
西施舌群体遗传结构及分化的RAPD分析
2011-09-24 08:11孟学平高如承程汉良
海洋科学 2011年2期
孟学平, 王 帅, 高如承, 申 欣, 程汉良, 田 美
(1. 淮海工学院 海洋学院, 江苏 连云港 222005; 2. 福建师范大学 生命科学学院, 福建 福州 350108)
西施舌群体遗传结构及分化的RAPD分析
孟学平1, 王 帅2, 高如承2, 申 欣1, 程汉良1, 田 美1
(1. 淮海工学院 海洋学院, 江苏 连云港 222005; 2. 福建师范大学 生命科学学院, 福建 福州 350108)
采用55个随机引物对西施舌(Coelomactra antiquata)5个群体, 即长乐(CL)、启东(QD)、连云港(LYG)、胶南(JN)和北海(BH)群体进行RAPD 扩增。用PopGen(Version1.32)软件进行遗传多样性分析。结果共筛选出12个重复性好的引物, 得到88条清晰稳定的条带, 扩增片段在100 bp~3 000 bp之间, 其中引物S299扩增的600 bp片段为长乐群体特有。多态位点比例为62.07%~78.26%(JN、LYG、BH、QD、CL群体分别为 78.26%、77.05%、72.58%、70.05%、62.07%), Shannon指数为0.210 5~0.312 3。群体间遗传距离为0.068 3~0.223 9, 其中长乐群体与其余4个群体间的遗传距离为0.182 7~0.223 9, 4个非长乐群体的遗传距离为0.068 3~0.136 7。群体间遗传分化系数(Fst)为 0.313 01(P<0.05), 表明在整个遗传变异中群体间占31.30%。CL群体与QD, BH, JN, LYG群体间的遗传分化系数(Gst)分别为0.364 9,0.344 8, 0.325 0, 0.309 0; 而非长乐群体之间的遗传分化系数为0.148 2~0.240 3; 以上数据显示长乐群体遗传分化最大。聚类结果显示, 启东和胶南群体首先聚为一支, 然后与北海群体聚在一起, 再和连云港群体相聚; 而长乐群体则单独形成一支。
西施舌(Coelomactra antiquata); 遗传结构; RAPD
西施舌(Coelomactra antiquata)为浅海埋栖性双壳贝类, 隶属于软体动物门(Mollusca)、双壳纲(Bivalvia)、帘蛤目(Veneroida)、蛤蜊科(Mactridae)。西施舌自然分布于太平洋西部的日本、朝鲜、韩国及中国沿海和印度半岛。我国南北沿海均有分布。西施舌营养价值高[1], 养殖前景看好。近10多年来,由于需求量的不断增加, 各种掠夺性的捕捞和环境污染, 使西施舌产量逐年减少[2-3]。为了保护开发这种名贵贝类, 自20世纪60年代起, 国内学者便开始对西施舌的生物学[4]、人工育苗[5]等进行了研究。近年来, 群体遗传差异研究报道日渐增多[6-10], 但全国西施舌总体遗传关系尚不明朗。且目前报道的核苷酸多态性研究结果不能解析大多数群体间的差异。随机引物扩增多态性 DNA(RAPD)分析技术已广泛应用于水产动物的遗传背景研究[11-13]。本研究用RAPD分子标记分析我国 5个自然群体西施舌遗传差异, 对存在的争议作深入研究, 进一步揭示西施舌群体间的遗传差异, 为其遗传多样性分析、种质鉴定及其资源保护和利用提供核基因组DNA资料。
1 材料与方法
1.1 材料和试剂
实验样本于2004年10月至2007年5月分别采自山东胶南(JN)、江苏连云港(LYG)、启东(QD)、福建长乐(CL)、广西北海(BH)5海区的野生西施舌。取闭壳肌浸于75%乙醇中, 4°C保存备用, 每个群体30个样本。
DNA提取试剂、Taq酶(BBI)等 PCR试剂购自生工生物工程(上海)有限公司。
1.2 DNA提取及浓度检测
将酒精浸泡的西施舌闭壳肌(或斧足), 洗涤除去乙醇吸干表面水分, 称取50~100 mg, 剪碎, CTAB裂解液加蛋白酶 K裂解, 酚-氯仿抽提, 乙醇沉淀,TE缓冲液溶解。DNA经电泳检测, 用核酸蛋白检测仪测定浓度后备用。
1.3 PCR扩增
实验所用的 12个随机引物从生工生物工程(上海)有限公司合成的55个引物中筛选得到, 引物编号及其序列见表1。
PCR 反应总体积 25 μL, 其中含 10×Buffer2.5 μL,MgCl22.5 mmol/L, dNTP 0.2 mmol/L, 引物 0.5 μmol/L,Taq酶1.0 U, 50 ng模板。扩增条件: 94 ℃预变性4 min,然后进入PCR循环, 即: 94 ℃变性45 s, 36 ℃退火45 s,72 ℃延伸1.5 min, 共进行40个循环。最后72 ℃延伸8 min。扩增产物用 1.5%琼脂糖凝胶电泳分离,GoldView染色, 凝胶成像系统观察并拍照。

表1 随机引物及其序列Tab. 1 RAPD primers and sequences
1.4 数据处理
电泳结果按照条带的有无转化为 1, 0距阵: 将电泳图谱中每一条带的迁移位置记作一个位点, 当相同迁移位置的扩增带出现时记为1, 缺失记为0。将1、0谱带矩阵输入PopGen(Version1.32)[14]软件进行Hardy-Weinberg平衡定律检验(卡方检验)、计算群体多态位点率、Shannon信息指数、Gst等相关参数及进行聚类分析。用Arlequin 3.1软件进行分子方差分析(AMOVA)。
2 结果与分析
2.1 PCR扩增
从供试的55个随机引物中筛选出条带清晰、稳定、分布合理、重复性好、个体或群体特异性引物12个, 共扩增出 88个条带, 扩增产物分子量为100~3000 bp。其中引物S299扩增效果好, 能将长乐与其他群体分开。长乐群体多数个体有特征带。部分RAPD图谱见图1。
2.2 遗传多样性分析
西施舌 5个群体的多态位点百分率在62.07%~78.26%之间, Shannon’S 信息指数Gst为0.2105~0.3123, 其中CL群体两个多样性指数最低,JN群体最高(表2)。
群体间的遗传距离显示(表 3), 长乐群体与其余4个群体间的遗传距离为0.182 7~ 0.223 9, 遗传相似度为0.799 4~0.833 0; 而非长乐的4个群体的遗传距离为0.068 3~0.136 7, 遗传相似度为0.872 2~0.934 0。这表明长乐群体和其他群体的亲缘关系相对远。
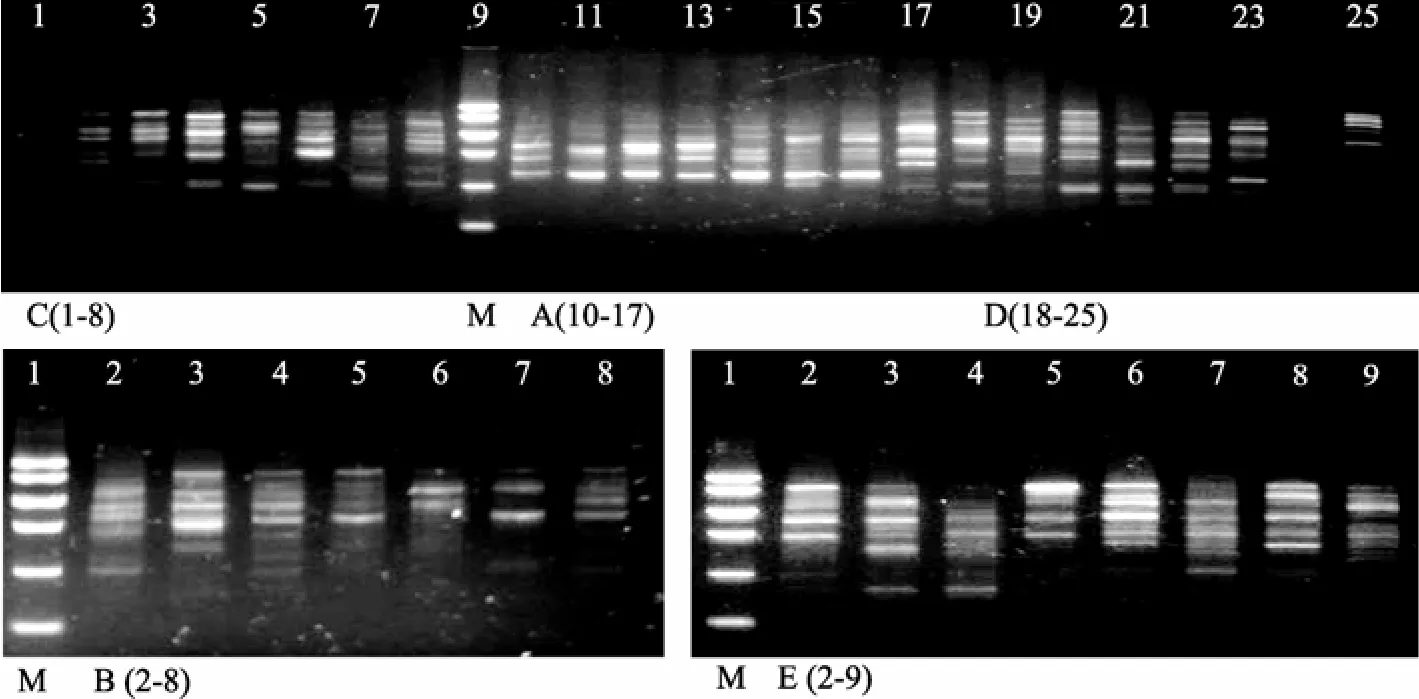
图1 引物S299对西施舌不同地理群体的RAPD扩增结果Fig. 1 PCR results of five stocks ofCoelomactra antiquatausing primer S299
AMOVA 分析结果(表 4)显示: 群体间遗传分化系数Fst=0.313 01(P<0.05), 表明在整个遗传变异中群体间占 31.30%, 遗传分化大部分来自群体内。由表5可知, CL群体与QD, BH, JN, LYG群体间的遗传分化值分别为0.364 9, 0.344 8, 0.325 0,0.309 0; 而QD, BH, JN, LYG 4个群体之间的Gst值为 0.148 2~0.240 3; 以上数据显示长乐群体和其他群体间的遗传分化最大, 群体间的基因流(Nm)同样呈这一趋势。
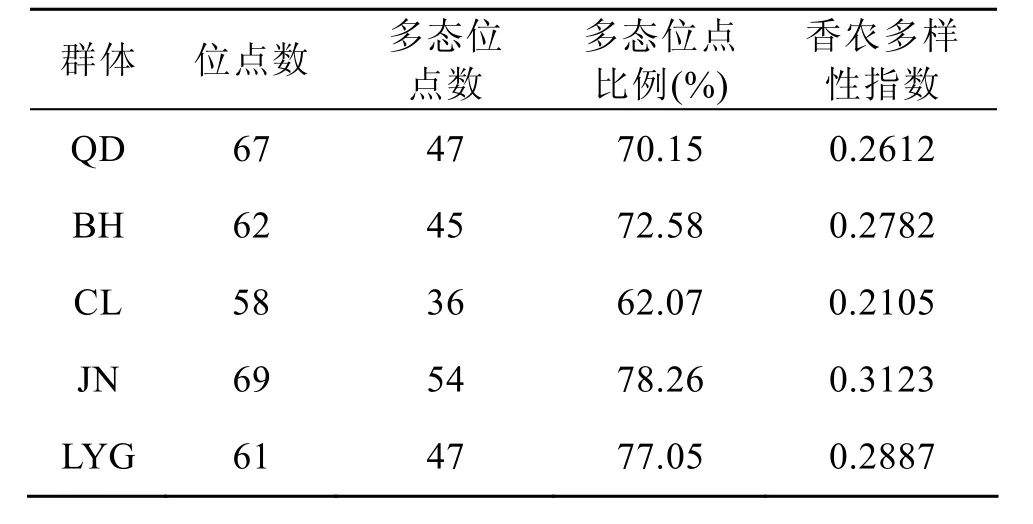
表2 西施舌群体遗传多样性Tab. 2 Genetic diversity ofC. antiquata
2.3 聚类分析
用 Popgen(Version1.3.1)软件构建 5个西施舌群体的UPMGA系统进化树。由图2可见启东和胶南群体首先聚为一支, 然后与北海群体聚在一起, 再和连云港群体相聚; 而长乐群体则单独形成一支。聚类结果显示北部湾(BH)群体与黄海(QD、JN、LYG)群体亲缘关系近, 与东海群体(CL)亲缘关系远, 其演化关系不规律。
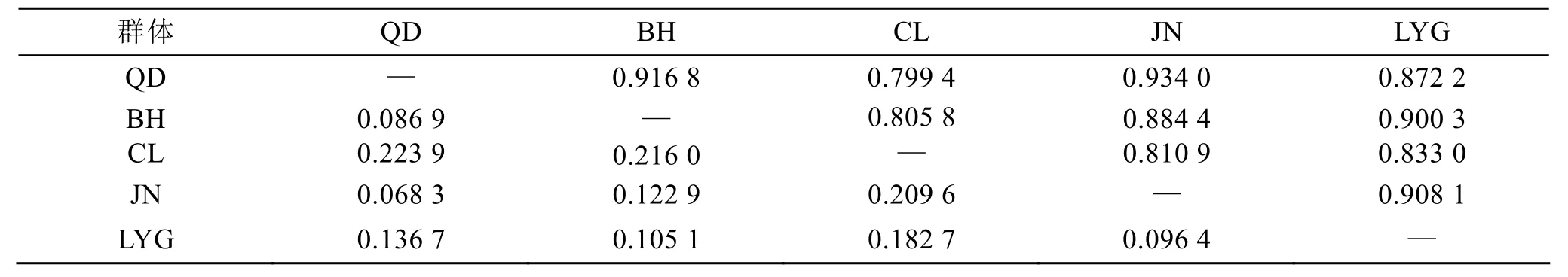
表3 西施舌群体间遗传距离(下三角)及遗传相似性指数(上三角)Tab. 3 Genetic distances and identities ofCoelomactra atiquatastocks
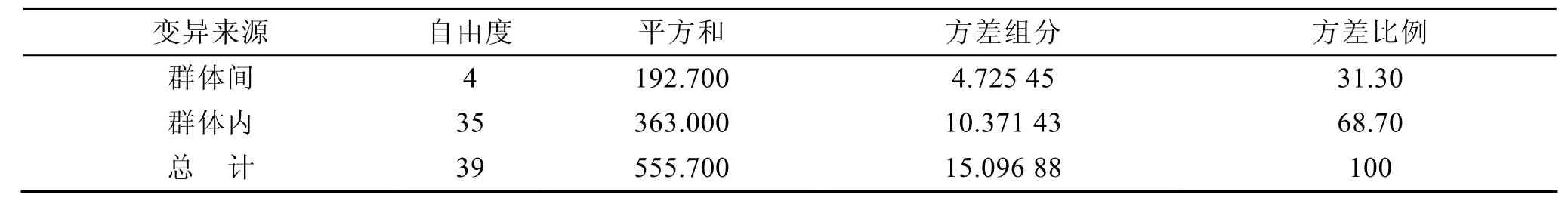
表4 群体间分子变异分析Tab. 4 Analysis of molecular variance (AMOVA) among stocks
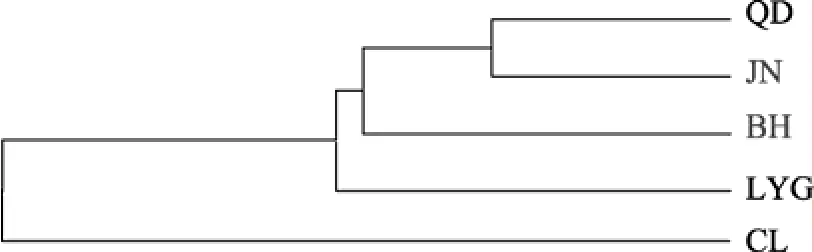
图2 基于RAPD的西施舌5个地理群体的UPGMA分子系统树Fig. 2 The UPMGA phylogenetic tree of five stocks ofCoelomactra antiquatabased on RAPD marker
3 讨论
本研究显示西施舌 5个自然群体多态位点比例(62.07%~78.26%)和香农信息指数(0.210 5~0.312 3)都比较高, 说明西施舌遗传多样性较高。5个自然群体间遗传距离为0.068 3~0.223 9。其中长乐群体与其余4个群体间的遗传距离为0.182 7~ 0.223 9, 4个非长乐群体的遗传距离为0.068 3~0.136 7。AMOVA分析结果显示遗传分化大部分(68.7%)来自于群体内。其余的遗传分化来自于种群间。长乐群体和非长乐群体(QD, BH, JN, LYG)的遗传分化值(Gst=0.309 0~0.364 9)较高, 而非长乐群体间的Gst较低(0.148 2~0.240 3), 说明长乐群体有明显的遗传分化。本研究发现广西群体与北方群体亲缘关系近, 与福建群体亲缘关系远, 这与一般规律不符。
尤仲杰等[9]用 RAPD标记对我国山东胶南(JN),江苏南通(NT, 即启东), 浙江台州(TZ), 福建福州(FZ, 即长乐), 广西北海(BH)5个自然群体西施舌的遗传差异进行了形态学和RAPD分析, NJ法聚类分析显示北海群体与福州群体聚为一支, 用 UPGMA法聚类显示, 北海群体单独为一支。尤仲杰认为北海群体可能已经形成了地理群体。黎中宝[10]用等位酶对启东、福建漳港、广西北海 3个地理群体的西施舌生化遗传多样性进行了研究, 结果显示, 漳港群体与北海群体聚为一支, 启东群体为单独一支。林昕[15]对启东、福建长乐和深沪湾三个地理群体的西施舌进行了ITS1分析, 发现江苏西施舌与福建西施舌遗传距离较大, 但未形成区别于其他地区西施舌的独特的种。孔令锋等[6-8]用形态学数据、AFLP和16S分子标记对北方的4个地理群体(即墨、胶南、日照和启东)与福建的 2个群体(长乐、漳港)的遗传差异进行了研究, 认为福建的西施舌与北方的 4个群体存在明显的遗传差异, 聚类时明显的分为两支,北方的 4个群体聚为一支, 福建的 2个群体聚为一支。本研究聚类分析发现西施舌 CL群体聚为一支,这与孔令锋的结果一致, 与黎中宝、林晰的结果不完全一致, 但本研究显示西施舌群体间遗传关系与其所处的地理位置远近不完全相符。广西群体(北海)与北方的3个群体(QD, JN, LYG)聚在一起, 这与尤仲杰的结果相反。广西群体与福建群体地理位置近,易发生基因交流, 因此应该与福建群体遗传关系近,或形成特殊的地理群体, 但本研究的结果显示它与北方群体亲缘关系近, 与福建CL群体亲缘关系远。
到目前为止, 我国学者从形态学, 生物化学(同工酶)和分子生物学(RAPD, AFLP, ITS1, 16S rRNA基因)等多个方面对西施舌群体遗传多样性及遗传差异进行了研究, 大多数结果认为福建群体(包括长乐,神户湾、漳州等群体)遗传结构独特, 但广西群体的分类地存在分歧。因此, 广西群体的系统发生地位仍不明朗, 需用其他分子标记作深入的研究。我国西施舌分布于大连、江苏、浙江、福建、广东、广西等沿海地区。但福建长乐的西施舌个体大, 品质好, 也有其独特的分子标记。本研究的RAPD引物中, S299引物扩增出的约 600bp条带为长乐群体所特有, 且稳定清晰, 可作为鉴定长乐群体的分子标记。其他群体无典型的共享RAPD条带。
[1] 孟学平, 高如承, 董志国, 等. 西施舌营养成分分析与评价[J]. 海洋科学, 2007, 31(1): 17-22.
[2] 吴进锋, 陈利雄, 陈素文. 西施舌增养殖的研究现状与展望[J]. 大连水产学院学报, 2005, 20(2): 137- 141.
[3] 孟学平, 程汉良, 董志国. 我国西施舌研究现状及展望[J]. 河北科技师范学院学报, 2005, 19(4): 71-75.
[4] 陈文龙, 刘德经, 许万竹. 西施舌(Mactra antiquataSpengler)人工育苗初步研究[J]. 水产学报, 1966, 3( 2): 130-141.
[5] Liu H, Zhu J X, Sun H L, et al. The clam, Xishi tongueCoelomactra antiquata(Spengler), A promising new candidate for aquaculture in China[J]. Aquaculture,2006, 255(1-4): 402-409.
[6] Kong L F, Li Q, Qiu Z X. Genetic and morphological differentiation in the clamCoelomactra antiquata(Bivalvia: Veneroida) along the coast of China[J].Journal of Experimental Marine Biology and Ecology,2007, 343(1): 110-117.
[7] Kong L F , Li Q. Genetic comparison of cultured and wild stocks of the clamCoelomactraantiquata(Spengler) in China using AFLP markers[J].Aquaculture, 2007, 271(1-4): 152-161.
[8] Kong L F, Li Q. Genetic evidence for the existence of cryptic species in an endangered clamCoelomactra antiquata[J]. Marine Biology, 2009, 15(7): 1 507-1 515.
[9] 尤仲杰, 包永波, 张爱菊. 中国沿海西施舌 5个自然群体形态差异和 RAPD分析[J]. 海洋学报, 2007,29(3): 98-104.
[10] 黎中宝, 王展林, 张桂玲, 等. 西施舌(Coelomactra antiquata)群体遗传多样性与分化的研究[J]. 海洋与湖沼, 2009, 40(30): 302-306.
[11] 高悦勉, 孙洋, 才慧梅.3种扇贝遗传结构的 RAPD分析[J].海洋水产研究, 2007, 28(6): 25-31.
[12] 刘萍, 何玉英, 孙昭宁, 等.中国对虾生长性状相关遗传标记的筛选与克隆[J].海洋水产研究, 2007,28(2): 1-6.
[13] 杨奔, 尤锋, 李军, 等. RAPD 对圆斑星鲽和条斑星鲽养殖群体的遗传多样性研究[J]. 海洋科学, 2009,33(8): 26-29.
[14] Yeh F C, Yang R, Boyle T J, et al. PopGene32,Microsoft Windows-based Freeware for Stock Genetic Analysis. version 1.32[R]. Edmonton, Alberta, Canada:Molecular Biology and Biotechnology Centre,University of Alberta, 2000.
[15] 林昕, 梁君荣, 高亚辉, 等. 3个地区西施舌的ITS-1基因片段序列分析[J]. 生命科学研究, 2008, 12(1):14-19.
Received: Nov., 18, 2009
Key words:Coelomactra antiquata: genetic structure: RAPD
Abstract:The Randomly Amplified Polymorphic DNA (RAPD) technique was applied to assess the genetic variations among five stocks [Changle (CL), Qidong (QD), Lianyungang (LYG), Shandong (JN) and Guangxi (BH)] ofCoelomactra antiquatausing 55 random primers. Genetic diversity was analyzed with PopGen (Version 1.31)software. A total of 12 highly reproducible primers and 88 clear and stable bands were obtained. The lengths of amplified fragments were from 100 bp to 3000 bp. The primer S299, which amplified 600 bp fragment, was specific to the CL stock. The proportion of polymorphic loci was from 62.07% to 78.26% and Shannon index was from 0.210 5 to 0.312 3. Genetic distance among five stocks was from 0.068 3 to 0.223 9. The genetic distance between the CL stock and the other four stocks was 0.182 7~0.223 9, and the genetic distance among the four non-CL stocks was 0.068 3~0.136 7. Coefficient of genetic differentiation among stocks (Fst) was 0.31301 (P<0.05), indicating the genetic variation among stocks accounted for 31.30%. The value of genetic differentiation (Gst) between CL and the other four stocks were 0.364 9, 0.344 8, 0.325 0, and 0.309 0 for QD, BH, JN, and LYG, respectively. The values of genetic differentiation (Gst) among non-CL stocks were 0.148 2~0.240 3. The above data show that the maximum genetic differentiation occurred in the CL stock. Phylogenetic analysis shows that QD and JN stocks formed a clade,which subsequently clustered with BH stock, and then got together with LYG stock. Meanwhile CL stock formed one separate clade.
(本文编辑:张培新)
RAPD analysis of genetic structure and differentiation of Coelomactra antiquata
MENG Xue-ping1, WANG Shuai2, GAO Ru-cheng2, SHEN Xin1, CHENG Han-liang1,TIAN Mei1
(1. Marine School of Huaihai Institute of Technology, Jiangsu Key Laboratory of Marine Biotechnology,Lianyungang 222005, China; 2. College of Life Sciences of Fujian Normal University, Fuzhou 350108, China)
Q951+.3
A
1000-3096(2011)02-0006-04
2009-11-18;
2010-04-13
江苏省自然科学基金项目(BK2007066); 国家 863计划项目(2004AA603140); 江苏省海洋生物技术重点建设实验室开放课题(2005HS009, 2009SH13); 江苏省教育厅自然科学基金项目(05SJD240028)作者简介: 孟学平(1955-), 男, 硕士, 内蒙古商都县人, 教授, 主要从事水生生物生化及分子生物学研究, E-mail: mxp2002@hotmail.com, 电话: 0518-85890806
猜你喜欢
教育评论(2022年8期)2022-09-12
巴蜀史志(2021年2期)2021-09-10
老年教育(老年大学)(2020年3期)2020-06-02
海峡姐妹(2019年9期)2019-10-08
福建基础教育研究(2019年2期)2019-09-10
侨园(2016年8期)2017-01-15
中国卫生(2016年11期)2016-11-12
公民与法治(2016年15期)2016-05-17
红领巾·成长(2016年6期)2016-05-14
中老年健康(2016年1期)2016-03-07
