
毛红椿光合速率及生理生态因子的日变化规律
2011-11-16 09:09吴际友王旭军彭辉明杨广文
湖南林业科技 2011年2期
吴际友, 程 瑞, 王旭军, 程 勇, 彭辉明, 杨广文
(1.湖南省林业科学院, 湖南 长沙 410004; 2.岳阳市林业科学研究所, 湖南 岳阳 414000)
毛红椿光合速率及生理生态因子的日变化规律
吴际友1, 程 瑞1, 王旭军1, 程 勇1, 彭辉明2, 杨广文2
(1.湖南省林业科学院, 湖南 长沙 410004; 2.岳阳市林业科学研究所, 湖南 岳阳 414000)
2010年8月利用Li-cor 6400便携式光合分析仪对2年生毛红椿幼树光合速率及生理生态因子进行测定,对其日变化规律进行了研究。研究结果表明:毛红椿光合速率日变化呈双峰曲线,呈现出光合“午休”现象;净光合速率最高峰出现在上午8∶00,次高峰出现在中午13∶00;相关分析和回归分析结果是,毛红椿光合速率的主要影响因子为光合有效辐射、气孔导度和大气中CO2浓度等。
毛红椿; 光合速率; 生理生态因子; 日变化规律
毛红椿(Toonaciliatavar.pubescens)是楝科(Meliaceae ) 香椿属(Toona),落叶大乔木植物,为阳性喜光树种,喜温暖湿润气候,耐寒性好,能耐-15℃的低温,对土壤要求不高,在水湿地和干旱贫瘠地均能正常生长,主要产于四川、贵州、福建和安徽,多呈天然零星分布,垂直分布于海拔500~2500m[1]。毛红椿木材色泽艳红,纹理美观,素有“中国桃花心木”之称,已成为重要的工业阔叶用材树种[2]。光合作用是植物最重要的生理过程,是评价植物第一生产力的标准之一。已有报导对毛红椿的研究主要注重于育种和栽培[3-5]等方面,关于其光合生理特性等方面的研究[6]甚少。我们测定了2年生毛红椿幼树的光合日变化规律,为毛红椿栽培和良种选育提供基础数据。
1 试验地概况
试验地设在湖南省林业科学院试验林场苗圃,该处地理坐标为东经112°59′、北纬28°05′,年平均气温16.8℃,平均日照时数1496~1850h,年降水量1400~1900mm,无霜期264d,属中亚热带季风湿润气候区,光照充足,雨量丰沛;海拔为110m,土壤为砂岩发育的红壤,土层厚达60cm以上;土壤肥力中等,pH值为6.2。
2 材料与方法
2.1试验材料
以田间栽培的毛红椿2年生幼树为试验材料。
2.2试验方法
于2010年3月将毛红椿2年生幼树移入大棚,株行距1m×1m,常规管理。2010年8月选择晴朗无云的天气,选取3株生长环境条件比较一致的健康植株作为供试材料,挂牌标记树冠中上部南面受光条件好的枝条从上至下第4、5片健康的成熟叶片(每株3~5片),利用Li-cor 6400便携式光合分析仪(美国产)进行活体测定,每片5次取值。试验从上午8∶00开始,下午18∶00结束,每隔2h测定1次,并在中午13∶00加测1次。测定参数有:光合速率(Pn)、蒸腾速率(Tr)、光合有效辐射(PAR)、气孔导度(Cond)、叶面温度(Tl)、气温(Ta)、大气相对湿度(RH)、胞间CO2浓度(Ci)、大气中CO2浓度(Ca)等。
2.3计算及分析方法
应用Excel2003、SPSS13.0统计分析软件对数据进行作图、相关分析与回归分析。
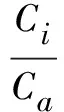
3 结果与分析
3.1外界环境因子的日变化规律
3.1.1 光合有效辐射(PAR)的日变化规律 由图1可以看出,光合有效辐射(PAR)日变化为“单峰”曲线,随着太阳入射角的增大,光合有效辐射逐渐增大,下午13∶00达到全天最大值,约为1579.711μmol·m-2·s-1,此后逐渐下降,并于下午18∶00的时候达到全天最低值。
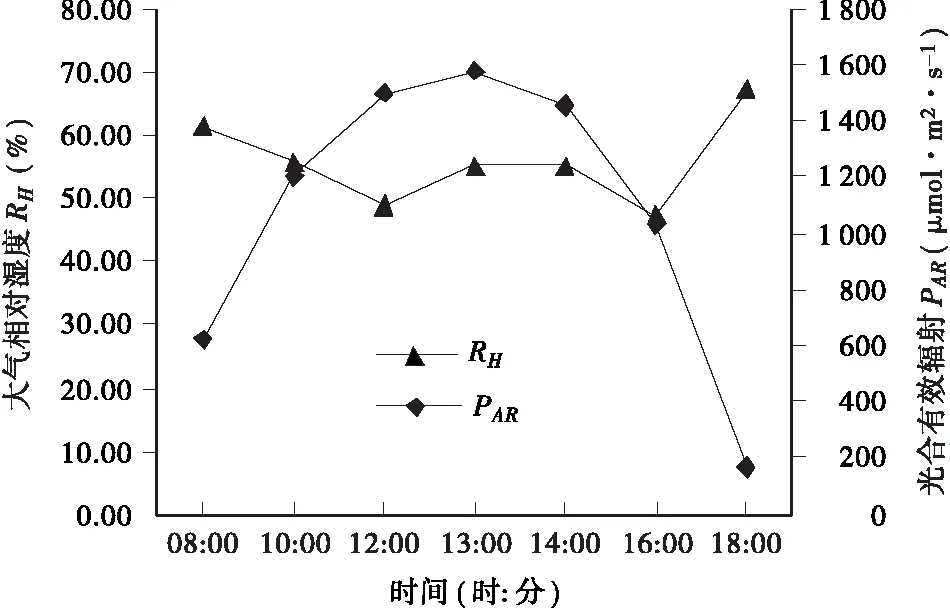
图1 大气相对湿度及光合有效辐射日变化 Fig.1 Diurnal variation of atomsphere relative humidity and photosynthetically active radiation
3.1.2 大气相对湿度(RH)的日变化规律 大气相对湿度(RH)日变化大体表现为“W”形状变化,从上午8∶00开始呈下降趋势(图1),于中午12∶00左右达一低谷值;但到下午13∶00 时,RH出现迅速回升,并在一定时间内保持相对稳定,而且当RH达到全天最小值时,Vpdl达到全天最大值(图7)。叶面水汽压亏缺增大会加速蒸腾作用,降低叶片水势,影响叶片气孔的开度,从而影响叶片的光合速率。RH与Vpdl的变化曲线呈完全相反趋势,而与Cond的变化曲线(图5)在10∶00到16∶00大致相同,说明RH对气孔导度的变化产生影响,从而影响到Pn。
3.1.3 叶面温度(Tl)和气温(Ta)的日变化规律 叶面温度(Tl)和气温(Ta)的变化趋势相同。由图2可以看出当出现光合“午休”时,全天的最高Ta出现,为42.71℃,随后温度降低,但相对比较稳定,Ta和Tl到下午18∶00左右达到最低值,但仍然高达34.96℃和34.99℃。
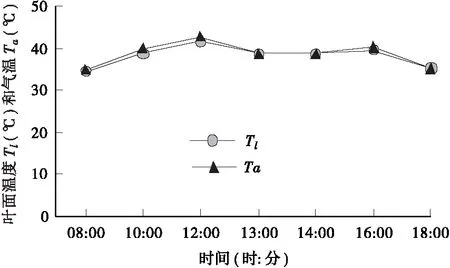
图2 叶面温度及气温日变化曲线Fig.2 Diurnal variation of leaf temperature and atmospheric temperature
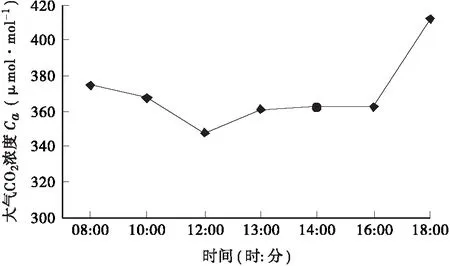
图3 大气中CO2浓度日变化曲线Fig.3 Diurnal variation of atmospheric CO2 concentration
3.1.4 大气中CO2浓度(Ca)的日变化规律 大气中CO2浓度(Ca)经过夜间的累积,在早上8∶00达到一个高峰,随着植物光合速率的进行,在中午12∶00之前逐渐降低,在下午13∶00出现小幅度的回升,并比较平稳的维持到下午16∶00,这与Pn的变化曲线(图4)大致相同,随后出现大幅度升高。
3.2毛红椿光合特性
3.2.1 净光合速率(Pn)日变化规律 在生长季节,毛红椿净光合速率(Pn)日变化曲线呈双峰型(图4),在早上8∶00Pn达到全天最高值,约为20.62μmol·m-2·s-1,随后显著降低,在中午12∶00,净光合速率下降为最大时的69%,形成低谷;然后又缓慢上升,在下午13∶00左右形成第二个高峰,但上升幅度不大,且第二峰值明显低于第一峰值,约为14.33μmol·m-2·s-1。由此可见,毛红椿存在 “午休”现象,但不是很明显。此后,毛红椿Pn持续下降,在下午16∶00时急剧下降,至下午18∶00时达到全天最低值,约为1.82μmol·m-2·s-1。
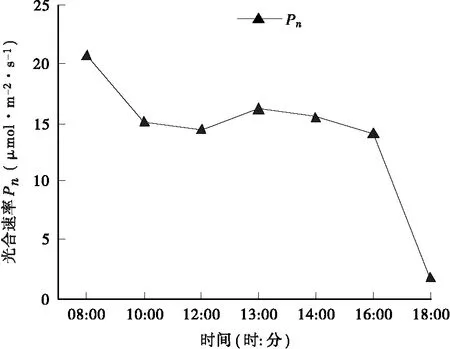
图4 毛红椿净光合速率日变化曲线Fig.4 Diurnal variation of net photosynthetic rate (Pn) of T. ciliata var. pubescens
3.2.2 气孔导度(Cond)和胞间CO2浓度(Ci)的日变化规律 气孔导度(Cond)是指植物气孔传导CO2和水汽的能力,它对光合速率有一定的影响。气孔导度与胞间CO2浓度(Ci)全天的变化规律相似,上午10∶00到下午18∶00的日变化趋势几乎完全一致,在上午10∶00 到中午12∶00逐渐降低,但是在中午12∶00到13∶00开始升高,随后缓慢下降(图5)。由图5还可以看出,Ci在上午10∶00到下午16∶00有个缓慢降低的波动变化,且与Pn的变化基本相同,这与张露等对毛红椿幼苗光合特性研究中Ci与Pn变化恰好相反[6]。Farquhar和Sharkey认为,只有当Ci和光合速率的变化相同,两者都减少,且气孔限制值增大,才可以认为光合速率下降的主要原因是由于气孔导度下降造成的。否则,当光合速率下降,Ci上升,气孔限制值(Ls)减小,便可以认为光合速率的下降主要是由于非气孔因素造成的[7]。据此,可以认为毛红椿光合“午休”为“非气孔限制型”(图6)。Salvucci曾指出:光合作用关键酶 — Rubisco的最适活化温度在25~30℃,其活性的高低直接影响光合速率的大小[8]。因此,气温偏高导致光合酶活性降低可能是毛红椿光合“午休”现象的原因之一。
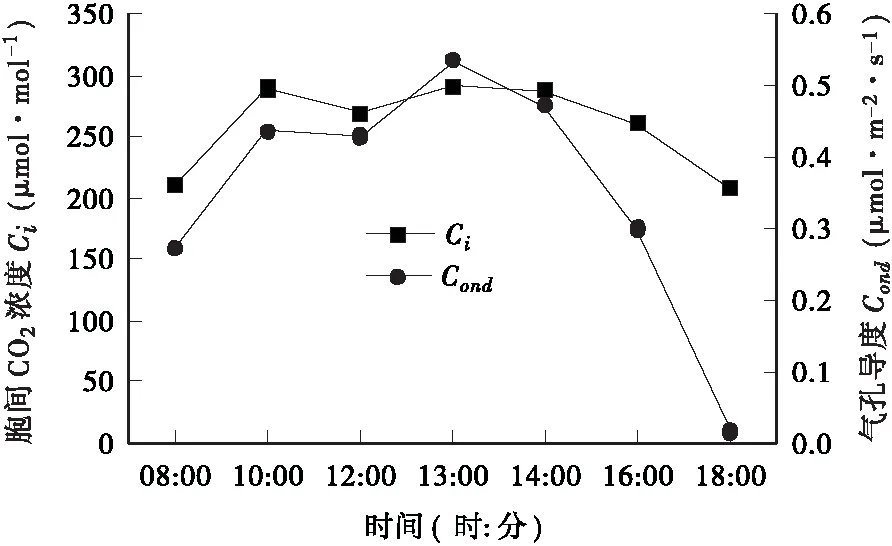
图5 胞间CO2浓度及气孔导度日变化曲线Fig.5 Diurnal variation of intercellular CO2 concen- tration and stomatal conductance
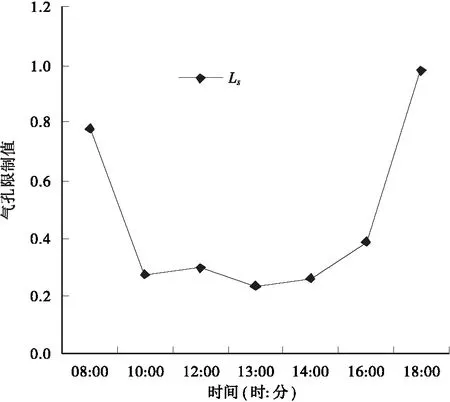
图6 气孔限制值日变化曲线Fig.6 Diurnal variation of stomatal limitation
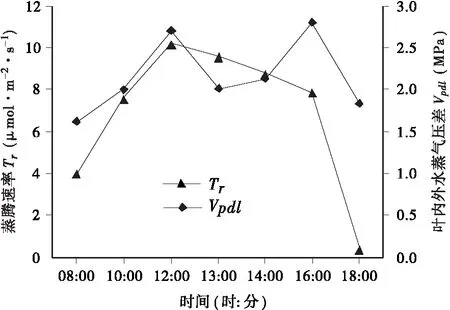
图7 蒸腾速率及叶内外水蒸气压差(Vpdl)的日 变化曲线Fig.7 Diurnal variation of transpiration rate and water vapor pressure between the inside and outside of leaf
3.2.3 蒸腾速率(Tr)及叶面水汽压亏缺(Vpdl)的日变化规律 由图7可知,毛红椿蒸腾速率(Tr)呈单峰曲线,在中午12∶00达到全天最高值,为10.14μmol· m-2·s-1,随后降低,并在下午16∶00后开始急剧下 降,18∶00达到全天最低值,这与Pn的变化相同。叶面水汽压亏缺(Vpdl)的日变化曲线呈明显的双峰型,峰值分别出现在中午12∶00和下午16∶00,Vpdl的变化曲线和Pn的大致相反。
3.3毛红椿净光合速率影响因子的相关与回归分析
植物的光合作用是一个对生态、生理因子敏感的复杂的生理过程,生态因子不仅直接影响光合作用,而且还通过影响植物的生理因子进而影响光合作用。
从表1看出,毛红椿的光合速率与胞间CO2浓度、叶片温度、气孔导度、光合有效辐射、大气温度、大气相对湿度呈正相关关系,其中与叶片温度、气孔导度、光合有效辐射、大气温度、大气相对湿度呈极显著正相关;而与叶面水汽压亏缺和大气CO2浓度呈极显著负相关。

表1 毛红椿净光合速率与各种生理生态因子的相关分析Tab1 Correlationanalysisofthenetphotosyntheticrate(Pn)andeco⁃physiologicalfactorsofT.ciliatavar.pubescensCiCaVpdlTlCondPARTaRHPn0008-0762∗∗-0162∗∗0194∗∗0740∗∗0540∗∗0254∗∗0256∗∗
因影响植物叶片净光合速率的因素很多,因此,净光合速率与生理生态因子的简单相关分析在某些情况下无法真实准确地反映变量之间的关系。而逐步多元回归分析方法能有效地从众多影响因子中筛选出对净光合速率贡献大的因子,并建立净光合速率与这些因子的最优回归方程。为了进一步了解生态因子对毛红椿光合作用的影响,以光合速率为因变量,分别以其他8个生理生态因子为自变量,作多元逐步回归分析。结果表明,Pn=6.583-0.001PAR+24.495Cond,进一步说明影响毛红椿净光合速率的主要因子是光合有效辐射、气孔导度,该结果与相关分析的结果基本吻合。光合有效辐射的改变可以影响空气温度以及叶面水汽压亏缺的变化,叶面水汽压亏缺的改变可以影响气孔的开闭,从而引起气孔导度和胞间CO2浓度的变化,导致树木蒸腾作用的改变,最终影响光合速率的变化。总体来看,生态因子和生理因子交互影响,共同对树木产生作用。
4 结论与讨论
毛红椿在夏天的晴朗天气里,净光合速率日变化曲线呈双峰型,峰值出现在早上8∶00和下午13∶00,在上午10∶00到12∶00由于光合有效辐射过强,气温达到全天最高,表现出“午休”现象。但与张露等人对毛红椿幼苗光合特性研究中全光照时净光合速率存在差异,而曲线趋势则与在遮荫80%、土壤含水量80%~100%条件下的净光合速率变化相似[6]。这是因为植物的光合作用受到植物内部因素和外部条件的共同影响[9],外界环境因子的改变可能会导致同一植物光合作用速率不同,在不同的土壤水分和遮荫条件下,净光合速率不同,可能与毛红椿在不同地域以及栽植方式不同导致外界环境因子变化有关。同时,相关分析和多元逐步回归分析表明,对毛红椿夏季净光合速率影响最大的因子是气孔导度和光合有效辐射。
[1] 邹高顺.珍贵速生树种红椿与毛红椿引种栽培研究[J].福建林学院学报, 1989, 19(3): 263-271.
[2] 孟平,张劲松,高峻.山茱萸幼树光合及水分生理生态特性[J].林业科学研究, 2005, 18(1): 47-51.
[3] 张纪卯,康木水,连书钗.毛红椿扦插育苗试验[J].西南林学院学报. 2008, 28(6): 57-60,64.
[4] 陶丹,武来成,张露.毛红椿不同种源及家系间苗期生长变异性研究[J].南昌工程学院学报. 2007, 26(4): 9-13.
[5] 张汝忠,彭佳龙,王坚娅.毛红椿播种育苗技术及苗期生长规律研究[J].浙江林业科技. 2007, 27(4): 51-53.
[6] 张露,郭联华,杜天真, 等.遮阴和土壤水分对毛红椿幼苗光合特性的影响[J].南京林业大学学报:自然科学版,2006, 30(5):63-66.
[7] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Ann. Rev. Plant Physiol. 1982, 33: 317-345.
[8] Salvucci M E, Ports A R, Ogren W L. Light and CO2response of ribulose-1,5-bisphosphate carboxylase/oxygenase activation in arobidopsis leaves[J]. Plant Physiol. 1986, 80:65-5659.
[9] 王旭军,康向阳,吴际友.响叶杨光合速率及生理生态因子的日变化研究[J].湖南林业科技. 2008, 35(5): 5-8,12.
[10] Berry J A, Downton W J S. Environmental regulation of photosynthesis[C]//Govindjee ed. Photosynthesis, Vol.Ⅱ. Development, Carbon Metabolism and Plant Productivity. New York: Academic Press. 1982:263-343.
Diurnalvariationofphotosyntheticrateandeco-physiologicalfactorsofToonaciliatavar.pubescens
WU Jiyou1, CHENG Rui1, WANG Xujun1, CHENG Yong1,PENG Huiming2, YANG Guangwen2
(1.Hunan Academy of Forestry, Changsha 410004, China;2.Forestry Institute of Yueyang City, Yueyang 414000, China)
The diurnal variation of net photosynthetic rate (Pn) of two-year-oldToonaciliatavar.pubescensand its eco-physiological factors were investigated by using Li-cor 6400 portable photosynthesis system on August 2010. The results showed that the diurnal variation of net photosynthetic rate ofT.ciliatavar.pubescensin sunny day varied with double peak curve. The peaks ofPnoccurred at 8∶00 and 13∶00, respectively, showing obvious midday depression. The correlation analysis and regression analysis revealed that the dominantly influencing factors ofPnofT.ciliatavar.pubescenswere photosynthetically active radiation, stomatal conductance and atmospheric CO2concentration.
Toonaciliatavar.pubescens; photosynthetic rate; eco-physiological factors; diurnal variation
2011 — 02 — 15
2011 — 03 — 01
S 792.33
A
1003 — 5710(2011)02 — 0005 — 04
10. 3969/j. issn. 1003 — 5710. 2011. 02. 002
(责任编辑:唐效蓉)
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
农业工程学报(2022年3期)2022-04-16
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
山东林业科技(2019年2期)2019-06-03
新课程·下旬(2017年7期)2017-08-14
中国果业信息(2017年4期)2017-05-09
上海蔬菜(2015年2期)2015-12-26
西北林学院学报(2015年5期)2015-01-03
