
青海扁茎早熟禾冬眠无性系种群年龄结构特征
2011-04-25 09:44:22周青平刘文辉颜红波雷生春梁国玲
草业科学 2011年11期
伍 磊,周青平,刘文辉,颜红波,雷生春,梁国玲
(1.青海省畜牧兽医科学院草原所,青海 西宁 810016;2.青海大学,青海 西宁810013)
种群是物种在自然界中存在的基本单位,是研究群落与生态系统结构与功能的基础。20世纪70年代初,Harper和White[1]提出了植物种群的构件结构理论,使植物种群生态学迈入新的台阶。国内对植物年龄结构和冬眠芽库进行了大量研究[2-10],主要研究地点集中在中国东北松嫩平原,研究对象包括朝鲜碱茅(Puccinelliachinampoensis)、野大麦(Hordeumbrevisubulatum)、羊草(Leymuschinensis)、冰草(Agropyronmichnoi)、赖草(L.secalinus)、拂子茅(Calamagrostispseudophragmites)和苔草(Carexkirganica)等植物,得出了环境对植物年龄结构和芽库的影响,并推测了种群的发展动态。国内现缺少青藏高原等地植物年龄结构和芽库特征的研究。
青海扁茎早熟禾(Poapratensisvar.ancepscv.Qinghai) 为多年生根茎疏丛型禾草,广泛分布于青藏高原海拔3 000 m以上的天然草地上,为优良栽培牧草品种。其植株耐寒、耐旱、耐刈割,根茎繁殖能力强,须根发达,为家畜喜食的优良牧草。青海扁茎早熟禾能依靠营养繁殖的子代补充更新,这些子代与亲株相连接,能独立生存,是典型的无性系植物。现已对青海扁茎早熟禾种群地上生物量积累动态、地下生物量垂直动态和栽培特征做了一定的研究[11-17],但有关青海扁茎早熟禾根茎繁殖构建龄级特征尚无报道。
通过对青海扁茎早熟禾1年到6年栽培草地冬眠构件及其年龄的统计分析,得出不同生长年限青海扁茎早熟禾冬眠构件的年龄结构和各种构件的龄级结构特征,揭示其冬眠芽库和苗库的形成规律及种群的发展动态和策略,为其在生态环境治理、草地畜牧业生产等方面的利用提供基础资料。
1 材料与方法
1.1试验地概况 试验地设在青海省畜牧兽医科学院草原所试验站,位于西宁城北区,地理坐标36°43′ N,101°45′ E,海拔2 295.2 m,年均温度5.5 ℃,最冷月1月平均温度-7.9 ℃,最热月7月平均温度16.8 ℃,年均降水量402.3 mm,年蒸发量1 310.8 mm,年平均相对湿度56%,全年日照时间为2 618.3 h,无霜期150 d,≥0 ℃年积温2 723.2 ℃·d,≥10 ℃年积温2 230.5 ℃·d。土壤为灌溉栗钙土[12,16-17]。
1.2研究方法 供试材料为青海扁茎早熟禾,由青海省畜牧兽医科学院草原所提供,青海扁茎早熟禾均为每年的成熟期收获的种子。
青海扁茎早熟禾于2005年6月、2007年6月和2009年6月条播,小区面积10 m2(2 m×5 m),各小区种植4行,共20 m样段。播量24 kg/hm2,行距50 cm,播时施225 kg/hm2磷酸二铵。种子成熟后地上部分刈割。
于2009年10月和2010年10月取样。2005年播种的样为5年(5 a)和6年(6 a),2007年播种的样为3年(3 a)和4年(4 a),2009年播种的样为1年(1 a)和2年(2 a)。每个小区内取5个大小为25 cm×25 cm样方。把带回的样洗净根部泥土,逐一计数每个无性系分蘖丛的亲株、冬眠苗和冬眠芽龄级。拔节的亲株作为亲株;刚破土变绿的营养苗和未拔节的亲株,一并作为冬眠苗;从刚萌生到尚未破土的全部芽一并作为冬眠芽。龄级划分方法按根茎禾草无性系种群年龄结构的研究方法[18]进行。
1.3数据分析 用Excel对数据进行初步处理,用SPSS 16.0对各构件年龄结构进行方差分析和曲线拟合。
2 结果与分析
2.1不同生长年限冬眠芽构件的年龄结构 芽在不宜于植物生长的环境下将保持休眠,生长缓慢;在适宜的环境下将生长成为新株。冬眠芽的特征能反映种群的活力。各年份的冬眠芽最高有5个龄级,最少有4个龄级。1 a的芽库由4个龄级组成,芽库中以0龄级的数量最大,占整个芽库的64%,并且随着龄级的增加迅速减少,到最高3龄级仅占3%。2 a的芽库由5个龄级组成,芽库中以0龄级的数量最大,占整个芽库的71%,并且随着龄级的增加迅速减少,到最高4龄级仅占1%。3 a的芽库由5个龄级组成,芽库中以0龄级的数量最大,占整个芽库的75%。4 a的芽库由4个龄级组成,芽库中以0龄级的数量最大,占整个芽库的71%。5 a的芽库由4个龄级组成,芽库中以0龄级的数量最大,占整个芽库的72%。6 a的芽库由4个龄级组成,芽库中以0龄级的数量最大,占整个芽库的72%。各生长年限年龄结构均以0龄数量最高,数量逐龄递减。
0龄冬眠芽数量的排序为2 a>1 a>3 a>4 a>5 a>6 a,1 a到4 a数量相差不大,4 a到5 a数量骤减,5 a数量不到4 a的1/3,5 a和6 a数量相近;1龄冬眠芽数量的排序为1 a>2 a>4 a>3 a>5 a>6 a,2 a、3 a和4 a数量相差很小,5 a和6 a数量相近;2龄冬眠芽数量的排序为1 a>2 a>4 a>3 a>6 a>5 a;3龄冬眠芽数量的排序为2 a>1 a>3 a>4 a>6 a>5 a;4龄冬眠芽数量的排序为2 a>3 a(表1)。
2.2不同生长年限冬眠苗构件的年龄结构 冬眠苗是植物越冬保存于地下的植物组织,由冬眠芽发育而来,将来成长成为植株。1 a苗库由3个龄级组成,苗库中以1龄级的数量最大,占整个苗库的80%,随着龄级的增加而依次减少的趋势,到3龄级仅占5%。2 a苗库由4个龄级组成,苗库中以1龄级的数量最大,占整个苗库的83%, 4龄级仅占1%。3 a苗库由4个龄级组成,苗库中以1龄级的数量最大,占整个苗库的82%,至4龄级仅占1%。4 a苗库由4个龄级组成,苗库中以1龄级的数量最大,占整个苗库的75%,至4龄级仅占1%。5 a苗库由3个龄级组成,苗库中以1龄级的数量最大,占整个苗库的81%。6 a苗库由3个龄级组成,苗库中以1龄级的数量最大,占整个苗库的72%。各生长年限年龄结构均以1龄最高,数量逐龄递减(表1)。
1龄冬眠苗数量的排序为1 a>2 a>3 a>4 a>5 a>6 a,2 a、3 a和4 a数量相差很小,4 a和5 a数量相差很大,5 a和6 a数量相近;2龄冬眠苗数量的排序为1 a>4 a>3 a>2 a>6 a>5 a;3龄冬眠苗数量的排序为1 a>4 a>2 a>3 a>6 a>5 a;4龄冬眠苗数量的排序为2 a>4 a>3 a。
2.3不同生长年限亲株构件的年龄结构 1 a亲株由2个龄级组成,1龄平均155.5株,2龄平均31.33株。2 a亲株由3个龄级组成,1龄数量最大,逐龄递减。3 a亲株由4个龄级组成,1龄数量最大,占75%,4龄占2%。4 a亲株由4个龄级组成,1龄数量最大,占65%,4龄占3%。5 a亲株由3个龄级组成,1龄数量最大,占70%。6 a亲株由3个龄级组成,1龄数量最大,占65%。随着年限增长,亲株龄级呈先增后减趋势(表1)。各生长年限年龄结构均以1龄最高,数量逐龄递减。
1龄亲株数量的排序为3 a>4 a>2 a>5 a>6 a>1 a,3 a和4 a数量相差很小,1 a和6 a数量相近;2龄亲株数量的排序为4 a>3 a>2 a>5 a>6 a>1 a;3龄亲株数量的排序为4 a>3 a>2 a>6 a>5 a;4龄亲株数量的排序为4 a>3 a。
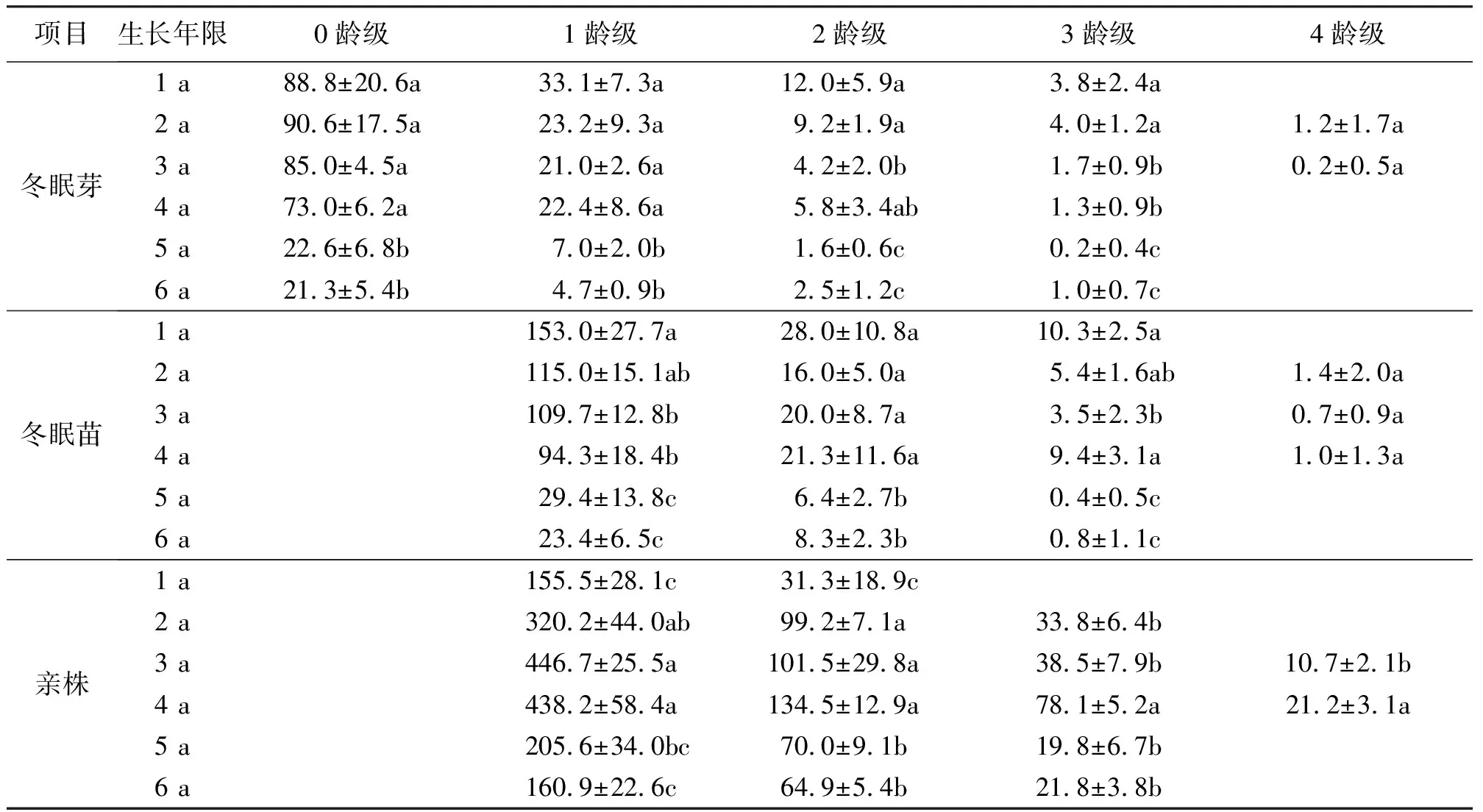
表1 冬眠芽、冬眠苗构件和亲株不同生长年限的年龄结构 株
2.4冬眠构件的时序划分 把各冬眠构件数量按生长年限汇总,并对各年限间数量进行方差分析,以期把各构件不同年限归类,判断生长的时序阶段。
亲株数量4 a最高,为672.3±37.74株,5 a和6 a较低,二者差异不显著,但显著高于最低的1 a。冬眠苗1 a与各年份间的差异均极显著且数量最高,2 a、3 a和4 a间无显著差异,5 a与6 a最低,二者无显著差异。冬眠芽1 a、2 a、3 a和4 a差异不显著,显著高于5 a与6 a,1 a数量最高,为137.67±28.97株(表2)。

表2 冬眠构件数量特征
青海扁茎早熟禾各冬眠构件的时序归类:亲株1 a、2 a、3 a和4 a各为一类,5 a和6 a为一类;冬眠苗1 a为一类,2 a、3 a和4 a为一类,5 a和6 a为一类;冬眠芽1 a、2 a、3 a和4 a为一类,5 a和6 a为一类。综合各冬眠构件的时序分类,冬眠构件数量特征时序可分为3个时期:1 a为一个时期,2 a、3 a和4 a为一个时期,5 a和6 a为一个时期。
2.5冬眠构件年龄结构曲线拟合 对于种群,预测其发展趋势是种群研究的一项重要工作。各生长年限的构件均以幼龄的比例高而高龄的比例低。幼龄和高龄之间的比例可以体现出种群的发展趋势,即幼龄比例高越有可能是增长型的龄级结构。仅用方差分析得出冬眠构件的时序,但无法预测种群将来的发展趋势。以龄级为自变量,各构件的数量特征为因变量进行曲线拟合,拟预测种群发展趋势,得出冬眠苗没有合适的拟合曲线;冬眠芽最合适的是逆变换曲线;亲株最符合生长曲线。
冬眠苗符合直线模型,其表达式为S=-35.96t+221.3(F=57.7>F0.01=15.52);冬眠芽最符合的是生长曲线模型,其表达式为S=e5.472-0.374t(F=68.7>F0.01=15.52);亲株的数量生长最符合二次曲线模型,其表达式为S=-86.417t2+547.455t-279.752 (F=136>F0.01=15.52)。式中t为生长季,S为数量。
从曲线拟合中得出:当冬眠苗和冬眠芽的各龄级数量的曲线的乘幂曲线F值越高,就越有可能是增长型龄级结构。从而1 a和2 a为增长龄级结构,3 a和4 a为稳定龄级结构,5 a和6 a为衰退龄级结构。
3 结论
青海扁茎早熟禾各构件最高龄级均为4龄,大于4龄的构件很罕见。一个亲株在环境胁迫和种内竞争的压力下最多能经历4个生长年限,大于4个生长年限的很罕见。虽然一个亲株可以产生多个克隆体,但就一个根颈的潜力来说,只能经历4个生长年限,甚至更短。这就需要不断有新的空间产生来容纳新植物的生长。
青海扁茎早熟禾第1年种子入土发芽后能长出2龄的亲株。1 a和2 a阶段,地面有充足的空间,种群密度低,营养繁殖旺盛,种群呈增长结构;3 a和4 a数量仍然在增长,但由于空间不足,密度过高,种群处于稳定结构,此时营养繁殖并不旺盛,植株主要进行有性生殖。由于无多余空间以及种群密度过高,5 a和6 a出现衰退现象,为渐衰退期。
对于多年生草本植物种群发展趋势的研究可以从是否符合某个曲线、符合程度(如F值大小)以及系数来考虑。本研究就从曲线拟合中得出:当冬眠苗和冬眠芽的各龄级数量的曲线的乘幂曲线F值越高,就越有可能是增长型龄级结构。
[1]Harper J L,White J.The demography of plants [J].Annual Review of Ecology and Systematics, 1974,5:419-463.
[2]李海燕,杨允菲.松嫩平原水淹恢复演替过程中羊草无性系种群构件的年龄结构[J].生态学报,2004,24(10):2171-2177.
[3]董云波,焦德志,李波,等.扎龙自然保护区羊草种群亲株的数量性状[J].安徽农业科学,2007,35(19):5732-5733.
[4]杨允菲,郑慧莹,李建东.放牧干扰对根茎冰草无性系种群年龄结构的影响[J].植物生态学报,2001,25(1):71-75.
[5]杨允菲,魏春雁,张宝田,等.松嫩平原碱化草甸旱地生境芦苇种群的芽流和芽库动态[J].应用生态学报,2005,16(5):854-858.
[6]焦德志,杨允菲.扎龙自然保护区不同土壤生境羊草种群冬眠特性研究[J].安徽农业科学,2006,34(3):479-480.
[7]杨允菲,张宝田,李建东.松嫩平原人工草地野大麦无性系冬眠构件的结构及形成规律[J].生态学报,2004,24(2):268-273.
[8]杨允菲,张宝田.松嫩平原贝加尔针茅无性系构件的结构及生长规律[J].应用生态学报,2005,16(12):2311-2315.
[9]张学勇,杨允菲,张殿顺,等.辽东半岛结缕草无性系种群构件生物量相关模型及形成规律[J].草原与草坪,2006(3):23-26.
[10]杨允菲,李建东.松嫩平原人工草地羊草和野大麦叶种群的趋同生长格局[J].草业学报,2003,12(5):38-43.
[11]周青平,颜红波,韩志林,等.原根茎型优质草种“青海扁茎早熟禾”的驯化选育[J].草地学报,2008,16(4):328-335.
[12]杨艳莉,周青平,颜红波.行距对青海扁茎早熟禾无性繁殖影响的研究[J].草业科学,2009,26(5):66-71.
[13]刘文辉,梁国玲,周青平.青海扁茎早熟禾种子成熟过程中落粒性与生长生理特性的研究[J].种子,2009,28(6):18-24.
[14]刘文辉,周青平,颜红波.青海扁茎早熟禾种子生长生理特性研究[J].草业科学,2007,24(5):69-73.
[15]石红霄,周青平,颜红波,等.三种根茎型禾草无性繁殖时空扩展初探[J].草业科学,2008,25(5):131-136.
[16]刘文辉,周青平,颜红波,等.青海扁茎早熟禾种群地上生物量积累动态[J].草业学报,2009,18(2):18-24.
[17]刘文辉,颜红波,周青平.青海扁茎早熟禾地下生物量垂直动态[J].西北农业学报,2009,18(4):233-239.
[18]杨允菲,郑慧莹,李建东.根茎禾草无性系种群年龄结构的研究方法[J].东北师大学报(自然科学版),1998(1):49-53.
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11 03:44:34
数学物理学报(2021年6期)2021-12-21 06:24:52
河南农业大学学报(2020年2期)2020-05-22 02:56:30
党的生活·青海(2019年12期)2019-12-23 08:28:56
数学物理学报(2019年5期)2019-11-29 07:47:04
山西林业科技(2019年1期)2019-05-27 12:24:52
岷峨诗稿(2017年4期)2017-04-20 06:26:36
中国卫生(2016年8期)2016-11-12 13:27:04
数学年刊A辑(中文版)(2016年1期)2016-10-30 01:46:20
海洋渔业(2016年6期)2016-04-16 03:00:35
