
凿贝才女虫形态与结构观察
2011-03-14 06:15杨红生张晓芳
海洋科学 2011年10期
高 燕,张 涛,杨红生,张晓芳
(1.中国科学院 海洋研究所,山东 青岛,266071; 2.中国科学院 研究生院,北京,100049; 3.獐子岛渔业集团股份有限公司,辽宁 大连,116001)
底播是中国贝类滩涂和浅海增养殖的重要方式之一,具有成本低、贝类生长快、效益高,产品质量高等优点。但是底播增养殖的贝类常常受到才女虫(Polydora ciliata)病的困扰。才女虫分泌腐蚀贝壳的酸性物质在贝壳上穿凿管道,使壳内面接近中心部位形成黑褐色的痂皮,俗称“黑壳”病或“黑心肝”病。病贝生长缓慢,由于管道的形成,使贝壳受损,特别使闭壳肌周围的壳变得脆弱,在养殖操作过程中容易破裂。当虫体钻穿贝壳达到软体部时,则直接侵害软体部,被侵组织周围发生炎症,局部形成脓肿和溃疡,引起细菌继发性脓疡,并产生一种特殊的臭味,大大降低了贝类的品质和价值,严重时导致贝类死亡。才女虫是贝类增养殖业最重要的病害生物之一,在世界范围内广泛分布,但至今尚未找到有效的防控措施[1-4]。
凿贝才女虫属于环节动物门(Annelida),多毛纲(Polychaeta),海稚虫科(Spionidae),才女虫属(Polydora)。凿贝才女虫在含有大量碳酸钙的基质中筑造管道,营管栖生活,在欧洲、美洲、亚洲、新西兰等地都有广泛分布,严重危害贝类养殖业及珊瑚礁的生长,并致大规模死亡[5-7]。在国内,关于凿贝才女虫对扇贝、珍珠贝、鲍等的危害已有报道[8-10],但是对本种乃至本属的研究,一直未得到应有的重视,长期以来一直停留在分类学的初始阶段。为了更深入地开展对该类群的研究,本实验选择了辽宁獐子岛底播虾夷扇贝养殖区作为研究基地,对严重感染凿贝才女虫的病贝的症状、虫体外部形态和内部结构进行了较为系统地研究,旨在丰富多毛类形态和结构的基础知识,为科学防控才女虫病提供理论依据。
1 材料与方法
1.1 试验用扇贝来源
于 2007年 10月在辽宁大连獐子岛海域(水深>30 m)通过底拖网收集感染有凿贝才女虫的虾夷扇贝,带回实验室后放入 20℃海水中暂养,每天投喂两次混合单胞藻。
1.2 光镜观察
1.2.1 活体观察
用钳子将贝壳掰碎后将凿贝才女虫从管道中取出,放入加有少量海水的培养皿中,置于解剖镜下观察。为方便观察,逐滴加入7%的MgCl2海水溶液至虫体麻醉。
1.2.2 组织学切片观察
取凿贝才女虫活体,用波恩氏液(Bouin’s)固定18~24 h。采用石蜡整体包埋切片法。主要步骤包括:固定、冲洗、脱水、透明、浸蜡、包埋、切片、染色(H.E.染色)、封片等。全部观察、测量、拍照均使用日本尼康显微成像系统Nikon80i完成。
1.3 扫描电镜观察
凿贝才女虫成虫用4%的戊二醛(0.1 mol/L PBS,pH 7.2-7.4)溶液于 4℃下固定。乙醇梯度脱水后入100%醋酸异戊酯中置换,临界点干燥仪干燥,IB-3型离子溅射仪喷金,在 KYKY2800B扫描电镜下观察。
1.4 透射电镜观察
取表皮、肌肉层和肠组织块各1 mm3,用2.5%戊二醛和1%锇酸双重固定,梯度乙醇脱水,Epon812环氧树脂包埋。切片厚0.05 µm,醋酸铀和柠檬酸铅双重染色,日立H-7000 透射电镜观察并拍照。
2 结果
2.1 病贝的症状
被凿贝才女虫感染的虾夷扇贝上壳的外表面有针尖大小的小孔,一般两个一组,为凿贝才女虫在贝壳中筑造管道的入口和出口。将病贝放入洁净的海水中静止片刻,肉眼可见有凿贝才女虫长长的触手从孔中钻出,受刺激马上缩回到管道中。将病贝肉体去除,可见其上壳的内表面有弯弯曲曲隆起的管道,呈黄褐色,用镊子掰开后可见管道内有大量絮状碎屑(图1)。

图1 被凿贝才女虫感染的虾夷扇贝上壳的外表面和内表面Fig.1 The left shell of Patinopecten yessoensis(1-1)showing the mud tubes on the outer surface of the shell,(1-2) showing the prominent knobs on the inner surface of the shell
2.2 凿贝才女虫形态特征
凿贝才女虫成虫虫体长一般为 10~40 mm,体宽0.7~1 mm,头部有1对触手,可以自由收缩,触手边缘有黑色波纹带。身体分节,但是没有明显的分区。在解剖镜下,可清楚地看见其背面中央充满红色血液的血管贯穿身体前后端,并有分支血管将血液输向各疣足部分。从颜色上区分,成虫有橙黄色,草绿色,棕褐色,身体前端为橙黄色,后部为棕褐色等4 种(图 2)。
凿贝才女虫围口节前端有口前叶,前端圆钝且分叉,摄食时通过纤毛的摆动,使食物顺着食物沟送入消化道中(图 3-1)。第二至第四刚节每一节两侧都生有运动性的长刚毛,对才女虫的身体起支撑作用,背部有分泌腺囊,可以分泌大量黏液(图3-2)。第五刚节大且变形、仅具一种变形粗足刺刚毛,且排成一直排或稍弯曲,通常伴随刚毛(图 3-3)。从第七刚节开始身体两侧有疣足和伴随运动性刚毛,背部生有一排约5~7个棘状刚毛,疣足向腹部弯曲,腹部每个刚节都有一条纤毛带(图3-4,3-5)。尾部呈吸盘状,肛部为收缩或扩张的袖口状、茶碟状或分叶,表面有大量纤毛,中间有肛门孔(图3-6)。触手长约1~2 mm,生有一条浅浅的食物沟,沟内布满纤毛,摆动时可以滤取海水中的食物颗粒,触手表面有大量乳突(图3-7,3-8)。

图2 凿贝才女虫成虫Fig.2 Photograph of Polydora ciliate
2.3 凿贝才女虫结构特征
凿贝才女虫身体结构相对简单,主要由表皮、肌肉层、消化系统组成。体腔内充满体腔液,肠壁外附有肌肉。肌肉层发达,每一体节具一对疣足,疣足上生有刚毛,运动能力比较强。肌肉组织为平滑肌,呈梭型,分为环肌、纵肌和斜肌。个体被横切时,环肌为外侧环生,在疣足处间断肠壁上附生的肌肉亦为环肌; 内层纵肌发达,横切呈多边形; 每个体节内,还有一对斜肌。表皮层主要由柱状上皮细胞和许多散在的腺细胞组成。柱状上皮细胞呈柱状,细胞核近圆形,位于细胞基底。柱状细胞间有许多散在的腺细胞,腹侧面和疣足叶基部腺细胞较多。腺细胞呈空泡状,表明在细胞中含有很多囊泡,胞核位于基部,为扁圆形; 表皮下的基膜不明显(图 4-1,4-2,4-3)。身体纵切显示消化道结构简单,由柱状上皮细胞组成,细胞排列紧密,外面包裹着肌层,内环外纵。疣足为每节体壁向两侧垂直伸出的肉质扁平叶。疣足壁结构和体壁一样,在疣足基部有刚毛囊,其内的细胞能分泌几丁质性质的刚毛(图4-4,4-5,4-6)。
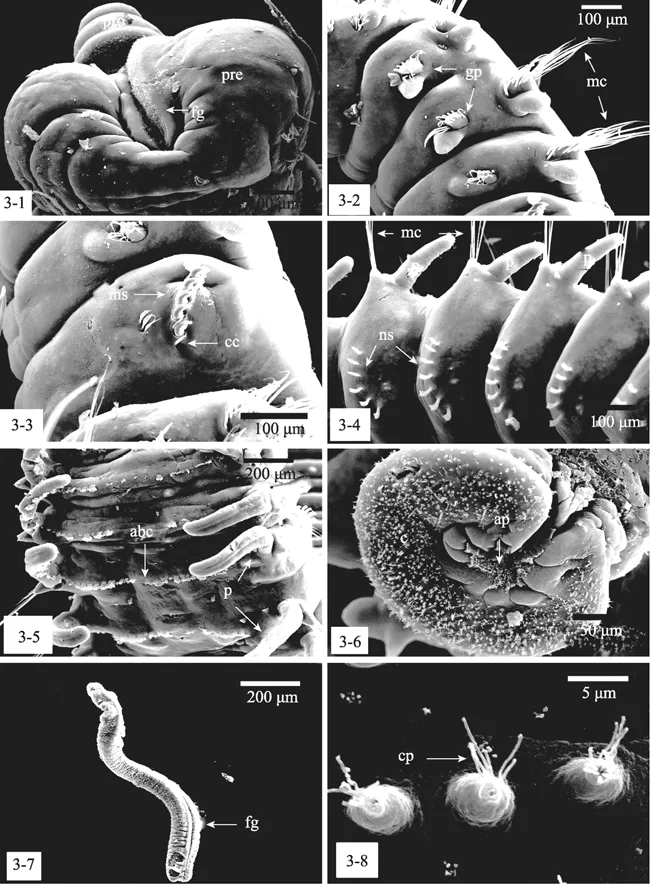
图3 凿贝才女虫扫描电镜观察Fig.3 Scanning electron micrographs of Polydora ciliata
凿贝才女虫表皮细胞外层着生有大量的微绒毛;在透射电镜下皮肤表层有细小的颗粒,表明皮肤外有黏液层,比较润滑(图 5-1); 腺细胞向表皮分泌的黏液(图 5-2)。凿贝才女虫的肌肉层透射电镜照片显示肌浆网内有大量的脂肪颗粒; 肌肉的横纹是因为组成肌纤维的原纤维上具有规则的、相间排列的明带和暗带的原故,电镜显示,每一条肌原纤维还含有许多称为肌丝的结构,一类为肌动蛋白肌丝,一类为肌球蛋白肌丝,这些肌丝在肌肉组织中的排列是很有规则的,因而产生肌原纤维的横纹样式。长度相等的肌原纤维也作规则的排列,当肌原纤维的横纹均等的隔开时,便使整根肌纤维具有规则的横纹。才女虫的 Z线呈断裂状,肌球蛋白和肌动蛋白连接不紧密(图 5-3)。凿贝才女虫的消化道内有大量的微绒毛,消化能力强(图 5-4)。消化道的腺细胞内含有很大的溶酶体颗粒,向消化内分泌消化酶(图 5-5)。在消化道内发现大量单细胞藻类,为凿贝才女虫与扇贝争食夺饵提供了证据(图5-6)。
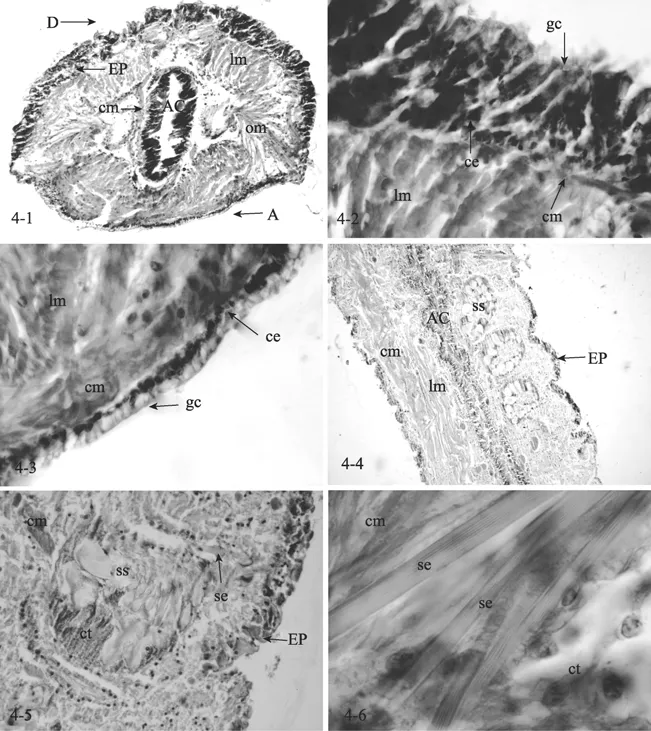
图4 凿贝才女虫组织学观察Fig.4 Histological observation of Polydora ciliata
3 讨论
3.1 凿贝才女虫对虾夷扇贝贝壳的损害
凿贝才女虫在贝壳内钻凿,分泌黏液黏附周围的颗粒碎屑铸造向上泥管,这些泥管的开口处高出贝壳表面1~2 cm(图1-1)。在贝壳的内表面可以观察到褐色或深褐色的弯弯曲曲的突起(图 1-2)。这些突起的形成是由于凿贝才女虫钻凿刺激扇贝不断分泌碳酸钙,包裹住虫体,阻止其向里面进一步钻凿。这些突起的形状是不规则的,在被才女虫侵蚀严重的贝壳上,这些突起往往连成一片,观察到的最大的一个突起直径约 4 cm。位于贝壳边缘的突起,阻碍扇贝上下壳合拢,严重影响了扇贝正常生活。这些管道使得贝壳变脆,容易破碎,减弱了其对软体部位的保护功能。

图5 凿贝才女虫透射电镜照片Fig.5 Transmission electron micrographs of Polydora ciliata
3.2 凿贝才女虫的体壁结构
就体壁的结构组成而言,凿贝才女虫与环节动物门多毛纲常见生物相似,体壁从外向内分4层: 角质层、表皮、肌肉层和壁体腔膜[11]。凿贝才女虫体壁表皮为单层柱状上皮细胞,其中夹有腺细胞和感觉细胞,尤以腹面和疣足叶基部的腺细胞较多。表皮细胞内含有大量的线粒体表明表皮细胞代谢旺盛(图5-1)。在其他多毛类表皮结构中也常常出现含有大量液泡的腺细胞[12]。
在大部分多毛类中,刚毛经常横向排列为带状或纵向排列为束状。在体型较小的种类中刚毛用来移动身体,在体型较大的种类中,刚毛的摆动还可以促进周围水流的流动[13]。凿贝才女虫背刚毛、腹刚毛和疣足上的刚毛围绕身体形成一个刚毛环; 第五刚节刚毛变形,形成爪状结构。推测这些刚毛的运动可以把贝壳碎屑摩擦下来,从而形成光滑弯曲的管道。Okoshi[14]描述了虾夷扇贝和长牡蛎被才女虫Polydora variegate,P.websteri,P.converxa和P.concharum侵蚀贝壳的超显微结构。在4种才女虫侵蚀贝壳形成的管道表面都发现同心的孔,在管道的侧面有与管道方向平行的抓损的刮痕。Haigler[15]认为才女虫凿贝钻孔的机制包括两个过程: 分泌黏液化学腐蚀和刚毛机械磨损。
疣足为体节体壁向两侧垂直伸出的肉质扁平叶,有触觉和运动功能。疣足内充满毛细血管,在疣足的顶部形成了毛细血管网。才女虫没有特殊的呼吸器官,微血管网则成为气体交换的主要场所。在疣足基部有刚毛囊,其内的形成细胞能分泌几丁质性质的刚毛。刚毛和疣足是环节动物的运动器官。大多数环节动物都具有刚毛,海产种类一般有疣足[11]。
3.3 凿贝才女虫形态结构与其穴居生活的关系
凿贝才女虫表皮的腺细胞向表皮分泌的黏液,含有酸性物质,有腐蚀贝壳和润滑身体的作用(图5-2)。感觉细胞分散在表皮中,可感受外界的刺激。肌肉层是由环肌、纵肌和斜肌组成,与环毛蚓只有环肌和纵肌不同。在功能上,环肌的收缩可以使虫体变细长,纵肌的收缩使虫体变粗短。斜肌的收缩使身体运动更灵活[16]。由于凿贝才女虫的身体是按节排列的,而斜肌的位置在每节体壁内侧的连接处,可能与才女虫体节之间的连接有关。凿贝才女虫的肌肉层透射电镜照片显示肌浆网内有大量的脂肪颗粒,说明肌肉兼有能量储备的功能。才女虫的 Z线呈断裂状,肌球蛋白和肌动蛋白连接不紧密,说明才女虫的运动能力弱,只能蠕动,与其穴居的生活方式相适应。
才女虫是典型的海底食碎屑动物,大部分种类生活在近岸海域,深海中比较少。才女虫有两种摄食方式,吞食和滤食。吞食主要是吞咽周围含有机质的泥沙,吸收其中的营养物质,滤食是用触手搅动周边的海水,摄食其中小的有机物颗粒。很多种类在这两种摄食方式之间可以自由转化,主要取决于其周围的环境。触手是摄食时的重要器官[17]。凿贝才女虫的触手来回摆动收集悬浮在水中的的颗粒,沿着触手的内表面是一个从一端延伸到另一端的长满纤毛的凹槽(图3-7,3-8)。触须末端的纤毛摆动,把颗粒向口中驱赶。
凿贝才女虫消化道发达,肠黏膜突起形成极其发达的指状绒毛,极大地增加了消化和吸收面积。肠的纤毛沟贯穿肠的前后,其中纤毛多而密集(图 5-4),在纤毛的摆动下,可以促使食物向身体后方运输,相对弥补了肠部肌肉不发达造成的对食物运输的不足。超微结构显示肠纤毛沟细胞中含有少量的酶原颗粒和酶活性,亦表明具有一定的消化功能。消化管中的黏液物质,除具润滑作用和保护作用外,有的学者认为中性黏液物质的存在与吸收有关,而酸性黏液物质的存在则与分泌有关[18]。超微结构显示肠的柱状上皮细胞具微绒毛、质膜内陷和游离端出现大量的胞饮泡、细胞中部出现进行物质降解的次级溶酶体和细胞内含有储存物质,这些都是吸收细胞的超微结构特征。
[1]Riascos J M,Heilmayer O,Oliva M E,et al.Infestation of the surf clamMescidesma donaciumby the spionid polychaetePolydora bioccipitalis[J].Journal of Sea Research,2008,59(4): 217-227.
[2]Silina A V.Tumor-like formation on the shells of Japanese scallopsPatinopecten yessoensis(Jay)[J].Marine Biology,2006,148: 833-840.
[3]Vargas L,P Quijón,Bertrán C.Polychaete infestation in cultured abalone (Haliotis rufescens Swainson) in Southern Chile [J].Aquaculture Research,2005,36(7):721-724.
[4]Mcdiarmid H,Day R,Wilson R.The ecology of polychaetes that infest abalone shells in Victoria,Australia[J].Journal of Shellfish Research,2004,23(4):1179-1188.
[5]Sato-Okoshi W.Polydorid species (Polychaeta : Spionidae) in Japan,with descriptions of morphology,ecology and burrow structure.1.Boring species [J].Journal of the Marine Biological Association of the United Kingdom,1999,79(5): 831-848.
[6]Boscolo R,Giovanardi O.Polydora ciliatashell infestation in Tapesphilippinarum Manilaclam held out of the substrate in the Adriatic sea,Italy [J].Journal of Invertebrate Pathology,2002,79(3): 197-198.
[7]Almeda R,Pedersen T M,Jakobsen H H,et al.Feeding and growth kinetics of the planktotrophic larvae of the spionid polychaetePolydora ciliata(Johnston) [J].Journal of Experimental Marine Biology and Ecology,2009,382(1): 61-68.
[8]崔秀林.扇贝养殖中黑壳病的发生和预防[J].河北渔业,1995,81: 16.
[9]刘慧玲.凿贝才女虫发育阶段的形态观察[J].湛江海洋大学学报,2003,23(6): 8-11.
[10]王爱民,石耀华,吴星.4种防治马氏珠母贝多毛类寄生病方法的效果比较[J].海洋水产研究,2004,25(2): 41-46.
[11]宋贞坪,李霞,王福景,等.双齿围沙蚕主要器官组织学的研究[J].大连海洋大学学报,2010,25(4):320-323.
[12]Westheide W.Ultrastructure of the genital organs in interstitial polychaetes.III.Penes and ejaculatory ducts inHesionides arenaria(Hesionidae)[J].Helgoland Marine Research,1982,35(4): 479-488.
[13]Tzetlin A B,Dahlgren T,Purschke G.Ultrastructure of the body wall,body cavity,nephridia and spermatozoa in four species of the Chrysopetalidae (Annelida,“Polychaeta”) [J].Zoologischer Anzeiger,2002,241:37-55.
[14]Sato-Okoshi W,Okoshi K.Microstructure of scallop and oyster shells infested with boringPolydora[J].Nippon Suisan Gakkaishi,1993,59(7): 1 243-1 247.
[15]Haigler Sarah A.Boring Mechanism ofPolydora websteriinhabitingCrassostrea virginica[J].American Zoologist,1969,9: 821-828.
[16]张子慧.动物的肌肉[J].生物学通报2004,39(8): 20.
[17]Miller D C,Jumars P A.Pellet accumulation,sediment supply,and crowding as determinants of surface deposit-feeding rate inPseudopolydora KempiJaponica Imajima & Hartman (Polychaeta,Spionidae)[J].Journal of Experimental Marine Biology and Ecology,1986,99(1): 1-17.
[18]Grau A,Crespo S,Sarasquete M C,et al.The digestive tract of the amberjackSeriola dumerilli,Risso: a light and scanning electron microscope study [J].Journal of Fish Biology,1992,41(2): 287-303.
猜你喜欢
自然杂志(2022年3期)2022-08-18
医学研究生学报(2021年4期)2021-12-02
小读者(2021年2期)2021-03-29
学生天地(2020年9期)2020-08-25
新农民(2020年15期)2020-06-22
中华皮肤科杂志(2019年5期)2019-06-24
浙江理工大学学报(自然科学版)(2016年5期)2016-09-15
小主人报(2016年4期)2016-02-28
中国环境科学(2015年7期)2015-08-30
天津医药(2011年1期)2011-03-16
