
温度和盐度分别对尖紫蛤胚胎发育的影响
2011-03-14 06:15黄海立吕广煊刘志刚
海洋科学 2011年10期
黄 洋,黄海立,吕广煊,刘志刚
(1.广东海洋大学 海洋高新科技园,广东 湛江 524025; 2.广东海洋大学 广东高校海产经济无脊椎动物健康养殖工程研究中心,广东 湛江 524025)
尖紫蛤 (Soletellina acutaCai et Zhuang)俗称“沙螺”或 “西施舌”[1],隶属软体动物门、瓣鳃纲、帘蛤目、紫云蛤科(Rsammobiidae),分布于我国福建和广东沿海,生活在河口咸、淡水交汇处[2],尤以广东省吴川市鉴江河口咸、淡水交汇处产量最多,最高年产量达 5万 kg[3]。其生活在河口河床、滩涂从低潮线附近直至2~3 m深的水域营潜沙生活,在江河入海的内湾繁殖生长。尖紫蛤肉嫩味美,营养价值高,据分析蛤肉干含蛋白质 58.65%,糖类 5.31%,脂类8.75%[4],且有药用功效。20世纪90年代以来,由于在鉴江河口咸、淡水交汇处过度采捕和水质污染,90年代后期,尖紫蛤自然资源濒临灭绝[4]。积极开展其生物学及增养殖研究是恢复其自然生物量,保护河口重要经济品种和海洋生物多样性的有效途径之一。
目前,国外关于尖紫蛤人工育苗和增养殖的研究内容尚未见报道。国内关于尖紫蛤的报道仅限于蔡英亚等[1-2,5]进行了广东鉴江尖紫蛤的生态调查,和尖紫蛤滩涂(中、低潮区)人工养殖试验; 杨耀聪等[4]采用组织切片方法研究报道了福建省尖紫蛤的繁殖期; 符韶等[3]进行尖紫蛤的人工育苗试验。但关于尖紫蛤胚胎发育的资料国内外未见有报道,水温和盐度直接影响到胚胎发育的速度和质量(孵化率和畸形率)。因此,进行尖紫蛤胚胎发育的研究,可为尖紫蛤人工繁育和增养殖研究提供理论依据。
1 材料与方法
1.1 实验材料
实验在广东海洋大学东海岛海洋生物研究基地进行。亲贝采自广西山口镇洗米河口,利用塑料筐铺沙悬空流水法促熟培育。待亲贝性腺成熟后进行催产,水温26.3 ℃,盐度17。挑选个体大、性腺饱满并开始排放的雌雄亲贝各一个分别放入装有过滤海水的塑料桶中排放5 min,然后混合受精并记录时间,混合后10 min用清洁海水洗卵去除多余的精子作为实验材料,洗卵和分卵过程在10 min内完成。
1.2 实验方法
1.2.1 尖紫蛤胚胎发育的观察
在水温27 ℃、盐度17的环境下,从精卵混合后每隔 5 min 用吸管吸取受精卵或胚胎置于载玻片上,在显微镜下观察卵子的受精及胚胎发育情况,记录不同胚胎发育阶段的发育时间、胚体大小及特点,并对不同胚胎发育阶段的胚体进行拍照,胚胎发育观察至直线铰合期。
1.2.2 温度实验
温度实验设6个梯度,依次为17,20,23,26,29,32 ℃,实验海水盐度调节至16,pH为8.2,光照强度约 500~1 000 lx。受精卵在容积为1 000 mL的烧杯中进行孵化实验,密度为 10 个/mL,每个温度梯度设有3个平行组。为了减少温差的刺激,各试验组水温均从26 ℃开始,按每5 min升或降 1 ℃达到各自设定的试验水温。试验过程不充气,每隔10 min用玻璃棒搅动一次保证水体中充足的溶氧。
1.2.3 盐度实验
盐度实验设8个梯度,依次为2,5,8,11,14,17,20,23,温度保持在 27 ℃。pH 为 8.2,光照强度约500~1 000 lx。受精卵在容积为1 000 mL的烧杯中进行孵化实验,密度为 10个/mL,每个盐度梯度设有3个平行组。为了减少盐度变化的刺激,各试验组盐度均从16开始,按每3 min升或降 1达到各自设定的试验盐度。试验过程不充气,每隔10 min用玻璃棒搅动一次保证水体中充足的溶氧。
1.2.4 实验用水的处理
海水从海区直接抽取,淡水采用地下井水,试验组用水由盐度 31的自然海水添加地下井水配制,所有实验用水均经过0.45 μm微孔滤膜过滤。温度试验控温装置采用白色泡沫箱、500 W 石英加热管、1 000 W 电子继电器、0~-50℃水银触点温度计(±0.1℃)组成,所有温度计均经过严格校正,低温端用冰袋降温及超温报警装置。盐度实验组使用误差±0.5℃的恒温水槽控制水温。
1.2.5 评价方法与主要观测指标
以孵化率不低于 60%作为胚胎发育适宜温、盐度范围的衡量指标; 以孵化率不低于 80%作为其最适温、盐度范围的衡量指标。孵化时间:从受精卵开始发育到直线铰合期幼虫所经历的时间。孵化率:胚体发育到直线铰合期幼虫与受精卵总数的比率。幼虫畸形率:畸形幼虫与直线铰合期幼虫总数的比率。
1.2.6 数据处理
采用 spss13.0 统计处理软件对实验数据进行处理和分析。

表1 尖紫蛤胚胎发育进程Tab.1 Embryonic development of Soletellina acuta Cai et Zhuang
2 结果
2.1 尖紫蛤胚胎发育进程
结果见表1和图1,尖紫蛤精卵接触后即行受精,受精卵产生一层透明的受精膜,卵核模糊。在孵化水温27 ℃,盐度17条件下,受精后30 min,在卵子动物极出现第一极体随后出现第二极体。受精后 65 min开始第一次分割成为 2细胞期,以后继续分割经过4细胞期、8细胞期、16细胞期、32细胞期,经6次分裂胚体呈桑椹状,到达桑椹期。卵裂继续,至受精后 280 min左右进入囊胚期,胚体发育成为圆球形,周身长出细小纤毛,开始在水中做顺时针旋转,运动缓慢4s/圈,运动螺径250 μm左右,上浮于水体中,有集群习性。胚体继续发育,至受精后430 min左右胚体长出一纤毛环,中央具鞭毛束,称为担轮幼虫,以600 μm直径作不规则的圆周运动,部分做直线运动,游动迅速。历时710 min左右,胚体开始分泌原壳,消化系统开始分化,运动器官面盘已具雏形,此时为面盘初期幼虫。历时 980 min左右,胚体的面盘、摄食消化器官完全形成,贝壳完全覆盖身体,绞合部平直,卵黄颗粒丰富,称为直线绞合期幼虫。
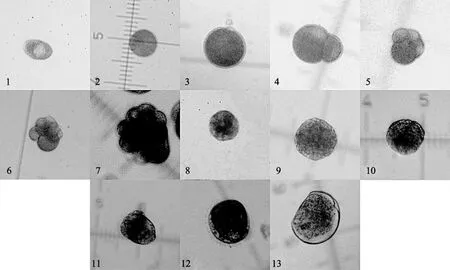
图1 尖紫蛤胚胎发育各期的形态Fig.1 Embryonic development of Soletellina acuta Cai et Zhuang in each stage
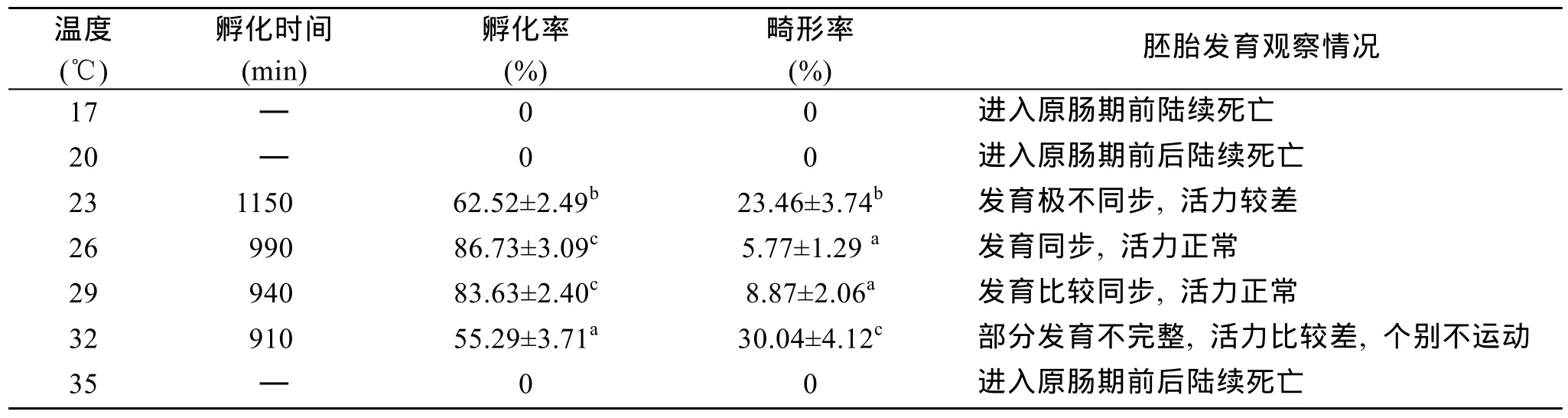
表2 不同温度下尖紫蛤胚胎发育的情况Tab.2 Embryonic development of Soletellina acuta Cai et Zhuang in different temperatures
2.2 温度对尖紫蛤胚胎发育的影响
表2和图2是不同温度下尖紫蛤的孵化时间跟孵化率情况。由表2可见,水温低于20 ℃和高于35 ℃的实验组胚胎发育至原肠期前后陆续死亡,17 ℃实验组绝大部分胚胎发育至 8细胞期便停止了发育;水温在23~32 ℃之间胚胎均能发育到直线铰合期幼虫。水温为23 ℃的实验组,孵化时间最长1 150 min,孵化率达到62.52 %,畸形率为23.36%; 水温为32 ℃的实验组,孵化时间最短 910 min,但孵化率只有55.29 %,畸形率高过23 ℃组,幼虫运动迟缓; 26 ℃和29 ℃实验组孵化时间比较接近,孵化率差别不大,无显著差异(P>0.05),分别是86.73 %和83.63 %,畸形率都低于10 %。尖紫蛤的胚胎发育在适宜温度范围内,孵化水温(x)与孵化时间(y)存在着多项式函数关系:y=3.6111x2-224.28x+ 4393.6(R2=0.9882)。孵化水温(x)与孵化率(y)间存在着多项式函数关系:y=-1.5858x2+86.982x-1103.1(R2=0.9963)(图 2)。由此可以推算出,尖紫蛤胚胎发育适宜温度范围在23.1~31.2 ℃(孵化率在60 %以上),尖紫蛤胚胎发育最适温度范围在25.0~29.9 ℃(孵化率在80 %以上)。

图2 盐度为16时尖紫蛤胚胎孵化时间、孵化率和畸形率与水温的关系Fig.2 Relationship between temperature and the incubation duration,the percentage of hatching and abnormal larvae of Soletellina acuta Cai et Zhuang in 16 degree salinity

表3 不同盐度下尖紫蛤胚胎发育的情况Tab.3 Embryos development of Soletellina acuta Cai et Zhuang in different salinities
2.3 盐度对尖紫蛤胚胎发育的影响
表3和图3是不同盐度下尖紫蛤的孵化时间和孵化率情况。盐度为2的实验组大部分胚胎在 2细胞期和4细胞期便停止了发育,少数发育至8和16细胞期停止了发育,在原肠期前便全部死亡; 盐度为5和26的实验组胚胎虽然能发育到原肠期但不能发育到面盘幼虫初期; 当盐度为8和23时孵化时间非常接近,分别为1 150 min和1 140 min,胚胎能发育到直线铰合期,但孵化率非常低分别为 20.59%和22.48 %,幼虫活力很差,畸形率非常高达到71.61%和67.59 %,因此尖紫蛤胚胎发育的最低和最高临界盐度为8和23。
盐度为11~20的实验组孵化时间无显著差异(P>0.05),在1 000 min左右,盐度为14~17的实验组孵化率无显著差异(P>0.05),都达到80 %以上,孵化出来的幼虫活力正常,而且畸形率小于 10%,差异不显著; 盐度为11和20的实验组虽然孵化率超过80%,但是其畸形率偏高超过 10%,故可认为盐度 11~20为尖紫蛤胚胎发育适宜的盐度范围,14~17为尖紫蛤胚胎发育的最适宜的盐度范围。

图3 温度为27℃时尖紫蛤胚胎孵化时间、孵化率和畸形率与盐度的关系Fig.3 Relationship between salinity and the incubation duration,the percentage of hatching and abnormal larvae of Soletellina acuta Cai et Zhuang in 27℃ Ambient temper ature
3 讨论
温度和盐度是影响海洋贝类幼虫生长的两个重要的环境因素[6],因其容易测量和调节而引起生态学家的关注。林瑞才[7]等学者进行温度和盐度对海湾扇贝幼虫附着变态影响的研究后发现,在不同盐度下,幼虫附着变态的适宜温度不同,反之亦然,但也有学者认为,只有温度和盐度或其中之一接近极限范围时,温度和盐度相结合才显示出明显的相互关系,反之,当温度或者盐度处于安全范围时,没有明显的关系[8]。本研究将温度或盐度控制在适宜的条件下,研究尖紫蛤胚胎对单一因子的耐受范围。关于温度和盐度对该种类胚胎发育的互作效应,有待后续研究。
从实验结果可以看出:温度、盐度既影响尖紫蛤胚胎发育的孵化率又影响其发育速度; 尖紫蛤胚胎发育的适宜温度范围为23.1~31.2 ℃,适宜盐度范围为 11~20,表明尖紫蛤孵化的盐度范围比较宽,其所需的温度比较高; 温度超过其适宜范围时,孵化率明显下降或幼虫畸形率明显增大,即:在一定范围内,提高温度可缩短孵化时间,但进一步提高了温度引起畸形分割[9-12],产生败育; 温度低于适温下限,胚胎发育则产生滞育[13]。
目前,国内对尖紫蛤的研究还不多,因此对其进行人工催产技术、胚胎发育以及最适饵料等方面的研究尤为重要。在温度试验中知道,其胚胎发育需要有较高的温度,一方面,尖紫蛤属于热带、亚热带种,但温度过高(高于33 ℃),其发育就停止,尖紫蛤繁殖季节在9~11月,最高温的7~8月并不是其繁殖期[3],也与此相符。 在7、8月暂养期间,室外的温度曾经处于35 ℃以上,结果发现尖紫蛤钻出到沙面上,两贝壳打开,活力变差,还出现陆续死亡现象。所以从温度实验32 ℃的实验组里也能知道,胚胎虽能够孵化出来,但只有个别是能够发育到直线铰合幼虫。在盐度试验中,盐度低于8的实验组胚胎不能正常发育,在亲贝采集地进行资源调查时发现,退潮时在尖紫蛤自然分布区盐度有时只有3,远远低于尖紫蛤胚胎发育所需的盐度; 尖紫蛤属于潜沙贝类,并没有迁移的习性,稚贝和成贝对盐度的适应性会不会更强有待进一步的研究。
[1]蔡英亚,邓陈茂,刘志刚.广东鉴江尖紫蛤的生态调查[J].湛江水产学院学报,1992,12(1):7-11.
[2]蔡英亚,庄启谦.紫云蛤科一新种[J].热带海洋,1985,4(3):64-65.
[3]符韶,蔡英亚,邓陈茂,等.尖紫蛤的人工育苗[J].湛江海洋大学学报,2000,20(1):15-17.
[4]杨耀聪,李复雪.尖紫蛤生殖周期的研究[J].热带海洋,1994,13(2):61-67.
[5]蔡英亚,邓陈茂,符韶.西施舌的人工养殖[J].中国水产(台),2002,592:50-52.
[6]包永波,尤仲杰.几种环境因子对海洋贝类幼虫生长的影响[J].水产科学,2004,23(12):39-40.
[7]林瑞才,陈敏.温度和盐度对海湾扇贝幼虫附着及变态的影响[J].台湾海峡,1989,8(1):60-67.
[8]刘建勇,卓健辉.温度和盐度对方斑东风螺胚胎发育的影响[J].湛江海洋大学学报,2005,25(1):3-4.
[9]陈文龙,刘德经,许万竹,西施舌人工育苗的初步研究[J].水产学报,1966,3(2):130-138.
[10]林笔水,吴天明.温度和盐度对缢蛏浮游幼虫发育的影响[J].生态学报,1984,4(4):385-392.
[11]何义朝,张福绥.贻贝胚胎发育的有效温度范围的变化[C]// 中国贝类学会.贝类学论文集(2).北京:科学出版社,1986:89-93.
[12]谢玉坎,张偲,陈国和,等.温度变化影响大珠母贝幼虫的发育[J].热带海洋研究,1988,3:131-133.
[13]刘德经,陈杰明.西施舌早期胚胎发育温度效应的研究[J].动物学杂志,1998,33(2):2-3.
猜你喜欢
汽车实用技术(2022年14期)2022-07-30
中国蚕业(2022年1期)2022-03-09
今日农业(2021年6期)2021-11-27
电子制作(2019年7期)2019-04-25
电子制作(2018年17期)2018-09-28
广东海洋大学学报(2015年3期)2015-12-22
华南农业大学学报(2015年5期)2015-12-04
现代畜牧科技(2015年12期)2015-10-21
中国洗涤用品工业(2015年5期)2015-02-28
创业家(2015年3期)2015-02-27
