
自主复制序列结合因子 1结合位点拷贝数对基因沉默的影响
2011-11-19 11:21:38张新民王浩天颜炜群吉林大学药学院吉林长春300
中国老年学杂志 2011年4期
张新民 于 群 毕 鑫 刘 楠 王浩天 成 岩 颜炜群 (吉林大学药学院,吉林 长春 300)
抗衰老的主要目的是控制过速衰老,预防与年龄相关的疾病的发生和发展,提高人的生活质量〔1,2〕。目前,对人类生命健康危害最大的疾病是癌症,癌症一般是由细胞内一些基因的异常表达尤其是一些本该沉默的基因表达引起的〔3~7〕,对于基因表达沉默的机制目前还没研究清楚。真核生物DNA与组蛋白组成核小体进而折叠形成染色质,整个染色质分为结构松散的基因转录表达的常染色质和高度浓缩的基因不表达异染色质。酿酒酵母作为一种模式生物解决了许多遗传学、分子生物学等方面的难题。在酿酒酵母中,第三号染色质的 (HML以及HMR)区域形成了与高等生物异染色质相似的特性结构的染色质〔8,9〕。酿酒酵母的沉默子 HML-E和 HML-I对其第三号染色质的 HML区域基因沉默起着决定作用,该沉默子由复制起点识别复合物 (Origin Recognition Comp lex,ORC)结合位点,Rap1p(Rep resso rActivator Protein)结合位点和自主复制序列结合因子 1(Abf1)所组成,这些结合位点称为原沉默子 (p rotosilencer)〔10〕。这些原沉默子独自无法产生基因沉默作用,但是与沉默子相互作用下,能够增强原沉默子或沉默子的基因沉默作用,但增加原沉默子拷贝数是否能增强基因沉默,甚至是产生基因沉默没有相关研究。本研究用右侧带有报告基因乳清苷酸脱羧酶基因(URA3)的不同拷贝数的原沉默子 Abf1p结合位点,以及不带任何结合位点的序列替代酵母基因组 HML区域的 HML-I沉默子后发现,Abf1p结合位点单独存在无论拷贝数多少都无法对右侧沉默基因,而在沉默子存在时,随着 Abf1p拷贝数的增加基因沉默的能力逐渐增强。
1 材料与方法
1.1 材料 酿酒酵母菌种 YXB6〔MATa、HMRa、HMLa、ED79-113TSUP4-o、IΔ242、LEU2-GAL10-FLP1、ura3-52、ade2-1、lys1-1、his5-1、can1-100(ciro)〕,大肠杆菌 (DH5α)由本实验室保存。DNA taq酶、蛋白胨、酵母提取物购自 Invitrogen公司,DNA T4连接酶,BstB I,Kpn I,HindⅢ,SnaB I等限制性内切酶购自 NEB公司。
1.2 质粒的构建 将酿酒酵母第三号染色质 AatⅡ和 BamH I之间 HML基因序列并插入到 pUC12 AatⅡ和 BamH I之间制成质粒 pQY253,接着将 pQY253上的 HML-I沉默子替换成 HMRE沉默子构建出 pQY254。接着在 pQY254的 EcoRV位点插入URA3制作出质粒 pXZ8。使用点突变 PCR法将 pXZ8的 HMRE的三个结合位点 ORC、Rap1、Abf1依次突变成 Kpn I、Spe I、Mfe I;kpn I、Spe I、Abf1p1p结合位点 ;Kpn I、Abf1p1p 结合位点、Abf1p1p结合位点;Abf1p1p结合位点、Abf1p1p结合位点、Abf1p1p结合位点的质粒,依次命名为 pXZ10,pXZ11,pXZ12以及pXZ13。
1.3 酵母菌种的构建以及 Southern印迹法鉴定 使用BspH I/NgoMⅣ两个限制性内切酶酶切 pXZ10,pXZ11,pXZ12,pXZ13这四个质粒,LiAc法转化到酿酒酵母 YXB6中,在缺失尿嘧啶合成缺陷型培养板 (-ura)上进行筛选。挑取单克隆菌落,-ura培养液培养,玻璃珠法提取基因组 DNA,使用 EcoRV以及BamHⅠ酶切 DNA,以 BglⅡ之间的 URA3基因 DNA为探针Southern印迹鉴定。最后获得含有 0,1,2,3个 Abf1p结合位点的酵母菌种 YXZ75,YXZ79,YXZ80,YXZ81。然后再使用两端带有 HML-E两侧的同源序列的 KanMX基因分别替换掉上述菌种的 HML-E,采用 Southern印迹法鉴定菌种,最终依次得到不带有 HML-E沉默子的酵母菌种 YXZ75DL,YXZ79DL,YXZ80DL,YXZ81DL。菌种构建示意图见图 1。
1.4 利用URA3基因检测不同拷贝数 Abf1p结合位点的基因沉默表型 阳性克隆酵母菌 (YXZ75,YXZ79,YXZ80,YXZ81)在全合成培养基 (SC)中培养 24 h,在 96孔板对菌液进行 10倍比稀释,分别在 SC,-ura,SC+5-氟乳清酸 (FOA)上点样,在 30℃培养 24 h,48 h,72 h,96 h时,使用扫描仪扫描菌生长状态。

图 1 菌种构建示意图
2 结 果
2.1 酵母菌种的构建以及 Southern印迹法鉴定 pXZ10,pXZ11,pXZ12以及 pXZ13经 BspH I以及 NgoMⅣ酶切后转化酵母菌种 YXB6,如果同源重组发生在 HML-I左侧,提取基因组DNA经 EcoRⅤ酶切,Southern印迹鉴定在图谱上将看到2 384 bp,1 150 bp的两条特异性的条带;如果同源重组发生在HML-I右侧,HML-I序列中有一个 EcoRV位点,会看到2 384 bp,982 bp两条特异性条带,酶切不彻底时还能在1 150 bp附近看到条带。限制性内切酶 EcoRV消化提取基因组DNA的 EtBr胶图谱,可以看出 DNA条带弥散,说明消化的比较完全。Southern印迹鉴定的结果,可以看到阴性对照及所有的菌株都有两条非特异性的条带,这是由于该菌种为-ura3-52的突变型菌株,基因组上仍存在一份突变无功能的-ura3基因,它与野生型-ura3基因相同的序列,在阴性对照组 YXB6和转化菌种都会存在一些非特异的条带;1、2号菌株的 2 384 bp、1 150和982 bp处有三条特异性条带,这说明同源重组发生在 HML-I右侧 ,未能替代 HML-I。4、6、7、8、10、12、13、15、16 菌株在2 384 bp,1 150 bp的有两条特异性条带,说明他们是正确的阳性克隆。由于 YXZ79,YXZ80,YXZ81结果与 YXZ75相差不多,所以本文中没有一一列出。见图 2A~2C。
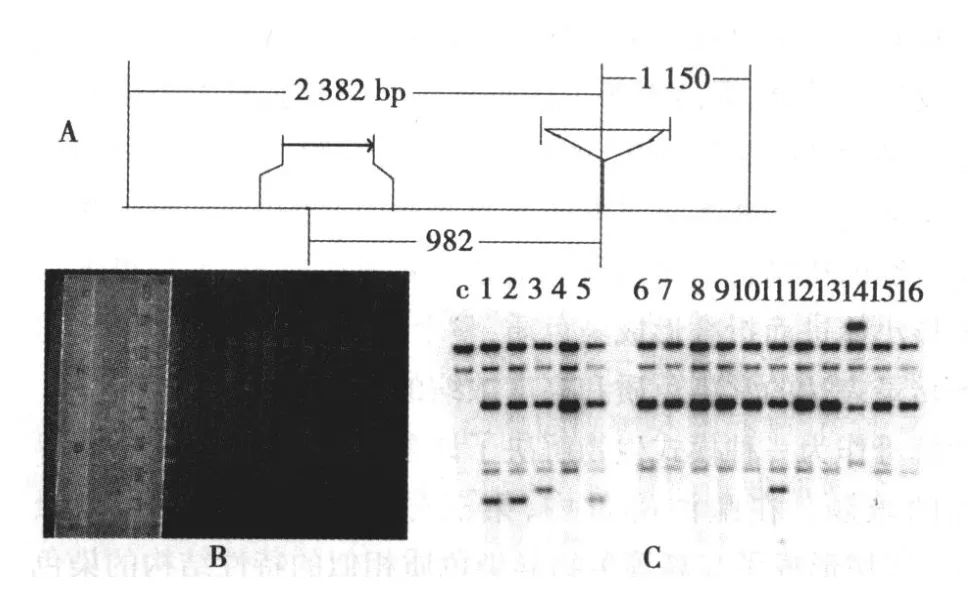
图 2 YXZ75菌株的 Southern印迹鉴定结果
2.2 基因沉默表型的研究 URA3基因是一种在基因沉默研究中广泛使用的报告基因,在生存环境中缺失尿嘧啶 (-ura)时,URA3可以被 pp rl1激活从而大量表达,因此-ura培养板菌体生长状态反映了URA3基因的激活转录状态,在 SC平板上尿嘧啶存在,URA3基因仅仅有少量正常表达,URA3蛋白作用于嘧啶代谢途径,使 FOA变成一种有毒的代谢产物,引起酵母细胞死亡,因此 SC+FOA平板上反映了基因沉默的情况。无论是HML-E存在与否,SC平板上酵母的生长状态没有任何区别,这说明替换 HML-E以及 HML-I沉默子对酵母的正常生命活动没有影响。在 HML-E沉默子存在的条件下,在-ura培养板上生长状态基本没有区别,说明原沉默子 Abf1p结合位点的拷贝数对激活水平的表达影响不大;在 SC+FOA平板上,随着拷贝数的增加,FOA平板上酵母的生长逐渐变多,说明基因沉默逐渐增强。在 KanMX替换了 HML-E情况下,无论是在-ura还是 SC+FOA平板上,酵母的生长状态都没有任何区别,说明 Abf1p独自无法对右侧的基因产生沉默作用,而且增加拷贝数也不能。见图 3。
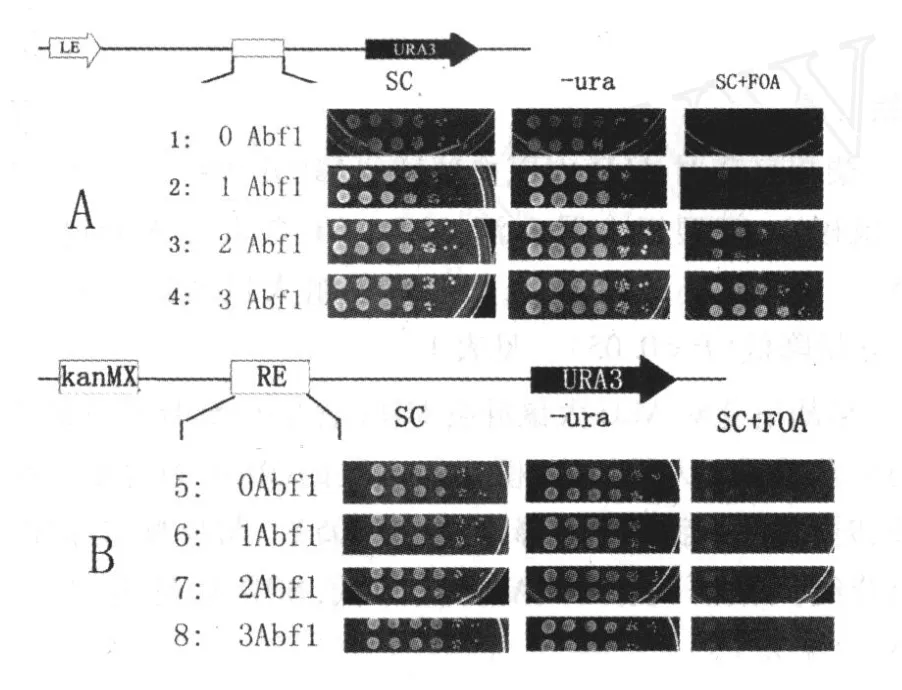
图 3 Abf1p结合位点基因沉默表型示意图
3 讨 论
衰老以及癌症的发生与遗传信息的突变、基因组的重排、表观遗传学的改变如 DNA的甲基化、组蛋白的乙酰化等这些染色质异常有关,这些情况除了遗传信息突变会导致蛋白质发生变化外,基本都是基因表达量发生了变化,对于这种变化的本质目前还没有研究清楚〔11,12〕。酿酒酵母是一种生物研究的模式生物,酵母基因组与高等生物的染色质相似分为常染色质和异染色质区域,基因不表达的异染色质 HML区域形成与沉默子密切相关,沉默子由原沉默子组成,单独的原沉默子可以增强沉默子基因沉默的能力,延长沉默子基因沉默的范围,但是尚未有人研究原沉默子拷贝数对基因沉默的影响。Abf1是一种DNA位点特异性结合蛋白,它结合到 5-TnnCGTnnnnnnTGAT-3特定的保守序列〔13〕。在酿酒酵母基因组的沉默配型座位,复制子,端粒 X-区域,许多基因启动子区域等许多位置都有Abf1p结合位点的分布。Abf1p结合到这些区域的位点上直接参与许多与染色质相关的事件,例如 DNA复制,基因沉默,染色质重构,核苷酸剪切,基因的激活与抑制等〔14,15〕。本研究研究了Abf1p在基因沉默中的作用,Abf1p结合位点单独存在时,无论拷贝数是多少,都无法对外源基因进行沉默,HML-E沉默子的存在时,Abf1p结合位点能对外源基因起沉默作用,并且随着拷贝数的增加沉默作用增强。对于其他原沉默子如 Rap1p,ORC结合位点增加拷贝数是否能够增强基因沉默,以及 Abf1p结合位点增强基因沉默的内在机制还需要进一步的研究。
1 胡 征,刘丹丹 .冬虫夏草钙制剂对衰老小鼠的抗衰老和免疫调节作用〔J〕.中国老年学杂志,2010;30(19):1410-3.
2 袁 丹,陈海平 .抗衰老 Klotho基因的功能及与人类疾病的关系〔J〕.中国老年学杂志,2010;30(14):2074-76.
3 王自强,周燕虹,蔡志民,等 .抑癌基因 ING1在大肠癌中的表达与突变 (英文)〔J〕.中国临床康复,2005;9(22):262-4.
4 刘维强,孙筱放,银益飞,等 .正常与异常核型人类胚胎干细胞体外分化不同时期基因表达的异同〔J〕.现代生物医学进展,2009;9(18):3422-6.
5 Yusa A,Miyazaki K,Kimura N,et a l.Ep igenetic silencing of the sulfate transporter gene DTDST induces sialyl Lewisx exp ression and accelerates p roliferation of colon cancer cells〔J〕.Cancer Res,2010;70(10):4064-73.
6 Malek A,Tchern itsa O.Evaluation of targets forovarian cancer gene silencing therapy:in vitro and in vivo app roaches〔J〕.Method s Mol Biol,2010;62(3):423-36.
7 Ruckert F,Samm N,LehnerAK,et a l.Simultaneous gene silencing ofBcl-2,X IAP and Survivin re-sensitizes pancreatic cancer cells towards apop tosis〔J〕.BMC Cancer,2010;10:379.
8 Yu Q,Kuzm iak H,Olsen L,et a l.Saccharomyces cerevisiae Esc2p interactswith Sir2p through a small ubiquitin-like modifier(SUMO)-binding motif and regulates transcrip tionally silent chromatin in a locus-dependent manner〔J〕.J Biol Chem,2010;285(10):7525-36.
9 Xu EY,Bi X,Holland MJ,et a l.Mutations in the nucleosome core enhance transcrip tional silencing〔J〕.MolCellBiol,2005;25(5):1846-59.
10 Lebrun E,Revardel E,Boscheron C,et a l.Protosilencers in Saccharomyces cerevisiae subtelomeric regions〔J〕.Genetics,2001;158(1):167-76.
11 Sp icakova T,O′Brien MM,Duran GE,et a l.Exp ression and silencing of the microtubule-associated p rotein Tau in breast cancer cells〔J〕.Mol Cancer Ther,2010;9(11):2970-81.
12 Nicolas FE,Lopez-Gomollon S,Lopez-Martinez AF,et a l.Silencing human cancer:identification and uses ofm icroRNAs〔J〕.Recent Pat AnticancerDrug Discov,2011;6(1):94-105.
13 Beinoraviciute-Kellner R,Lipp s G,Krauss G.In vitro selection of DNA binding sites forABF1 p rotein from Saccharomyces cerevisiae〔J〕.FEBS Lett,2005;579(20):4535-40.
14 Yu S,Sm irnova JB,Friedberg EC,et a l.ABF1-binding sitesp romote efficient global genome nucleotide excision repair〔J〕.J Biol Chem,2009;284(2):966-73.
15 GanapathiM,Palumbo MJ,Ansari SA,et a l.Extensive role of the general regulatory factors,Abf1 and Rap1,in deter mining genome-wide chromatin structure in budding yeast〔J〕.Nucleic Acids Res,2010:16.
猜你喜欢
陶瓷研究(2022年3期)2022-08-19 07:15:18
畜牧兽医学报(2022年3期)2022-03-30 02:29:20
中国畜牧兽医(2022年1期)2022-02-15 10:46:40
云南画报(2021年10期)2021-11-24 01:06:56
河北医学(2021年10期)2021-10-27 00:37:14
现代泌尿外科杂志(2019年10期)2019-10-31 07:31:54
中国临床医学影像杂志(2019年6期)2019-08-27 02:59:50
生物学通报(2019年2期)2019-06-15 01:33:42
小学生优秀作文(高年级)(2018年4期)2018-09-11 01:23:22
发明与创新(2015年25期)2015-02-27 10:39:16
