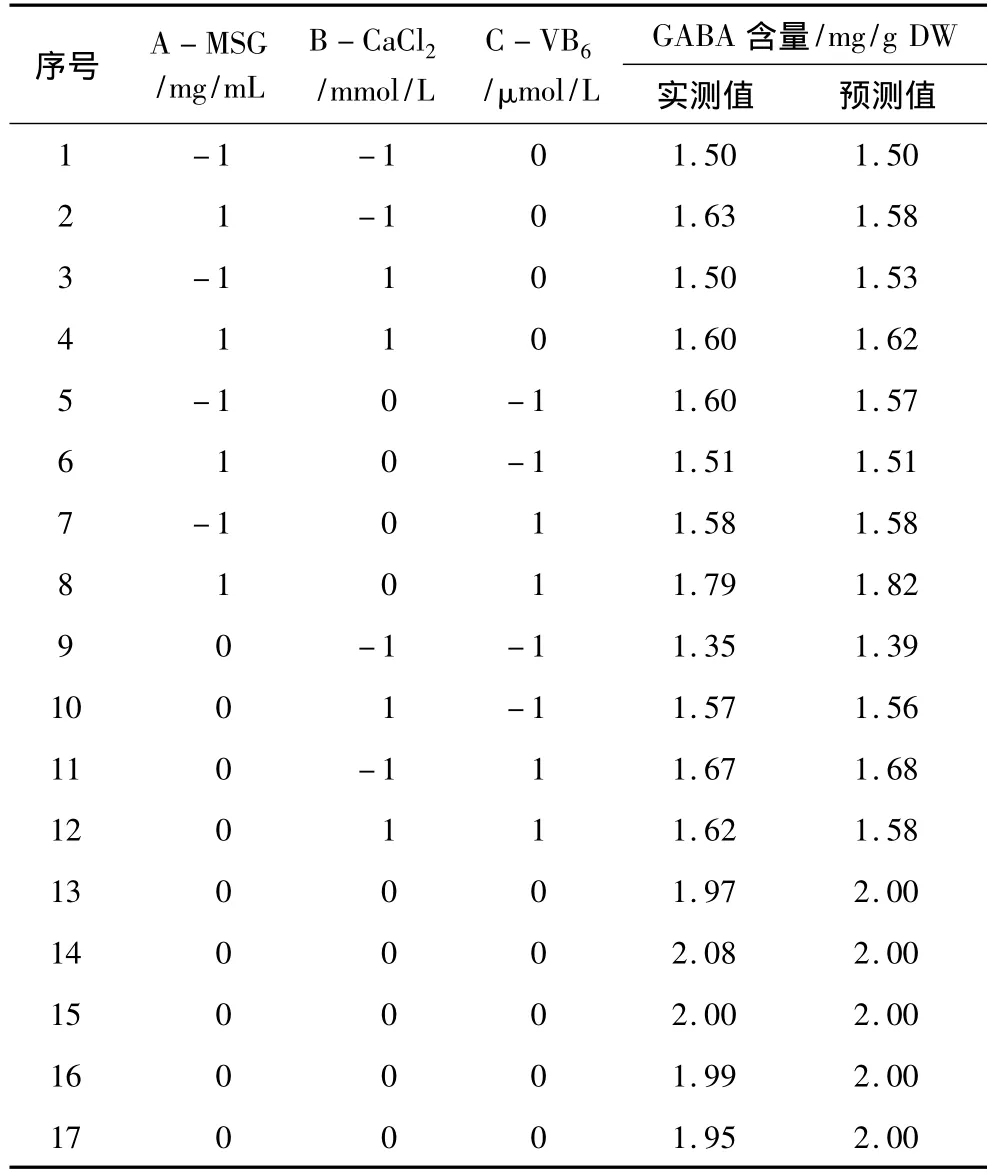
表5 中心组合试验设计和结果
对上述回归模型进行方差分析(表6)。结果表明,模型是显著的(P<0.001),回归模型的决定系数为0.976,说明该模型能够解释97.6%的变异。因此,可用此模型对GABA含量进行分析和预测。

表6 回归模型方差分析
2.4.2 发芽蚕豆富集GABA培养液组分优化
利用Design Expert软件对表5数据进行二次多元回归拟合,所得到的二次回归方程响应曲面对应的等高线见图1~图2。
图1显示,CaCl2浓度为6 mmol/L时,MSG和VB6对发芽蚕豆GABA含量的交互作用显著。因为等高线的形状反映交互效应的强弱大小,椭圆形表示两因素交互作用显著[12]。VB6浓度一定时,GABA含量随MSG浓度增加先升高后降低。VB6浓度较低(10~35μmol/L)时,图1中等高线分布较密,表明此时MSG浓度对GABA含量的影响较大。当MSG浓度为1.1 mg/mL,VB6浓度为72 μmol/L 时,GABA含量达到最高值。MSG是GAD作用的底物,添加适量的MSG可以提高GAD活性,增加GABA含量。

图1 MSG和VB6的交互作用对GABA含量的影响
CaCl2和VB6对发芽蚕豆GABA含量的交互作用显著(图2)。与VB6相比,CaCl2浓度对GABA含量的影响较大。当CaCl2浓度一定时,GABA含量随VB6浓度增加呈逐渐升高的趋势。CaCl2为 6.1 mmol/L,VB6浓度为72μmol/L时,GABA含量达到峰值。由此可见,培养液添加适当浓度的CaCl2和VB6可提高发芽蚕豆中GABA含量。

图2 CaCl2和VB6的交互作用对GABA含量的影响
Glu是GABA的前体物之一,而MSG是Glu的盐存在形式,其在水溶液中解离后,渗入蚕豆种子,从而被发芽过程中所激活的GAD降解产生GABA;植物GAD氨基酸序列中C末端存在CaM结合区,该区域与Ca2+/CaM结合后可提高GAD活性,在足够多的底物存在的情况下,从而提高GABA产率;PLP是GAD的辅酶,VB6与PLP结构相似,可起到辅酶功能,在蚕豆发芽过程中参与GAD的合成,提高GAD活力,促进Glu脱羧形成GABA,从而使GABA含量增加。
目前报道,GABA的形成有两条途径,即GABA支路和多胺氧化降解途径[13]。其中GABA支路对发芽蚕豆GABA富集的贡献约占70%(数据未展示)。在GABA支路中,GAD活力的提高和足够的底物(MSG)对GABA的产生必不可少,而VB6是GAD辅酶的结构类似物,参与酶的合成来提高GAD活力。所以,MSG和VB6的加入对GABA的形成具有协同作用。作为第二信使,Ca2+通过信号传递的方式与VB6共同调节GAD活力,对GABA的产生具有协同作用。
2.4.3 验证性试验
对发芽蚕豆GABA含量的二次多项模型解逆矩阵后得出,在低氧联合盐胁迫培养条件下,培养液组分中MSG 质量浓度为 1.1 mmol/L、CaCl2浓度6.1 mmol/L、VB6浓度72μmol/L时,发芽蚕豆中GABA富集量的预测值为2.01 mg/g DW,实测值为(1.98±0.09)mg/g DW,是对照[(1.08 ±0.01)mg/g DW]的1.83倍,且高于随机选择组。相关性分析表明,实测值与预测值接近,表明实验所拟合的模型可用来预测培养液组分和试验响应值之间的关系(表7)。

表7 验证试验设计和结果
3 结论
低氧联合盐胁迫下,培养液中3种组分(MSG、CaCl2和VB6)对发芽蚕豆GAD活性和GABA含量的影响均达到显著水平(P<0.005)。当MSG质量浓度为 1.1 mg/mL、CaCl2浓度为 6.1 mmol/L、VB6浓度为72μmol/L时,发芽蚕豆中GABA富集量是对照[(1.08 ±0.01)mg/g DW]的1.83 倍,达到(1.98 ±0.09)mg/g DW。
[1]Kmiecik W,Lisiewska Z,Jaworska G.Content of ash components in the fresh and preserved broad bean(Vicia faba v major)[J].Journal of Food Composition and Analysis,2000,13:905-914
[2]张晖,徐满英.γ-氨基丁酸作用的研究进展[J].哈尔滨医科大学学报,2006,40(3):267-268
[3]Shelp B J,Bown A W,Mclean M D.Metabolism and functions of gamma- aminobutyric acid[J].Trends in Plant Science,1999,411:446-452
[4]Fukao T,Bailey- Serres J.Plant responses to hypoxia- is survival a balancing act[J].Trends in Plant Science,2004,9:449-456
[5]Reddy V S,Reddy A S N.Proteomics of calcium- signaling components in plants[J].Phytochemistry,2004,65:1745-1776
[6]Komatsuzaki N,Tsukahara K,Toyoshima H,et al.Effect of soaking and gaseous treatment on GABA content in germinated brown rice[J].Journal of Food Engineering,2007,78(2):556-560
[7]Li Y,Bai Q Y,Jin X J,et al.Effects of cultivar and culture conditions onγ-aminobutyric acid accumulation in germinated fava beans(Vicia faba L.)[J].Journal of the Science of Food and Agriculture,2010,90:52-57
[8]Bai Q Y,Fan G J,Gu Z X,et al.Effects of culture conditions onγ-aminobutyric acid accumulation during germination of foxtail millet(Setaria italica L.)[J].European Food Research and Tech-nology,2008,228:169-175
[9]顾振新,蒋振辉.食品原料中γ-氨基丁酸(GABA)形成机理及富集技术[J].食品与发酵工业,2002,28(10):65-69
[10]王玉萍,韩永斌,顾振新,等.谷氨酸钠和抗坏血酸对发芽糙米中GABA富积效果的影响[J].南京农业大学学报,2006,29(2):94-97
[11]张磊,白青云,曹晓虹,等.米糠富集γ-氨基丁酸的培养液组分优化[J].食品与发酵工业,2008,34(7):77-81
[12]范龚健,韩永斌,顾振新.用响应面法优化红甘蓝色素提取工艺参数[J].南京农业大学学报,2006,29(1):103 -107
[13]李岩.发芽蚕豆γ-氨基丁酸富集途径及高效富集技术研究[D].南京:南京农业大学,2009.
Optimization of Culture Solution Compositions for γ-aminobutyric Acid Accumulation in Germinated Fava Beans(Vicia faba L.)
Chen Hui Yang Runqiang Han Yongbin Gu Zhenxin
(Key Laboratory of Agricultural and Animal Products Processing and Quality Control,Ministry of Agriculture,Nanjing Agricultural University,Nanjing 210095)
In this paper,the effects of MSG,CaCl2and VB6on glutamate decarboxylase(GAD)andγ-aminobutyric acid(GABA)of germinated fava beans were investigated.The culture solution compositions used for GABA accumulation in fava beans were optimized with Box-Behnken design.The results showed that under the condition of hypoxia combined with salt stress,the effects of MSG,CaCl2and VB6on GAD and GABA of germinated fava beans all reached significant levels(P <0.005).The quadric regression equation was established between GABA content and culture solution compositions.The R-squared reached 0.976,which meant that the equation could predict the changes of accumulation contents of GABA well.The optimum compositions of culture solution for GABA accumulation were MSG of 1.1 mg/mL,CaCl2of 6.1 mmol/L and VB6of 72 μmol/L.Under these conditions,the content of GABA was(1.98 ±0.09)mg/g DW,which was 1.83 times as much as that of the control[(1.08 ±0.01)mg/g DW].
germinated fava beans,γ - a minobutyric acid,accumulation,optimization
A
1003-0174(2011)11-0027-05
中央高校基本科研业务费专项资金项目(KYZ200 917)
2011-01-26
陈惠,女,1986年出生,硕士,食品中功能成分的富集技术
顾振新,男,1956年出生,教授,博士生导师,生物技术与功能食品
猜你喜欢
天津医科大学学报(2021年3期)2021-07-21 09:03:42
现代畜牧科技(2021年6期)2021-07-16 05:49:54
小读者(2021年2期)2021-03-29 05:03:30
公民与法治(2020年13期)2020-07-27 01:30:36
中国粮油学报(2019年4期)2019-07-12 09:06:28
学生天地(2018年9期)2018-09-06 13:11:18
现代园艺(2017年11期)2017-06-28 11:32:37
国外医药(抗生素分册)(2016年5期)2016-07-12 14:25:34
磁共振成像(2015年5期)2015-12-23 08:52:54
实验动物与比较医学(2014年5期)2014-02-28 14:53:12
