
利用转座标签mTn-lacZ/leu2插入突变发酵性丝孢酵母2.1368-Leu−筛选高效产油突变株
2011-09-29 07:26:44宋安东刘玉博谢慧王风芹鲍晓明
生物工程学报 2011年3期
宋安东,刘玉博,谢慧,王风芹,鲍晓明
1 河南农业大学生命科学学院,郑州 450002
2 山东大学微生物技术国家重点实验室,济南 250100
利用转座标签mTn-lacZ/leu2插入突变发酵性丝孢酵母2.1368-Leu−筛选高效产油突变株
宋安东1*,刘玉博1*,谢慧1,王风芹1,鲍晓明2
1 河南农业大学生命科学学院,郑州 450002
2 山东大学微生物技术国家重点实验室,济南 250100
为提高微生物油脂产率,降低其生产成本,以转座标签 mTn-lacZ/leu2插入突变发酵性丝孢酵母 2.1368-Leu−筛选高效产油突变株。利用LacZ显色反应、脂肪酸合成酶抑制剂Cerulenin和磷酸香草醛反应,最终在玉米秸秆糖化液中筛选出一株高效产油突变株2.1368-Leu−-7。结果表明其油脂含量为38.30%,比对照的29.33%高了8.97%,而其产油率为8.35%,比对照的6.92%提高了20.63%;在玉米秸秆糖化液中的糖利用率为77%,每100 g玉米秸秆可转化油脂8.32 g。可为未来生物柴油产业提供了廉价原料。
转座标签,磷酸香草醛反应,玉米秸秆糖化液,生物柴油
Abstract:To improve microbial lipid production, we inserted mTn-lacZ/leu2 into Trichosporon fermentans 2.1368-Leu−to obtain high lipid production mutants. By observing the LacZ chromogenic change, the positive reaction between Cerulenin (inhibitor of fatty acid synthase) and phosphate vanillin, a higher lipid-producing mutant 2.1368-Leu−-7 grown on corn-stalk hydrolysate was obtained. The lipid content of this mutant reached 38.30% (8.97% higher than that of the control) and the lipid production rate was 8.35% (20.63% higher than that of the control). The rate of sugar utilization was 77%, meaning that 100 g corn-stalk could be converted to 8.32 g lipid. The study provided an effective method for microbial lipid production by using cheap raw materials for biodiesel.
Keywords:Trichosporon fermentans, transposon tagging, corn stalk, biodiesel
生物柴油是典型的“绿色能源”,大力发展生物柴油对发展低碳经济、推进能源替代和减轻环境压力具有重要的战略意义[1-2]。当前利用动植物油脂生产生物柴油,原料成本偏高,并且稳定、充足的油脂原料供应体系尚未形成。而微生物油脂发酵周期短,不受场地、季节、气候变化等因素的影响;且产油微生物菌种资源丰富,能利用和转化各种农林废弃木质纤维素原材料,对农业大国具有特殊的意义。因此,利用微生物转化法获取油脂具有非常大的发展潜力[3-4]。
转座子标签技术是根据转座子随机插入突变的特性而发展起来的研究基因功能的新方法。山东大学微生物技术国家重点实验室利用 mTn3转座标签对酿酒酵母Saccharomyces cerevisiae W303.1A进行诱变、筛选得到突变体263-H9,该突变体表现出对多种逆境胁迫敏感的表型特征,并最终确定了263-H9突变体的盐敏感相关基因[5]。转座子标签技术主要用于植物基因组和功能序列的分离与研究。任争光等[6]利用转座子 Mini-Tn5构建致病菌株MH21的突变体库,筛选得到 1株致病性完全丧失的突变体M543。对M543中转座子插入基因的克隆和测序表明其突变基因为燕麦食酸菌Ⅲ型分泌系统(TTSS) 中的保守基因hrcR[6]。
发酵性丝孢酵母 Trichosporon fermentans 2.1368,酵母目隐球酵母科毛孢酵母属,属于茁芽毛孢酵母的一种,是可以进行全糖发酵的产油酵母,具有易培养、木糖转化率高等特点。近几年,对发酵性丝孢酵母2.1368的研究主要集中在发酵条件的优化,而对此菌株的改造尚未见报道。用浓度为 1.0 mg/mL的亚硝基胍,处理 1.0 h,诱变2.1368,得到2.1368的亮氨酸缺陷型2.1368-Leu−。本研究以2.1368的亮氨酸缺陷型2.1368-Leu−为出发菌株,采用转座标签 mTn-lacZ/leu2插入突变的方法,筛选出一株在玉米秸秆糖化液发酵中产油率明显提高的突变株。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒
菌种:发酵性丝孢酵母Trichosporon fermentans 2.1368,购于中国工业微生物菌种保藏中心;发酵性丝孢酵母Trichosporon fermentans 2.1368的亮氨酸营养缺陷型2.1368-Leu−,本实验室筛选;大肠杆菌DH5α,河南农业大学微生物学系保藏菌种。
质粒:mTn3转座子标签系列的 mTn-lacZ/leu2转座标签插入文库质粒15个,编号分别为P21-P29、P31、P34-P38,山东大学鲍晓明教授赠送。mTn-lacZ/leu2转座子标签插入示意图如图1所示。
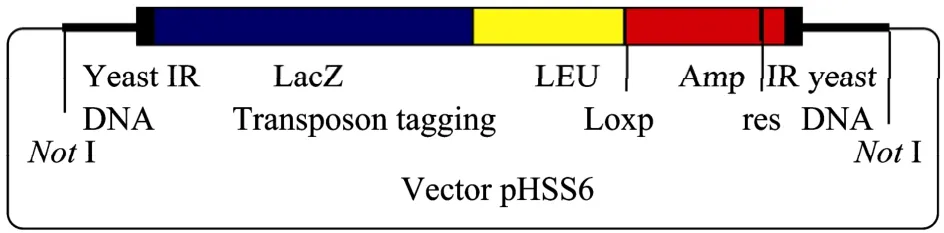
图1 mTn-lacZ/leu2转座子标签插入文库Fig. 1 Pools of the yeast genomic library with mTn-lacZ/leu2 transposon tagging.
转座子标签插入文库是以 mTn3转座标签对构建于载体 pHSS6上的酿酒酵母 Saccharomyces cerevisiae染色体文库进行随机插入而得。它具有38 bp的末端重复序列、res位点与lox位点,并加入了 Amp抗性标记、Leu缺陷型标记和没有启动子的报道基因LacZ,以及2个相同的Not І酶切位点[7]。
1.1.2 试剂与溶液
2.24×103mol/L Cerulenin (浅蓝菌素) 母液:5 mg Cerulenin充分溶解于10 mL DMSO (二甲基亚砜) 中。
磷酸香草醛试剂:0.12 g香草醛溶解于20 mL蒸馏水中,用85%的磷酸定容至100 mL。
1.1.3 培养基
发酵性丝孢酵母2.1368菌种活化培养基 (麦芽汁培养基):糖度为12-13波美度的麦芽汁,2%琼脂。
2.1368营养缺陷型筛选基本培养基 (SD)[8](g/100 mL):酵母氮源 (不含氨基酸) 0.67,葡萄糖2,琼脂2,硫酸铵 0.5,pH 6.0,(或加腺嘌呤80 mg/L)。
YAPD培养基 (g/100 mL):酵母浸膏1,蛋白胨2,葡萄糖2,腺嘌呤80 mg/L,琼脂2 (YAPD培养基加腺嘌呤可抑制ade1和ade2突变株的回复突变)。
X-gal显色平板培养基(g/100 mL):0.6 mL X-gal贮存液,Na2HPO43.5814,硫酸镁0.0245,琼脂1.6,pH 7.0。
发酵性丝孢酵母2.1368产脂培养基 (g/100 mL):葡萄糖12,酵母膏0.8,蛋白胨0.3,KH2PO40.7,MgSO4·7H2O 0.1,乙醇 0.2,pH 5.8。
玉米秸秆糖化液发酵培养基:用体积分数为1%的稀H2SO4于121 ℃预处理60 min后,加入蒸馏水调节 pH值至 5.0,使固液比达到 10∶1,然后加入纤维素酶25 g/kg和木聚糖酶液20 mL/kg,120 r/min、50 ℃处理48 h,过滤除渣得糖化液。糖化液中总还原糖浓度为 5.2%,其中葡萄糖浓度为3.9%,木糖为1.2%,其余为杂糖。
1.2 试验方法
1.2.1 转座标签文库质粒插入片段的获得
将15个mTn-lacZ/leu2转座标签插入文库质粒分别转化大肠杆菌DH5α,挑取单菌落提取质粒,将质粒用Not I酶切,并回收转座子标签文库质粒的插入片段 (8 kb左右)。
1.2.2 酵母的高效转化方法[9]
将菌种 2.1368-Leu−用 100 mmol/L醋酸锂(LiAc) 处理并悬浮细胞,每 100 µL等份分装到1.5 mL EP管,离心后除去LiAc。按如下量加入转化混合液:PEG 3350 (50%,W/V),240 µL;1.0 mol/L LiAc,36 µL;单链载体 DNA (2.0 mg/mL),25 µL;水和质粒DNA,50 µL;剧烈振荡后置28 ℃保温30 min转化酵母,然后于42 ℃水浴中热激25 min。10 000 r/min高速离心30 s除去转化混合液后,吸0.4 mL无菌水加到每个EP管中,悬浮沉淀,用等份的200 µL转化混合液涂布选择平板SD,28 ℃培养3 d左右。
本试验用 P26号质粒酶切回收片段为例研究2.1368-Leu−的最佳转化热激时间,设计10、15、20、25、30、35、40 min为热激时间梯度。
1.2.3 同源重组质粒的筛选
挑取 SD平板上生长出来的转化子落涂片于YAPD平板上,每板的密度为100个铺片,28 ℃条件下培养24 h,用一张无菌滤纸将该平板复制于另一加有腺嘌呤的 SD平板上,28 ℃过夜。将滤纸取出,克隆面向上,置于一密闭容器中,容器底部加入10 mL氯仿处理10~30 min,取出滤纸,再将其克隆面向上置于X-gal平板 (120 µg/mL X-gal,0.1 mol/L Na2PO4和1 mmol/L MgSO41.6% 琼脂糖)上,28 ℃条件下显色1 d。
1.2.4 浅蓝菌素Cerulenin初筛高产油脂酵母
确定2.1368原始菌株对Cerulenin的最小抑制浓度,即在每 10 mL 2.1368固体培养基中分别加入2.24×10−3mol/L的 cerulenin母液 0、10、20、30、40、50、60、70、80 µL制成梯度培养基,按照常规方法涂布平板,统计cerulenin对菌体菌落形成率的影响。
1.2.5 磷酸香草醛反应筛选高产油脂突变株[10-11]
对初筛菌株进行产脂培养后,取10 mL菌液,4 000 r/min离心10 min,蒸馏水洗1~2次。定容至2 mL,取100 µL (对照取蒸馏水) 加入一带塞试管中,加入18 mol/L的H2SO42 mL,沸水浴中孵化10 min,常温水浴5 min,加入5 mL磷酸香草醛试剂,37 ℃保温15 min,常温水浴10 min,于530 nm测其OD值。
1.2.6 生物量的测定
通过4 500 r/min离心l0 min,收集菌体并干燥(干燥温度:80 ℃) 至衡重,准确称取干菌体量。
1.2.7 发酵验证试验[12]
对所筛突变株在玉米秸秆糖化液中进行发酵试验验证。其中
干菌体含油率=粗油脂重量/干菌体重量×100%。产油率=粗油脂重量/加入糖化液糖质量×100%。
2 结果与分析
2.1 所提取的转座标签插入文库质粒及其 NotⅠ酶切结果
将 15个转座标签插入文库质粒转化大肠杆菌DH5α、挑取阳性克隆提取质粒备用。纯化后的质粒经NotⅠ酶切,相应质粒的酶切产物所得到的两条带。含有酵母同源序列和 mTn-lacZ/leu2转座子的片段(6 654 bp) 不均一,约8~10 kb大小,而不含有酵母同源序列的载体pHSS6片段则大小一致,为2.1 kb。
2.2 发酵性丝孢酵母2.1368-Leu−的高效转化
用回收后的DNA片段进行酵母高效转化,确定最佳热激时间为20 min,适宜的热激时间范围扩大至15~35 min。将15个转座子标签插入文库质粒采用以上转化条件 (即热激时间为20 min或25 min)转化2.1368-Leu−,在SD筛选平板上得到的转化子情况如表1所示。
由表1可以看出,经过转化试验共得到166个转化子,其中成功实现转化的质粒为P22、P24、P25、P26、P27、P29、P31、P34、P35、P37、P38,用所得到的15个转座子标签文库质粒进行酶切转化,有4个没有实现成功转化。
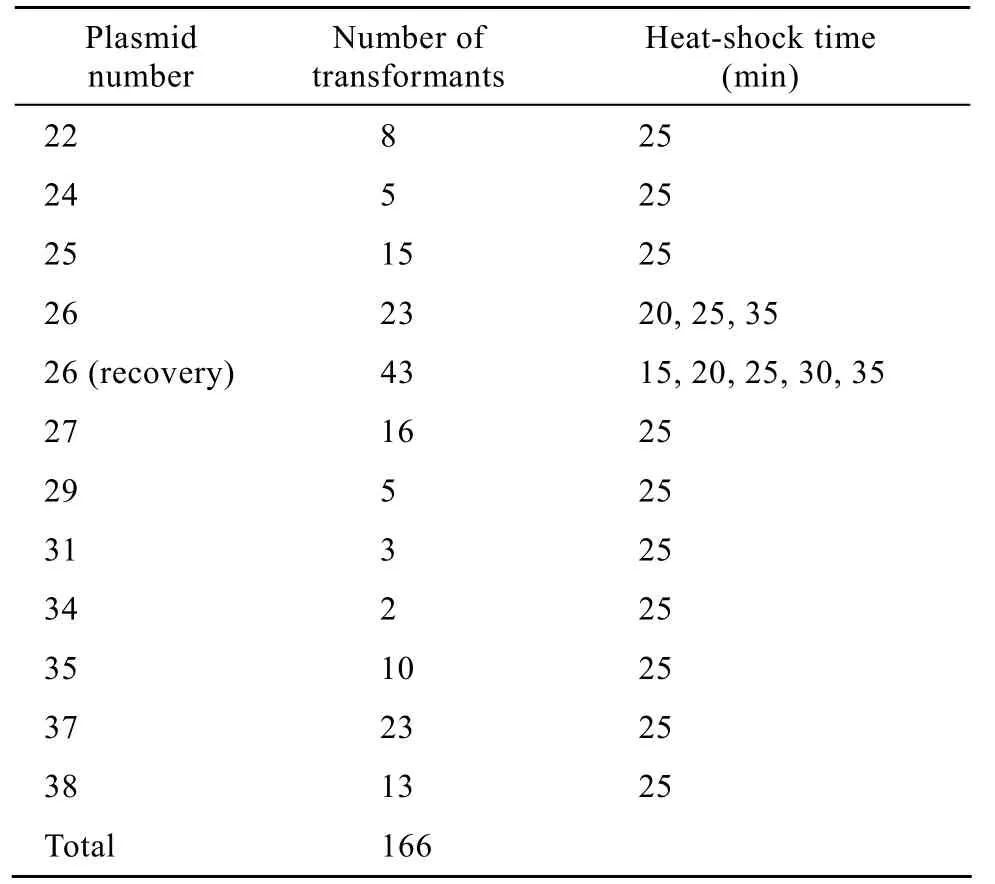
表1 酵母高效转化所得转化子情况Table 1 Transformants gained by yeast efficient transformation
试验中所用的转座子的报道基因 LacZ没有启动子,缺失了前端的ATG序列,载体在Not I线性化之后重组片段自身就不携带启动子,转座子系统中,LacZ基因下游有—LEU基因,因此选用亮氨酸缺陷型的酵母菌株作为重组菌株,该菌株不能够在缺少亮氨酸的 SD培养基上生长,只在含有上述转座系统的质粒同源重组到酵母中,并且有启动子在其上游启动它转录,且能够按三联密码子正确读框,该重组酵母菌株才能够编码合成亮氨酸,从而在亮氨酸缺陷型的SD平板上生长。
2.3 同源重组质粒的筛选
利用LacZ显色反应,根据其显色水平,可以判断LacZ基因前端的启动子强弱。在含X-gal平板上对试验中得到的 166个转化子进行同源重组子筛选,结果如图2所示。
从图2中可以看出,P26-3和P26-5号不显蓝色(图中实箭头所指),其他P26号质粒的转化子蓝色都较深,说明这两个转化子不是同源重组子,可能是阅读框架错误,应该是试验误差;P35和 P25号质粒的转化子蓝色较弱,其中P25-5基本不显蓝色 (图中方框处),可能因为 LacZ基因上游的启动子太弱,不能很好完成转录所致。因此,在用mTn-lacZ/leu2转座标签对发酵性丝孢酵母进行插入突变过程中,P22、P24、P26、P27、P29、P31、P34、P37 和 P38这9个质粒是比较适合的同源载体。筛选166个转化子,最终得到 139个同源重组子,可以作为筛选高产突变株的出发菌株。
2.4 优良重组子的筛选
2.4.1 Cerulenin初筛高产油脂突变株
Cerulenin是一种蓝色头孢霉的自然代谢产物,它通过与脂肪酸合成酶中 β-酮脂酰-ACP合酶末端丝氨酸上的-SH基结合,形成羟基内酰胺环而使之失活。β-酮脂酰-ACP合酶是脂肪酸合成的关键酶,故低浓度的Cerulenin能够不可逆地抑制生物体的内源性脂肪酸合成,从而抑制细胞的生长代谢。在一定浓度的Cerulenin培养基上,只有脂肪酸合成酶活性比较高的菌株才能存活下来而被选出[11]。

图2 单菌落酵母转化子在X-gal板上的显色结果Fig. 2 The color result of transformant colony on X-gal plate.(a) The number of P38plasmid (13), P26(7), P31(3), P34(2). (b)The number of P37plasmid (23). (c) The number of P24plasmid(5), P26 (recovery) (43). (d) The number of P35plasmid (10),P25 (15), P22 (8). (e) The number of P26plasmid (16), P27 (16),P29 (5).
经试验确定当2.24×10−3mol/L Cerulenin的量达到60 µL时,菌落形成率约为3%,以此浓度作为筛选最佳剂量。得到的重组子中,在Cerulenin作用下有部分菌落比出发菌株存活率高,相同Cerulenin抑制浓度下,选取菌落形成率比出发菌株大的作为初筛菌株,从32株菌中得到了15株可能高产油脂的突变株。
2.4.2 磷酸香草醛反应法分析初筛突变株
经过发酵产脂的酵母经处理后进行磷酸香草醛反应,会生成一种有色物质,其反应体系的颜色从浅绿色到茶红色,随油脂含量的增加而加深[10]。将初筛后的15个重组子经过产脂培养,其菌体通过磷酸香草醛反应后,反应颜色有明显变化,根据颜色深浅可以判断其含油率高低,从15株初筛突变株中选出10株颜色较深的突变株进行OD值测定和油脂含量测定。
发酵产脂的酵母经处理后进行磷酸香草醛反应后,其在530 nm处的光吸收值与氯仿甲醇抽提法测得的菌体油脂含量成正比,测得结果见表2。
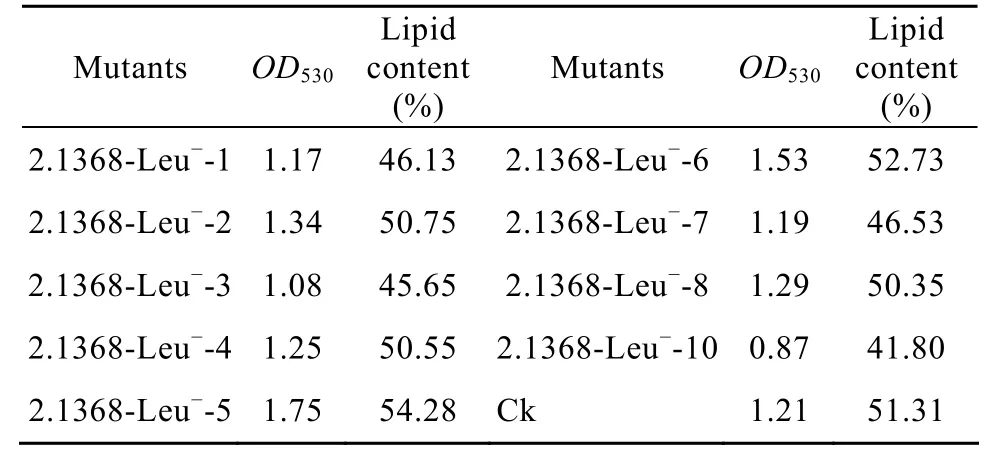
表2 磷酸香草醛反应法测定的初筛突变株OD530值和油脂含量Table 2 OD530results and lipid content results of primary screening mutants by the methods of phosphate and vanillin reaction
由表 2 可以看出,2.1368-Leu−-1、2.1368-Leu−-2、2.1368-Leu−-4 、 2.1368-Leu−-5 、 2.1368-Leu−-16 、2.1368-Leu−-7、2.1368-Leu−-8这几个突变株的 OD值和油脂含量大于Ck (原始菌株) 或者和对照相当,并且各重组子在 530 nm处的光吸收值与油脂含量基本上呈正相关。所以筛选这几株菌进行玉米秸秆糖化液发酵试验。
2.4.3 发酵试验复筛高产突变株
将上述 7个突变株进行玉米秸秆糖化液发酵试验,研究这7株突变对玉米秸秆糖化液的耐受性及利用纤维质原料生产微生物油脂的转化效率。发酵结果如表3所示。
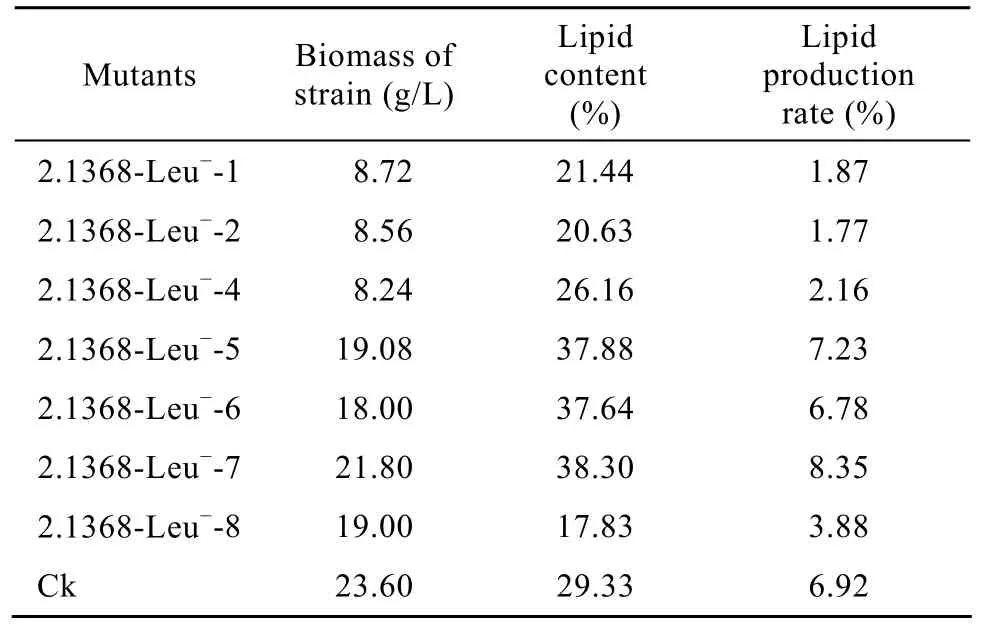
表3 突变株在秸秆糖化液培养基中的发酵结果Table 3 The fermentation results of the mutants under stalk saccharified liquid
由表3可以看出,发酵后这7个突变株的生物量均低于对照,其中突变株 2.1368-Leu−-1、2.1368-Leu−-2、2.1368-Leu−-4 的生物量低于 10 g/L,可能是由于突变株对秸秆糖化液中的有毒物质比较敏感,导致生长受到抑制;突变株2.1368-Leu−-7的油脂含量最高,为 38.30%,比对照的 29.33%高了8.97%,并且2.1368-Leu−-7的产油率最高,为8.35%,而对照为6.92%,产油率比对照提高了20.63%。可以确定2.1368-Leu−-7为最佳高效产油突变株。
在2.1368-Leu−-7进行玉米秸秆糖化液的发酵试验中,其残余总还原糖为1.20%,糖利用率为77%;每g糖可转化油脂1.60 g;每100 g秸秆转化油脂8.32 g。为微生物油脂的工业化奠定基础。
3 讨论
通过试验,确定最佳热激时间为20 min。将15个转座子标签插入文库质粒在最佳热激时间下转化2.1368-Leu−,在 SD筛选平板上得到的 166株转化子。其中成功实现转化的质粒为P22、P24、P25、P26、P27、P29、P31、P34、P35、P37、P38。
利用LacZ显色反应,根据其显色水平,对166株转化子进行同源重组载体进行筛选,得到 139株同源重组子。作为筛选高产菌的出发菌株。
利用Cerulenin和磷酸香草醛反应筛选高产油脂酵母菌,并对初筛菌株进行玉米秸秆糖化液发酵试验验证。最终确定 7号菌为最佳高效产油菌株,其油脂含量为38.30%,比对照的29.33%高了8.97%,而其产油率为 8.35%,比对照的 6.922%提高了20.63%;而在玉米秸秆糖化液中残余总还原糖为1.20%,糖利用率为77%;每g糖可转化油脂1.60 g;每100 g秸秆转化油脂8.32 g。
本文参考文献[9]和[13],将几种酵母转化方法进行比较和改进,最终确定了最佳转化方法。在酵母转化过程中发现导致转化率低的因素很多,如PEG浓度、热激时间、单链载体、质粒 DNA浓度等。通过多次试验发现,混合液中质粒 DNA添加4~5 µL (0.6 µg左右) 最佳,浓度较高时转化效果并不理想,这可能是线性化质粒串联整合所致。
利用Cerulenin筛选高产油脂酵母,可以实现酵母的高通量筛选,方便易行,是一种比较理想的油脂酵母筛选方法。在磷酸香草醛反应中,生物体与磷酸香草醛试剂反应,生成一种红色物质,其机理尚不清楚,但根据反应颜色的深浅即可以定性分析油脂含量的高低,该方法需要的生物量小,且无须细胞破碎便可直接对细胞油脂含量进行定性分析,快速方便,具有很好的应用前景。
本文仅完成了利用转座子标签插入突变筛选高产油脂突变株 2.1368-Leu−-7,初步分析该突变株仍是发酵性丝孢酵母。突变株的获得,这只是酵母功能分析的第一步。要对酵母突变株进一步研究,需要通过拯救质粒pRSQ2-URA (实验室已具有) 法确定转座子标签插入位点,根据拯救质粒全序列设计并合成引物对验证过的质粒测序,将测序结果与酿酒酵母基因组全序列数据库进行Blast分析,确定各转座标签的插入位点,进而对插入位点基因进行深入分析。
REFERENCES
[1] Corma A, Iborra S, Velty A, et al. Chemical routes for the transformation of biomass into chemicals. Chem Rev,2007, 107(6): 2411−2502.
[2] Service RF. Cellulosic ethanol: biofuel researchers prepare to reap a new harvest. Science, 2007, 315(5818):1488−1491.
[3] Zhao ZB, Hua YY, Liu B. How to secure triacylglycerol supply for chinese biodiesel nndustry. China Biotechnol,2005, 25(11): 1−6.
赵宗保, 华艳艳, 刘波. 中国如何突破生物柴油产业的原料瓶颈. 中国生物工程杂志, 2005, 25(11): 1−6.
[4] Mo YX, Liu HJ, Zhang JA, et al. Research on production of lipids by microbial fermentation. Renewable Energy Res, 2006, 130(6): 24−28, 32.
墨玉欣, 刘宏娟, 张建安, 等. 微生物发酵制备油脂的研究. 可再生能源, 2006, 1303(6): 24−28, 32.
[5] Yu DK, Zhang XH, Liu XY, et al. Identification of the gene correlated with salt stress in the Saccharomyces cerevisiae 263-H9 mutant. Hereditas, 2006, 28(10):1294−1298.
于典科, 张小华, 刘向勇, 等. 酿酒酵母转座标签插入突变体 263-H9中高盐胁迫基因的确定. 遗传, 2006,28(10): 1294−1298.
[6] Ren ZG, Hou L, Song ZG, et al. Screening of the pathogenicity mutants of Acidovorax avenae subsp.citrulli and cloning of the hrcR gene. Acta Phytopathol Sin, 2009, 39(5): 501−506.
任争光, 侯磊, 宋治国, 等. 甜瓜细菌性果斑病菌致病性突变体筛选与hrcR基因的克隆. 植物病理学报, 2009,39(5): 501−506.
[7] Ross-Macdonald P, Coelho PS, Roemer T, et al.Large-scale analysis of the yeast genome by transposon tagging and gene disruption. Nature, 1999, 402(6760):413−418.
[8] de Jesus Ferreira MC, Bao XM, Laizé V, et al. Transposon mutagenesis reveals novel loci affecting tolerance to salt stress and growth at low temperature. Curr Genet, 2001,40(1): 27−39.
[9] Adams A, Gottschling DE, Kaiser CA, et al. Methods in Yeast Genetics. Beijing: Science Press, 2000.
[10] Izard J, Limberger RJ. Rapid screening methods for quantity of bacterial cell lipids from whole cells. J Microbiol Methods, 2003, 55(2): 411−418.
[11] Li RM, Wang JF, Ma S, et al. Selection of yeast high in lipids-yield by fatty acid synthetase inhibitor and phosphoric acid-vanillin reaction. Microbiol China, 2008,35(4): 545−549.
李仁民, 王菊芳, 马爽, 等. 利用脂肪酸合成酶抑制剂和磷酸香草醛反应筛选高产油脂酵母菌. 微生物学通报, 2008, 35(4): 545−549.
[12] Cao J, Wang CH, Zeng S, et al. Comparison of several methods for extracting oil from Mucor circinelloides 3.2208. China Oils Fats, 2004, 27(4): 38−40.
曹健, 汪晨辉, 曾实, 等. 卷枝毛霉Mucor circinelloides 3.2208油脂几种提取方法的比较. 中国油脂, 2004,27(4): 38−40.
[13] Gietz RD, Schiestl RH. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nature Protocols, 2007, 2(1): 31−34.
Journals.im.ac.cn
Screening of high lipid production Trichosporon fermentans mutants by transposon tagging mTn-lacZ/leu2 insertion
Andong Song1*, Yubo Liu1*, Hui Xie1, Fengqin Wang1, and Xiaoming Bao2
1 Life Science College, Henan Agricultural University, Zhengzhou 450002, China
2 State Key Laboratory of Microbial Technology, Shandong University, Jinan 250100, China
Received: November 24, 2010; Accepted: February 23, 2011
Supported by: Key Scientific and Technological Projects of Henan Province (No. 092102210110).Corresponding author: Xiaoming Bao. Tel: +86-531-88365826; E-mail: bxm@shu.edu.cn Andong Song. Tel: +86-371-63555153; Fax: +86-371-63555790; E-mail: song1666@126.com
*These authors contributed equally to this study.
河南省重点科技攻关项目 (No. 092102210110) 资助。
猜你喜欢
工程塑料应用(2023年12期)2024-01-04 12:38:20
生物加工过程(2023年4期)2023-07-27 07:42:12
食品工业科技(2022年14期)2022-08-03 15:26:34
林业科学(2022年1期)2022-03-23 06:56:24
中国蜂业(2021年5期)2021-05-22 02:59:26
酒·饮料技术装备(2018年1期)2018-04-28 09:09:08
浙江农林大学学报(2016年6期)2016-12-12 12:01:32
中国粮油学报(2016年5期)2016-01-23 02:45:00
中国卫生标准管理(2015年25期)2016-01-14 09:29:30
邵阳学院学报(自然科学版)(2015年2期)2015-06-05 12:22:39
