
大针茅根系构型对草地退化的响应
2011-04-25 10:07:50周艳松王立群梁金华王旭峰
草业科学 2011年11期
周艳松,王立群,张 鹏,梁金华,王旭峰
(内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010018)
大针茅(Stipagrandis)是亚洲中部草原亚区特有的蒙古草原种,在我国分布较广,是划分草原植被的指示植物[1]。大针茅为多年生密丛型旱生草本植物,是良好饲用牧草[2],是典型草原区重要的畜牧业资源且在维系草原生态平衡方面发挥着重要的作用。但近年来,由于自然原因加之对草地不合理的利用方式,内蒙古自治区60%可利用草地出现不同程度的退化[3],关于以大针茅为建群种的草地退化的防治已经引起国内许多学者的重视,并开展了一些研究。安渊等[4]研究了大针茅草原不同退化阶段的植物和土壤化学性状的差异,王炜等[5]从羊草(Leymuschinensis)+大针茅草原退化机理方面进行了研究,张红梅等[6]从不同放牧条件下大针茅的种群形态变异方面进行了研究,宗振宏[7]还从抗旱性角度对大针茅的根系分布进行了分析,但关于大针茅根系构型在不同退化程度草地中的变化还未见报道。大针茅根系构型[8-10]为密丛型[11],其根系构型在不同退化程度的草地中的变化情况能直接反映土壤物理性状、土壤水分和家畜采食、践踏等综合环境因素对大针茅个体及种群的影响。因此,本研究分析大针茅根系构型在不同退化程度的草地中的变化情况,旨在为大针茅在草地植被中群落演替和防治草地沙化提供理论依据,也为草地植物根系构型的研究提供借鉴。
1 研究区概况
研究区位于内蒙古典型草原区的中国科学院内蒙古草原生态系统定位研究站退化样地中进行。其地理坐标为43°37′~43°38′ N,116°39′~116°41′ E,平均海拔1 182 m,地势平坦。属干旱半干旱草原气候,夏季温暖湿润,冬季寒冷干燥,年均气温-0.4 ℃,1月平均气温-23 ℃,7月平均气温17.9 ℃,年均降水量350 mm,主要集中于6-8月。土壤为沙质栗钙土和栗钙土。研究区内,气候顶级群落的建群种为大针茅,在过度放牧干扰下,则出现分别以冷蒿(Artemisiafrigida)和星毛委陵菜(Potentillaacaulis)为建群种的偏途顶级群落[12]。
2 材料与方法
2.1退化草地的划分 根据植被盖度[13]、地上生物量[14]、物种组成和放牧率4个指标确定草地退化梯度系列,并在各退化草地中典型地段设置样地。梯度包括:轻度退化草地(大针茅+羊草+冷蒿群落)、中度退化草地[大针茅+冰草(Agropyroncirstatum)+糙隐子草(Cleistogenessquarrosa)群落]、重度退化草地(冷蒿+大针茅+糙隐子草群落)和极度退化草地(星毛委陵菜+冷蒿+糙隐子草群落),分别用L、M、H和E表示。
2.2取样 于2010年7月17-23日开始取样。在每个样地中选择中等冠幅的大针茅5个株丛;采用壕沟法在距株丛中心40 cm处挖深70 cm、宽70 cm的土壤剖面,配合使用软毛刷、硬毛刷和喷水壶,尽最大的可能取到相对完整的根系。在挖根过程中,仔细观察根系在土壤剖面上的整体构型,标记根系集中分布的位置,并量取根幅、根深和着生于分蘖节处并垂向生长的不定根与水平方向的夹角,对不定根和当年生地上枝条记数,并测定整个株丛地表的分蘖节宽度,现场作图、拍照。将挖取的植物根系,低温保存(以防止根系因失水而收缩变脆),带回室内后清洗干净,用WinRHIZO 2009根系分析系统扫描提取数据。所获根系构型参数包括:总根长、根表面积、根总体积、根平均直径、根尖数、根分叉数、根平均分支角度、一级垂向根条数、一级垂向根长度、一级侧根数、一级侧根长度、二级侧根数、二级侧根长度和不定根条数。分别把测试后的根系按单株分开装袋,烘干后测其干质量。
2.3数据处理 应用Excel 2003和PASW Statistics 18.0进行数据处理,并做相应的单因素方差分析和主成分分析。
3 结果与分析
3.1草地退化对大针茅根系构型相关参数及地下生物量的影响 从表1可知,在L草地中,大针茅的根深和株丛分蘖节宽度显著高于其他退化程度的草地(P<0.05),其中根深较E的长5.2 cm,株丛分蘖节宽度较E的宽1.8 cm。从L、H到E退化系列,各退化程度的草地中大针茅根幅、根深、不定根条数、株丛分蘖节宽度和地下生物量都有减小的趋势。在M草地中,根幅大针茅的根深、不定根条数、株丛分蘖节宽度和地下生物量最小。这直接导致了大针茅根系在土壤中的分布范围减小,降低了根系对土壤资源的竞争能力,使之在群落中的竞争优势下降,出现“植物个体小型化现象”[5]。总体上分析,在借鉴“植物个体小型化”概念的基础上,随草地退化程度的加剧,可以发现大针茅的根系构型同样有小型化的趋势,但这种趋势并非直线下降,在M草地中根系构型小型化尤为突出,甚至超过了H和E草地。

表1 各退化梯度中大针茅根系构型相关参数比较
3.2大针茅根系构型参数的主要因子分析 本研究选定了14个根系构型参数(总根长、根表面积、根总体积、根平均直径、根尖数、根分叉数、根平均分支角度、一级垂向根条数、一级垂向根长度、一级侧根数、一级侧根长度、二级侧根数、二级侧根长度和不定根条数)作为分析大针茅根系构型在不同退化程度的草地中发生变化的因子。但对这14个参数都进行对比分析难免会导致主次混淆不能突出最重要的影响因子的问题产生,因此需对这14个根系构型参数做主成分分析,以发现能表明不同退化程度的草地中根系构型变化显著的因子,简化分析,并得到各成分特征值碎石图(图1)。通过主成分分析(表2),最终筛选出根分叉数、根表面积和总根长3个根系构型参数作为分析大针茅根系构型在不同退化程度的草地中变化情况的主要因子。这3个参数的特征值分别为8.8、2.7和1.2,依次减小,且值均大于1,其中根分叉数解释了根系构型总体变化的62.8%,解释力最强。这3个根系构型参数的累积贡献率为90.7%,即根分叉数、根表面积和总根长表征了不同退化程度的草地中大针茅根系构型变化的90.7%,可作为分析该变化的参数。
3.3不同退化草地中根分叉数、根表面积和总根长的变化 在主成分分析的基础上对这3个参数进行方差分析(表3)。L草地中大针茅的根分叉数、根表面积和总根长均显著高于其他3个退化程度的草地(P<0.05)。从L、H到E草地,根分叉数、根表面积和总根长3个根系构型参数均有减小的趋势。随草地退化程度的加剧,大针茅根系对土壤资源的利用能力及对土壤环境的适应能力降低,一定程度上影响地上生物量(表3),L草地中的地上生物量显著高于其他退化程度的草地(P<0.05)。从L、H到E草地,当年生地上枝条数虽无显著差异,但其数量和地上生物量均有随草地退化程度的加剧而减小的趋势。可见,大针茅地上部分的小型化和其根系构型的小型化是同步的,且后者起决定作用。
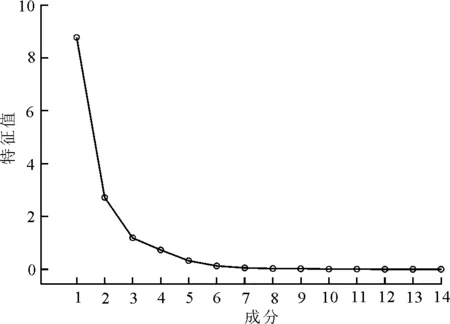
图1 各成分特征值碎石图

表2 大针茅14个根系构型参数主成分分析
4 讨论与结论
本研究表明根分叉数、根表面积和总根长3个参数可作为分析大针茅根系构型在不同退化程度的草地中变化的主要指标,其中根分叉数对该变化的解释力最强。根分叉数囊括了所有侧根与其上一级根之间的分叉数,分叉数越多根系构型越繁杂,则根系在土壤空间中的分布范围就越大,越有利于根系吸收代谢及固土蓄水能力的发挥;根系表面积直接反映了根与土壤的结合面积,结合面积越大则越利于根系对无机营养物质的吸收;总根长反映了根系生长及其在土壤中拓殖的能力。这3个参数集中反映了大针茅根系对土壤空间异质性的适应能力。专门对大针茅根系进行研究,国内文献报道并不多,而且针对天然草地中大针茅根系的研究少之又少,而多数集中于地上部分的研究,如贾美清等[15]对大针茅的叶性分析;吴建波等[16]分析了不同围封年限对大针茅种群及个体形态(生殖枝、营养枝等地上部分)的影响研究;仅见高慧等[17]有针对性的对大针茅地下部分进行了研究,选用的指标为地下生物量。本研究所得出的分析大针茅根系构型的指标(根分叉数、根表面积和总根长)是首次提出,尤其根分叉数能更好的体现大针茅根系对土壤空间异质性的适应能力。

表3 根分叉数、根表面积、总根长、当年生地上枝条数和地上生物量的方差分析
从轻度退化草地、重度退化草地到极度退化草地,大针茅的根幅、根深、不定根条数、株丛分蘖节宽度、根分叉数、根表面积、总根长和地下生物量均有减小的趋势,即大针茅的根系构型具有明显小型化的趋势。根系构型的小型化在一定程度上会影响地上部分的生长繁殖。据王炜等[5]对羊草+大针茅群落退化演替机理的研究发现,植物个体小型化及其生态学作用可认为是揭示在过度放牧条件下群落优势种演替的机理,该植物个体小型化包含根系分布的浅层化现象,但主要是指地上部分的小型化。而本研究表明,随草地退化程度的加剧大针茅根系构型小型化,是地上部分小型化的诱因。因此,可以认为根系构型的小型化及其生态作用是最终导致植被优势种在环境胁迫条件下发生退化演替的直接原因。
本研究发现,中度退化草地中大针茅根系构型小型化的现象尤为明显,其根幅、根深、不定根条数、株丛分蘖节宽度、根分叉数、根表面积、总根长、地下生物量和地上生物量等参数都小于其他退化程度的草地。羊草杂类草草甸也表现为中度放牧强度地上生物量小于轻度和重度放牧强度草甸[18]。野外调查发现,中度退化程度的草地中的物种多样性要高于其他3个退化程度的草地,符合中度干扰理论[15-16],且植物生长密集并普遍小型化,这必然会使得植物根系对土壤中有限的资源进行激烈竞争,使根系的生长在很大程度上受到限制[19-20],加之中度退化程度的草地中分布着家畜喜食的牧草,增加了家畜对大针茅的采食频度,同时家畜频繁践踏草地使土壤紧实度增加,因此使大针茅株丛的地下部分明显小型化。虽然草地处于中度退化阶段,但大针茅的根系生长状况甚至要比极度退化程度的草地中的差。重度和极度退化程度的草地中,植被盖度相对较低,有更多的土壤资源被释放出来供应大针茅根系的生长,因此其根系的生长状况要好于中度退化阶段的草地。
根系构型小型化势必会导致地上部分小型化,最终导致整个植物体小型化,并在群落退化演替进程中使大针茅的建群种优势作用逐渐减弱并最终为其他优势种取代,如冷蒿和星毛委陵菜,后果是草地载畜量下降,裸地面积增加,出现不同程度的草地沙化,使生态系统稳定性降低。从这种意义上讲,可以认为草地沙化与根系构型小型化之间存在一定的关联性,可为防治草地沙化提供借鉴。
[1]中国科学院内蒙古宁夏综合考察队.内蒙古植被[M].北京:科学出版社,1985:495-515.
[2]内蒙古植物志编辑委员会.内蒙古植物志(第五卷)[M].第二版.呼和浩特:内蒙古人民出版社,1994:198-199.
[3]李博.中国北方草地退化及其防治对策[J].中国农业科学,1997,30(6):1-9.
[4]安渊,徐柱,阎志坚,等.不同退化梯度草地植物和土壤的差异[J].中国草地,1999(4):31-36.
[5]王炜,梁存柱,刘钟龄,等.羊草+大针茅草原群落退化演替机理的研究[J].植物生态学报,2000,24(4):468-472.
[6]张红梅,赵萌莉,李青丰,等.放牧条件下大针茅种群的形态变异[J].中国草地,2003,25(2):13-17.
[7]宗振宏.不同群落状态下典型草原植物抗旱能力的比较研究[D].呼和浩特:内蒙古大学,2009:7-9.
[8]Lynch J P.Root architecture and plant productivity[J].Plant Physiology,1995,109:7-13.
[9]毛齐正,杨喜田,苗蕾.植物根系构型的生态功能及其影响因素[J].河南科学,2008,26(2):172-176.
[10]Staswick P E,Serban B,Rowe M,etal.Characterization of anArabidopsisenzyme family that conjugates amino acids to indole-3-acetic acid[J].Plant Cell,2005,17:616-627.
[11]陈世鍠,张昊,王立群,等.中国北方草地植物根系[M].长春:吉林大学出版社,2001:87-88.
[12]孙儒泳,李博,诸葛阳,等.普通生态学[M].四川:高等教育出版社,1993:132-135.
[13]张铜会,赵哈林,大黑俊哉,等.连续放牧对沙质草地植被盖度、土壤性质及其空间分布的影响[J].干早区资源与环境,2003,17(4):117-121.
[14]冯秀,仝川,张鲁,等.内蒙古白音锡勒牧场区域尺度草地退化现状评价[J].自然资源学报,2006,21(4):575-583.
[15]贾美清,高玉葆,杨勇.内蒙古中东部草原克氏针茅和大针茅的叶性分析[J].天津师范大学学报(自然科学版),2010,30(1):57-63.
[16]吴建波,包晓影,李洁,等.不同围封年限对典型草原群落及大针茅种群特征的影响[J].草地学报,2010,18(4):490-495.
[17]高慧,高玉葆,刘海英,等.不同坡位大针茅生长与生殖分配特征[J].应用生态学报,2009,20(9):2123-2128.
[18]韩文军,春亮,侯向阳.过度放牧对羊草杂类草群落种的构成和现存生物量的影响[J].草业科学,2009,26(9):195-199.
[19]Collins S L.Experimental analysis of intermediate disturbance and initial floristic composition: decoupling cause and effect[J].Ecology,1995,76:486-492.
[20]Connell J H.Diversity in tropical rain forests and coral reefs[J].Science,1978,199:1302-1310.
猜你喜欢
小小说月刊·下半月(2021年8期)2021-09-05 05:49:31
中学生数理化(高中版.高考理化)(2021年12期)2021-03-08 00:48:08
动漫界·幼教365(大班)(2018年3期)2018-05-14 11:50:46
山东农业科学(2017年4期)2017-05-18 17:06:13
北京航空航天大学学报(2017年10期)2017-04-20 08:51:23
中国卫生(2014年7期)2014-11-10 02:33:02
航天返回与遥感(2014年4期)2014-07-31 17:47:47
山东农业科学(2014年2期)2014-03-24 12:05:43
无机化学学报(2014年4期)2014-02-28 17:31:08
汽车与新动力(2012年5期)2012-03-25 10:09:40
