
滇中蛹虫草居群生物学及其生态学习性研究*
2011-08-08 04:42:54杨钟林陈自宏王元兵
中国食用菌 2011年5期
杨钟林,虞 泓,陈自宏,王元兵
(云南大学中草药生物资源研究所云百草实验室,云南 昆明 650091)
蛹虫草Cordyceps militaris(Fr.)Link,别名蛹草、北虫草、北冬虫夏草,分类学上属真菌界Fungi、子囊菌门Ascomycota、子囊菌纲Ascomycetes、粪壳菌亚纲Sordariomycetidae、肉座菌目 Hypocreales、麦角菌科 Clavicipitaceae、虫草属Cordyceps[1]。虫草属真菌是一类具有重要药用和开发价值的真菌,蛹虫草作为虫草属真菌的模式种,具有和冬虫夏草类似的活性成分和医疗保健功效[2],且人工规模培养已经成功,正逐渐成为冬虫夏草的重要替代品。目前主要集中于蛹虫草培养方法、菌种 “退化”与遗传变异、生物活性物质及生理功能等问题的研究[3]。
蛹虫草分布相对广泛,主要分布在韩国、朝鲜半岛、俄罗斯西伯利亚、我国东北和内蒙等温带区域[5-8],其分布和发生依然有其严格的地域性,并受一定生态地理环境因素制约。蛹虫草生长在次生林地带的阔叶林及其混杂林下,要求林间郁闭度在0.6左右,土质疏松,含水量70%~80%;主要以鳞翅目昆虫的蛹或幼虫为寄主,其次为鞘翅目和双翅目昆虫的幼虫与蛹;子座在6月~9月从地面长出[2,4]。
蛹虫草在云南有分布记载[7-16],在滇中、滇东南、滇西南、滇西、滇西北等地区均有分布。调查发现,蛹虫草通常分布在云南亚热带常绿阔叶林、常绿与落叶阔叶林混交林、常绿次生灌丛、常绿阔叶林与云南松Pinus yunnanensis Franch.混交林、常绿阔叶林与华山松Pinus armandii Franch.。分布于北方温带的蛹虫草,有学者对其生物学和生态学习性做过比较系统的研究[7,8,10,13]。 迄今为止, 关于云南亚热带地区分布的蛹虫草居群,其生物学和生态学习性未见系统的研究报道。通过对滇中地区不同蛹虫草居群进行调查和采样,从形态学和生态习性两方面对滇中地区蛹虫草进行调查研究,旨在为云南蛹虫草资源的理论研究、开发利用以及人工栽培提供基础资料。
1 材料与方法
1.1 试验材料
1.1.1 野生蛹虫草材料
2010年6月~10月调查采集于滇中地区昆明西山、昆明野鸭湖、嵩明大哨和楚雄紫溪山等地的大量新鲜蛹虫草(表 1)。
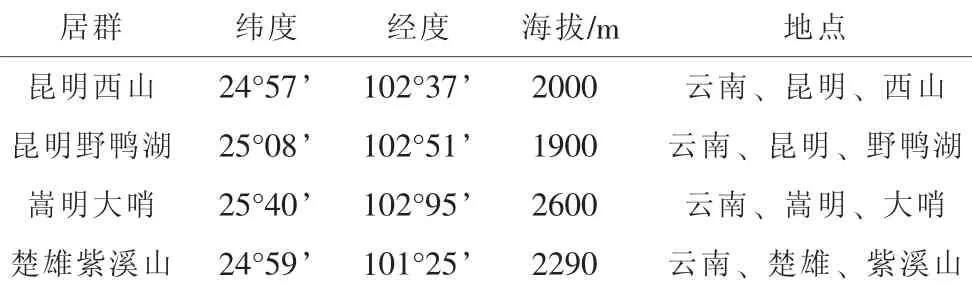
表1 蛹虫草不同居群分布
1.1.2 蛹虫草无性型菌株材料
从野生蛹虫草菌核或子座部位分离得到蛹虫草无性型菌株,编号为CMIL YY 11、CMIL SM 7、CMIL ZXS 9和CMIL XS 13。
1.2 培养基
蛹虫草菌株的分离和培养,使用加1%蛋白胨的PDA培养基。
1.3 蛹虫草有性型的观察
用游标卡尺测量新鲜蛹虫草子座长、粗及可孕部分的长、粗。挑取成熟子囊壳在Olympus CX40显微镜下进行显微观察并照相,用成都励扬精密机电有限公司生产的万能视频成像装置 LY-WN-HP super CCD专业软件Scopephoto,测量子囊壳、子囊、子囊帽和子囊孢子大小。
1.4 蛹虫草无性型的观察
用PDA培养基在凹陷载玻片上培养蛹虫草15 d左右,在Olympus CX40显微镜下进行显微观察并照相,用Scopephoto测量孢梗及分生孢子的大小。
2 结果与分析
2.1 滇中蛹虫草的分布及生态习性
云南省篙明县阿子营大哨居群,位于昆明以北42 km,海拔2600 m。植被是以华山松为主的人工林地,林下空旷,透光、透气好。成土母质为石灰岩、砂岩、页岩等,土壤为红壤。地表平整,无灌草丛,土表覆盖厚约2 cm~4 cm的华山松针叶腐殖层,该腐殖层透气性好,含水量高,有机质含量非常丰富,蛹虫草着生于此层中。由于发生地位于平坦及缓坡上,因而该林具有较好的保水、保肥能力。华山松林缘,分布有密毛蕨Pteridium revolutum(Bl.)Nakai、 光叶石栎 Lithocarpus mairei(Schottk)Rehd.、老鸦炮Vaccinium fragile Franch.、金丝梅Hypericum patulum Thunb.ex Murray、绶草Spiranthes lancea(Thunb.)Baker等植物。
昆明西山居群位于太华寺后山东坡,海拔为2000 m,东面为滇池。主要植被为半湿润常绿阔叶林,包括滇青冈CyclobalanopsisglaucoidesSchott、 高 山 栲 Castanopsis delavayi Franch.、 元 江 栲 Castanopsis orthacantha Franch.、滇石栎 Lithocarpus dealbatus(Hook.f.et Thoms)Rehd.、栓皮栎Quercus varilibilis Bl.等,混生有华山松和云南松,灌木和杂草主要有荩草Arthraxon hispidus(Thunb.)Makio、刚莠竹Microstegium ciliatum(Trin.)A.Camus、云南沿阶草Ophiopogon tienensis Wang et Tang、紫茎泽兰 Eupatorium adenophorum Spreng.、 猪秧秧 Rubia cordifolia Linn.、 鬼 针草Bidens pilosa Linn.、 牛膝菊 Galinsoga parviflora Cav.、 凤尾 蕨 Pteris longigolia Linn.、 密 毛 蕨 P.revolutum(Bl.)Nakai、 梁 王 茶 Nothopanax delavayi(Franch.)Harms ex Diels、老鸦炮Vaccinium fragile Franch.、金丝梅Hypericum patulum Thunb.ex Murray等。西山成土母质为石灰岩和玄武岩,土壤类型为红壤,土表覆盖厚约2 cm~3 cm枯枝层,主要由常绿阔叶树林的枯枝落叶以及腐烂的松针叶所组成,蛹虫草主要着生于此枯枝层中。枯枝层的下面是厚30 cm~50 cm的腐殖质层,此层土壤内有机质含量较高,土质肥沃,结构疏松,通气性良好,含水量高。西山蛹虫草子实体生长期地表温度为 10℃~22℃,土壤 pH值为5.5~6.5。
楚雄紫溪山居群位于紫顶寺南面溪谷南岸,海拔约为2290 m,溪谷东面有一小湖泊。植被类型为半湿润常绿阔叶林和山地次生常绿阔叶林。主要植物有有元江栲Castanopsis orthacantha Franch.、 滇石栎 Lithocarpus dealbatus(Hook.f.et Thoms)Rehd.、高山栲、滇青岗、大果冬青Ilex macrocarpa Oliv、 槲 栎 Quercus aliena Bl、 新 樟Neocinnamomum delavayi(Lec.)Liou,灌木和杂草混生形成灌、草丛。林内光照较弱,散射光线充足。土壤表层为枯枝落叶腐殖层,该层湿润、结构疏松、透气性好。蛹虫草主要着生在灌草丛下的腐殖层中。由于发生地在溪谷中,该地区气候温和,空气湿润。
昆明野鸭湖居群位于野鸭湖西南岸农田田埂,位于山的北坡,海拔为1990 m。原有植被受人为开垦的影响,高大树木仅为不多的干香柏Cupressus duclouxiana Hichel以及农田中的梨树Pyrus pyrifolia(Burm.f.)Nakai、野核桃Juglans cathayensis Dode等果树。田埂及路边长满杂草,主要为蒿类Artemisia spp,伴生有蕨Pteridium aquilinum var.Latiusculum(Desv.)Underw.、 凤 尾 蕨 Pteris multifida Poir、 黄索梅 (黄泡)Rubus pectinellus Maxim.Bull.、 千里光Senecio scandens Buch.-Ham.ex D.Don、 朝天罐 Osbeckia opipara C.Y.Wu et C.Chen、假酸浆Nicandra physaloides(Linn.)Gaertn.、 曼陀罗 Datura stramonium Linn.、 鬼针草Bidenspilosa Linn.、 白 茅 Imperata cylindrica(Linn.)Beauv.,地表长有苔藓Bryophyta。土壤为黄红壤,土层较厚,土质湿润、疏松、透气性好。因空气湿润,阳光充足,一年生杂草生长旺盛,所以地表腐殖质丰富,有机质含量,在草丛下的腐殖质中容易找到蛹虫草 (表2、图1)。
采集蛹虫草的4个居群分布在 N24.59~25.24、E101.24~102.57范围内,其中除嵩明大哨的海拔较高,为2600 m左右,其余居群的海拔接近,在2100 m~2300 m之间。昆明西山和楚雄紫溪山的植被类型相似,为混生华孢子断裂, 形成 (2.30~3.85) μm×(0.72~0.85) μm的次生子囊孢子。

表2 蛹虫草4个居群的生境

图1 蛹虫草4个居群的植被类型及野生子实体
2.2.4 昆明西山居群
子座单生或数个,从寄主昆虫蛹的各处长出,苍黄色、橙黄色至橙红色,不分支,长2 cm~7 cm。可孕部柱状至棒状,长1.2 cm~3.5 cm、粗2.2 mm~5.0 mm。子囊壳之间充满菌丝,致密半埋生,近圆锥形, (610~720)μm× (300~430) μm。 子囊细长, (254~455) μm× (3.0~3.9) μm, 子囊帽半球形, 宽 3.05 μm~3.95 μm、 高 1.85 μm~2.70 μm。 子囊孢子断裂, 形成 (1.95~4.25) μm×(0.72~0.88)μm的次生子囊孢子。蛹虫草4个居群的有性型产孢结构见表3、图2。山松和云南松的常绿阔叶林,林下有灌草丛;嵩明大哨是以华山松为主的人工林;而昆明野鸭湖主要为田埂上的草丛。
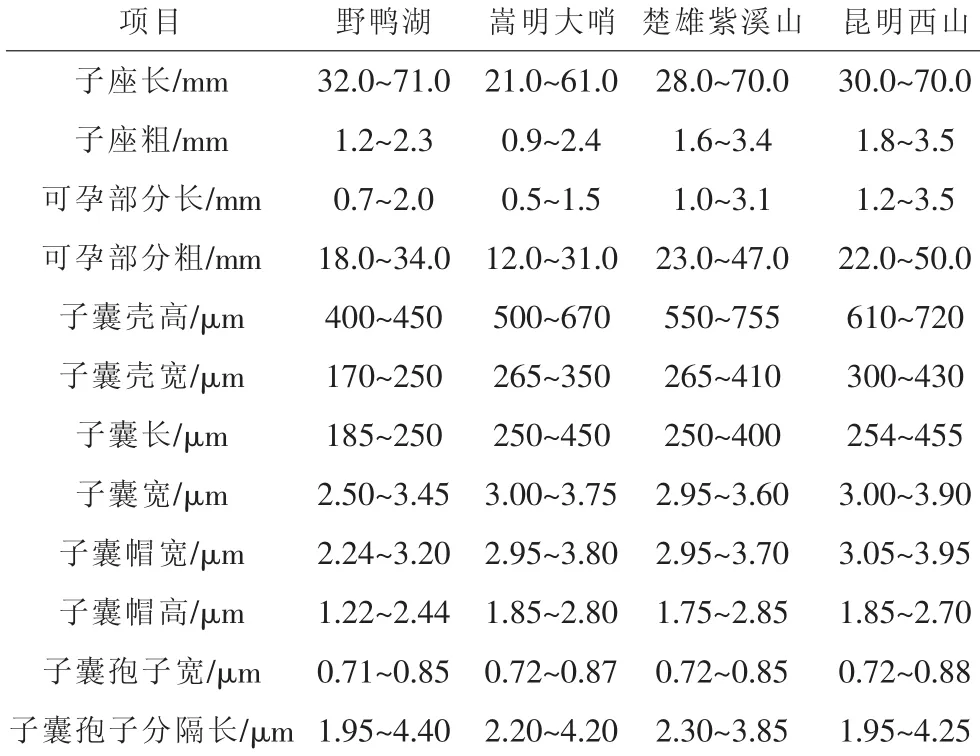
表3 蛹虫草4个居群的有性型产孢结构
2.2 滇中蛹虫草有性型形态变异式样
2.2.1 嵩明大哨居群
子座单生或2个到3个,从寄主昆虫蛹的各处长出,苍黄色至橙黄色,不分支,长2.1 cm~6.1 cm。可孕部柱状至扁平棒状,长0.5 cm~1.5 cm,粗1.2 mm~3.1 mm。子囊壳致密半埋生, 长椭圆形, (500~670)μm× (265~350)μm,大多数子囊壳有乳头状尖端, (155~296)μm×(60~120)μm。子囊细长,子囊孢子缠绕紧密, (250~450)μm×(3.0~3.8) μm。 子囊帽半球形加厚, (2.95~3.80)μm×(1.85~2.80) μm。 子囊孢子断裂, 形成 (2.2~4.2)μm×(0.72~0.87)μm的次生子囊孢子。
2.2.2 昆明野鸭湖居群
子座多数单生,少有2个到3个,从寄主昆虫蛹的各处长出,地上部分橙黄色至橙红色,埋在土中的部分为白色,通常不分支,长3.2 cm~7.1 cm。可孕部柱状至棒状,长0.7 cm~2.0 cm、粗1.8 mm~3.4 mm。子囊壳致密表生,椭圆形, 两头稍尖, (400~450) μm×(170~250) μm。 子囊细长,子囊孢子排列松散, (180~250) μm×(2.50~3.45) μm, 子囊帽半球形加厚, 宽 2.24 μm~3.20 μm, 高1.22 μm~2.44 μm。 子囊孢子断裂, 形成 (1.95~4.40) μm×(0.71~0.85)μm的次生子囊孢子。
2.2.3 楚雄紫溪山居群
子座单生或数个,从寄主昆虫蛹的各处长出,苍黄色至橙黄色,不分支,长2.8 cm~7.0 cm。可孕部柱状至棒状,长1.0 cm~3.1 cm、粗2.3 mm~4.7 mm。子囊壳之间充满菌丝,致密埋生,近圆锥形, (550~755)μm×(265~410) μm。 子囊细长, (250~400) μm× (2.95~3.60) μm,子囊帽半球形, (2.95~3.70)μm× (1.75~2.85)μm。 子囊
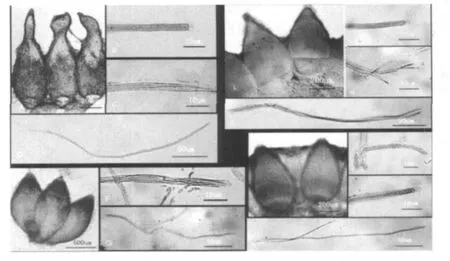
图2 蛹虫草4个居群的有性型产孢结构
4个居群的蛹虫草均为子座单生或数个,从寄主昆虫蛹的各处长出,苍黄色、橙黄色至橙红色,通常不分支,长度相差不大。可孕部位柱状至棒状,其中楚雄紫溪山和昆明西山的比嵩明大哨和昆明野鸭湖的稍长。
在4个居群中,子囊壳的差异最大。野鸭湖的为最小,为椭圆形,两头稍尖,嵩明大哨的比楚雄紫溪山的稍小,致密半埋生,长椭圆形,大多数子囊壳有尖端,楚雄紫溪山的为次大,致密埋生,近圆锥形,昆明西山的为最大,子囊壳之间充满菌丝,致密半埋生,近圆锥形。子囊均为细长形,嵩明大哨、楚雄紫溪山和昆明西山3个居群大小接近,野鸭湖居群的约为其余居群长度的2/3。嵩明大哨的子囊孢子缠绕紧密,其余居群的均松散排列在子囊中。子囊帽均为半球形加厚,除野鸭湖居群的较小,其余居群的基本接近。所采集的这几个居群中,子囊孢子宽及次生子囊孢子长基本相同。
2.3 滇中蛹虫草无性型形态变异式样
所采集的蛹虫草4个居群中,昆明嵩明和野鸭湖居群的蛹虫草无性型以轮枝孢型为主,孢梗为瓶形和细长锥形2种,偶有拟青霉型产孢结构。楚雄紫溪山和昆明西山2个居群的蛹虫草无性型为蛹虫草拟青霉型,其中楚雄紫溪山居群的孢子梗常簇生或轮生于营养菌丝上。蛹虫草4个居群的无性型产孢结构见表4、图3。
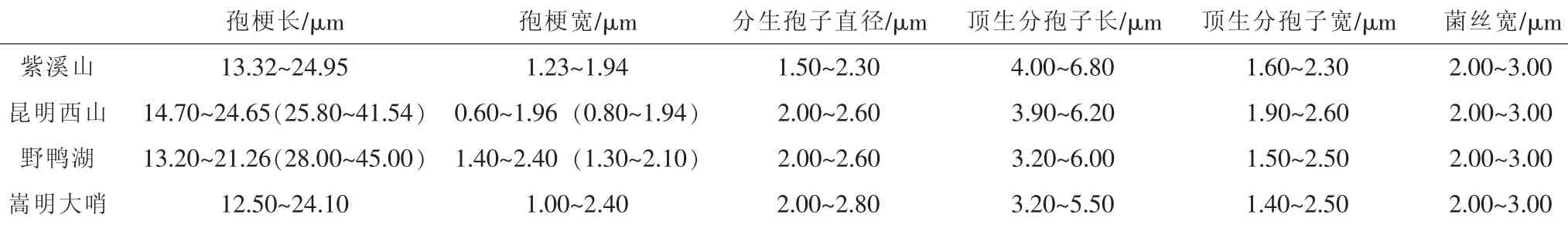
表4 蛹虫草4个居群的无性型产孢结构
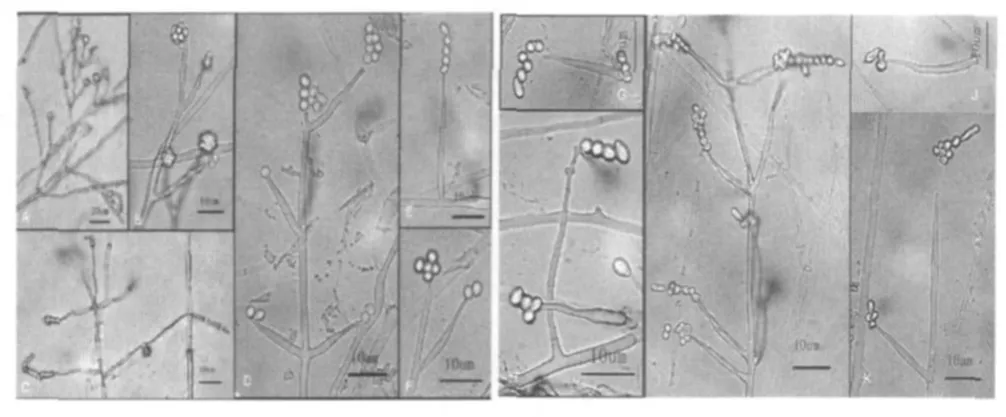
图3 蛹虫草4个居群的无性型产孢结构
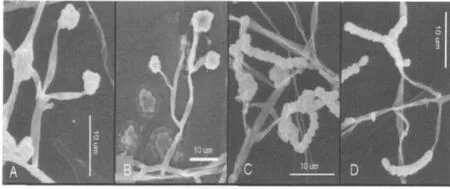
图4 蛹虫草无性型产孢结构电镜照片
嵩明蛹虫草无性型孢子梗簇生或轮生于营养菌丝上,孢子梗基部柱形或膨大,尖端变成细长管状,孢子梗长12.50 μm~24.10 μm、 宽 1.00 μm~2.40 μm。 分生孢子近球形,直径为2.00 μm~2.80 μm,常粘结成头状。其中,也有少数孢子梗为细长锥形, (28.00~45.00)μm×(1.30~2.10)μm,分生孢子为迭瓦状链。
野鸭湖蛹虫草无性型孢子梗簇生或轮生于营养菌丝上,孢子梗一般基部柱形或膨大,尖端变成细长管状,孢子梗为 (13.20~21.26) μm×(1.40~2.40) μm。 分生孢子近球形,直径为2.00 μm~2.60 μm,常粘结成头状。其中,少数孢子梗为细长锥形, (28.00~45.00) μm×(1.30~2.10)μm,分生孢子为迭瓦状链。
昆明西山蛹虫草无性型为拟青霉型 (Paecilomycestype)。孢子梗单生或簇生或轮生于营养菌丝上,孢子梗一般基部柱形或膨大,尖端变成细长管状,约为孢梗长的20%~50%, 孢子梗为 (13.32~24.95) μm× (1.23~1.94)μm, 少数孢子梗为细长的锥形, (25.80~41.54) μm×(0.80~1.94)μm。分生孢子单孢,近球形,直径为2.00 μm~2.60 μm,常连成迭瓦状链,孢子链顶部的分生孢子柱状, (3.90~6.20) μm× (1.90~2.60) μm。
紫溪山蛹虫草无性型大多为拟青霉型 (Paecilomycestype), 菌丝基部偶有轮枝孢型 (Verticillium-type)。 孢子梗单生或簇生或轮生于营养菌丝上,孢子梗一般基部柱形或膨大,近中部或以上向上突然变成细长管状,孢子梗长13.32 μm~24.95 μm, 膨大部宽 1.23 μm~1.94 μm。 分生孢子单孢,近球形,直径为1.50 μm~2.30 μm。分生孢子常连成迭瓦状链,在孢子链顶部的分生孢子柱状, (4.00~6.80) μm× (1.60~2.30) μm。
3 讨论
蛹虫草的发生,是虫草真菌感染寄主昆虫后,在各种环境因素相互作用下形成的复合体,最终形成子实体。子实体的发生除了与虫草真菌本身生理生化因素密切相关外,还与外界环境因素综合作用有很大关系[9]。影响蛹虫草发生的主要生态因素有温度、湿度、植被、寄主昆虫、土壤及地形地貌。蛹虫草子实体发生于每年夏末至秋季,此时云南气候较为温凉,气温为15℃~25℃,有利于子实体和子囊孢子形成。此时正是云南雨季,空气中相对湿度大,有利于蛹虫草子实体迅速生长,同时,子囊孢子借助于雨水和气流散落传播。
调查发现,在滇中地区蛹虫草常发现于半湿润常绿阔叶林、华山松林、云南松林、次生常绿阔叶林灌草丛等植物群落类型中。分布有蛹虫草的云南松林中混生有栎类(Castanopsis spp.、 Cyclobalanopsis spp.、 Lithocarpus spp.、Quercus spp.)和滇油杉Keteleeria evelyniana Mast.等植物。不同植物群落下有不同寄主昆虫分布,植被物种丰富多样的生境,寄主昆虫种类也较多,鳞翅目母蛾科、刺蛾科、枯叶蛾科、尺蛾科、天蛾科等都可以成为蛹虫草真菌的主要寄主昆虫。滇中地区山地植被保存完好,地表腐殖质丰富,且土质疏松,通风透气,排水性好,偏酸性。有一定阳光照射,海拔在1500 m~2500 m的向阳缓坡,较利于蛹虫草生长。
通过比较研究发现,滇中亚热带生长的蛹虫草变异式样较大,其子囊壳着生方式、大小、子囊大小随生境不同而发生变异。在常绿阔叶林下生长的蛹虫草子囊壳较大,成圆锥形,底部宽大,例如昆明西山居群是 (610~720)μm×(300~430) μm; 楚雄紫溪山居群是 (550~755) μm×(265~410)μm。蛹虫草分布生境不一样,蛹虫草子囊壳形态及其着生方式不一样,例如海拔为2600 m的嵩明大哨居群的子囊壳顶端有乳头状的突起,而海拔2000 m的昆明西山居群的子囊壳之间充满菌丝,致密半埋生;在海拔1900 m生长的昆明野鸭湖居群子囊壳较小,椭圆形,两头稍尖。蛹虫草分布生境不一样,蛹虫草成熟子囊大小也不一样,例如嵩明大哨、楚雄紫溪山和昆明西山3个居群大小接近, 昆明西山 (254~455) μm×(3.0~3.9) μm,楚雄紫溪山 (250~400) μm×(2.95~3.60) μm, 嵩明大哨(250~450) μm×(3.0~3.8)μm,野鸭湖居群的约为其余居群长度的2/3,且子囊的宽度均较窄,为 (180~250)μm×(2.50~3.45)μm。蛹虫草分布生境不一样,蛹虫草成熟子囊孢子排列方式也不一样,例如嵩明大哨居群的子囊孢子紧密缠绕排列于子囊中,而其余居群的均松散排列在子囊中。
梁月等[17]通过对蛹虫草大量有性孢子—子囊孢子的分离和培养性状的观察发现,不同来源的子囊孢子后代群体,不但群体之间在培养性状上有不同程度的差异,即使是来自同一子实体的不同子囊孢子菌株间在菌落颜色、质地、生长速率等性状上也存在一定差异,各自产生子实体的能力也不尽相同。
在滇中地区,从蛹虫草分离出的菌株,其无性型产孢结构表现出不同生境下拟青霉型和轮枝孢型所占的优势不同。从常绿阔叶林下分离到的菌株,其无性型以拟青霉型产孢结构占优势,在华山松林和田埂灌草丛下分离到的菌株,其无性型以轮枝孢型产孢结构占优势。本研究从某种程度上支持了高新华等[18]的研究,具有子实体形成能力的野生蛹虫草菌株,同时具有拟青霉型和轮枝型2种产孢结构。只有1种产孢结构的菌株一般难以形成子实体。对于蛹虫草无性型菌种的分类地位,Kobayasi最早正式提出为蛹草头孢霉Cephalosporim militaris,随后有不同学者对其分类地位提出过不同的看法。梁宗琦研究发现此菌有多型现象,在分生孢子聚集方式上存在头状的轮枝孢型和链状拟青霉型2种产孢方式,头状的轮枝孢型为角变株,野生型以拟青霉型为主,定名为蛹草拟青霉Paecilomyces militaris Liang[19]。 Zare 等[20]和 Gams等[21]根据核糖体 rDNA ITS序列以及大亚基和小亚基序列分析,将轮枝孢属中的所有虫生种都移入新属蚧霉属Lecanicillium中,此属的形态特征是分生孢子常排列呈单叠瓦状,瓶梗的形状及瓶梗在分生孢子梗上的着生方式则介于拟青霉和轮枝孢2个属之间。李春如[22]采用了上述分类系统,将蛹草拟青霉转属组合为蛹草蚧霉Lecanicillium militaris(Liang)Li,Fan&Li。梁宗琦[19]现研究发现,蛹草拟青霉在不同培养基上,也能产生拟青霉型和轮枝孢型两型产孢结构。拟青霉型的产孢结构在花生培养基上最多,其次是查氏培养基和萨氏培养基;而在马铃薯培养基上则以轮枝孢型为主。此外,两型产孢结构的比例与菌丝的发育阶段相关。拟青霉型的产孢结构多从基质菌丝上产生,菌落中心比边缘产生的数量大,成熟菌丝比幼嫩菌丝上产生的多。
[1]Kirk PM,Cannon PF,David JC,et al.Ainsworth&Bisby’s dictionary of the fungi[M].Walling:CABI Publishing,2001.
[2]梁宗琦.中国真菌志 (第32卷):虫草属[M].北京:科学出版社,2007.
[3]韩燕峰,梁建东,杜文,等.蛹虫草几个问题的最新研究进展[J].微生物学通报,2009,36(9):1423-1428.
[4]蒲蛰龙,李增智.昆虫真菌学[M].合肥:安徽科学技术出版社,1996.
[5]Sung JM,Choi YS,Kim YO,et al.Cordyceps species collected by Koren entomopathhogenic fungal collection[C]//Proceedings of the third Koren-China joint symposium for mycology(The Koran society of mycology and Chinese academy of sciences).Korea:The koran Society of Mycology,1997:49-60.
[6]Lee TS,Yoon KH.The Index of Koran-Japan mushroom nanes in Korea[M].Korea:Korea Forest Resesrth Institute,2002:1-176.
[7]毕志树,郑国杨,李泰辉.广东大型真菌志[M].广州:广东科技出版社,1994.
[8]袁明生,孙佩琼.四川覃菌[M].成都:四川科学技术出版社,1995.
[9]柴建萍,日兴荣,谢道燕.云南蛹虫草分布及发生生态因素浅析[J].云南农业科技,2002,4(19):25.
[10]邓叔群.中国的真菌[M].北京:科学出版社,1963.
[11]戴芳澜.中国真菌总汇[M].北京:科学出版社,1979.
[12]梁佩琼,陆大京.中国虫草属真菌已知种类资源[J].食用菌,1988(2): 1-4.
[13]邵力平,项存涕.中国森林蘑菇[M].哈尔滨:东北林业大学出版社,1997.
[14]Zhuang Wenying Y.Higher fungi of tropical China[M].Ithaca:Mycotaxon LTD,2001.
[15]朱国胜,刘作易,吴兴亮,等.中国滇黔桂喀斯特地区部分虫草属真菌研究[J].贵州科学,2004,22(1):27-30.
[16]Zang Mu,Kinjo N.Notes on the alpine Cordyceps of China and nearby nations[J].Mycotaxon,1998(66):215-229.
[17]梁月,张国珍,安沫平,等.蛹虫草子囊孢子萌发及其后代群体培养性状观察[J].菌物学报,2005,24(4):525-532.
[18]高新华,吴畏,钱国琛,等.北冬虫夏草单孢菌株配对后对子实体形成的影响与无性型产孢结构关系[J].上海农业学报,2000,16(增刊): 85-92.
[19]梁宗琦, Roland TV Fox.蛹虫草 (Cordyceps militaris) 无性型的多型现象[J].菌物系统,1998,17(1): 57-62.
[20]Zare R,Gains W,Culham A.A revision of verticillium sect.Prostrata I.Phylogenetic studies using ITS sequences[J].Nova Hedwigia,2000(71):465-480.
[21]Gams W,Zare R.A revision of verticillium sect.Prostrata III.Generic classification[J].Nova Hedwigia,2001(72):329-337.
[22]李春如,黄勃,南圣姬,等.具抗癌活性的RCEF0718虫草菌株的鉴定研究[J].莱阳农学院学报:自然科学版,2006,23(4):263-267.
猜你喜欢
食品工业科技(2023年22期)2023-11-20 07:45:10
食品与生物技术学报(2021年9期)2021-09-28 08:07:50
乡村科技(2020年6期)2020-07-09 23:01:41
乡村科技(2020年7期)2020-05-19 09:48:38
中国森林病虫(2020年2期)2020-03-31 11:49:50
遗传(2019年11期)2019-11-28 12:03:42
安徽农业科学(2018年18期)2018-05-14 08:59:48
江苏农业科学(2016年4期)2016-06-14 15:20:09
化学工业与工程(2016年6期)2016-04-10 01:17:11
华南农业大学学报(2015年5期)2015-12-04 03:04:38
