
长江中下游地区湖北钉螺线粒体CO I基因遗传变异研究
2011-03-14 06:15李洪军曲国立邢云天李幼子魏剑英戴建荣梁幼生
海洋科学 2011年10期
李洪军,汪 伟,张 伟,曲国立,邢云天,李幼子,魏剑英,戴建荣,梁幼生
(1.江苏省血吸虫病防治研究所,江苏 无锡 214064; 2.卫生部 寄生虫病预防与控制技术重点实验室,江苏无锡 214064)
血吸虫病是一种严重危害人类健康、影响社会经济发展的“被忽略”的热带病(Neglected Tropical Diseases)[1],全球有76个国家和地区流行血吸虫病,感染者超过2亿,近8亿人口面临感染威胁[2]。在我国,日本血吸虫病仍然是重要的公共卫生问题[3-5]。截至2010年底,全国仍有血吸虫病患者约51.6万[6]。经过多年积极防治,我国的血吸虫病流行区主要被压缩在长江中、下游沿岸5省(湖南、湖北、江西、安徽和江苏)的江湖洲滩地区[7-10]。钉螺是日本血吸虫的唯一中间宿主,是构成血吸虫病传播的不可缺失环节[11]。研究表明,凡有血吸虫病流行的地区必有钉螺孳生[12]。控制与消灭钉螺是阻断血吸虫病传播的有效措施之一[13-14]。
既往已有采用现代分子生物学技术对中国大陆钉螺系统进化进行研究的报道[15-18],亦有学者对湖北省庙河地区和长江三峡库区钉螺线粒体基因遗传变异进行了探讨[19-20]。本研究通过对长江中、下游血吸虫病流行区江苏、安徽、江西、浙江和湖北 5省8个地理株湖北钉螺线粒体DNA细胞色素C氧化酶亚单位1(CO I)基因进行遗传多态性分析,探讨其核酸特异性,为进行长江中、下游地区钉螺遗传变异研究、建立DNA 指纹检测平台及钉螺分子鉴定提供参考依据。
1 材料与方法
1.1 材料
1.1.1 钉螺样本采集
分别采自长江中、下游地区的江苏、湖北、安徽、江西、浙江4个省的8个流行区现场钉螺,钉螺采集地分别为江苏丹徒和邗江、湖北武昌、安徽安庆和贵池、江西彭泽和余干、浙江平湖。所有钉螺均为肋壳钉螺,系湖北钉螺指名亚种(Oncomelania hupensis hupensis)。在实验室内适应性饲养后,采用群体逸蚴法鉴定钉螺是否感染血吸虫,选择阴性成螺备用。
1.1.2 主要仪器与试剂
PCR仪为德国analyticjena公司产品,台式离心机为德国Eppendorf 公司产品; 基因组DNA抽提试剂盒购自 promega公司,Taq DNA 聚合酶购自promega公司。
1.2 方法
1.2.1 钉螺基因组DNA的提取
选择每一地理株钉螺3~5只,彻底洗净后,去除螺壳及内脏肠管组织,将剩余组织进行混合,移入 EP管中,加入蛋白激酶 K,混匀,55 ℃水浴过夜。用基因组 DNA抽提试剂盒提取钉螺基因组DNA,并在核酸蛋白分析仪上测定 DNA浓度和纯度。
1.2.2 PCR扩增及产物鉴定、测序
上游引物 P1:5′-GGTCAACAAATCATAAAGATATTGG-3′,下游引物 P2:5′ -TAAACTTCAGGGTGACCAAAAAAYCA-3′,由上海生物工程有限公司合成。50 μL 反应体系为:10×reaction buffer 5.0 μL,25mmol/L MgCl25.0 μL,dNTP 2.0 μL,上、下游引物各 2.0 μL,模板 DNA 1.0 μL,Taq DNA聚合酶 1.0 μL,用ddH2O补足反应体系。反应循环条件为:94 ℃预变性10 min,94 ℃变性1 min,55 ℃退火90 s(COI),72 ℃延伸1 min,共35个循环,末次循环72℃延伸10 min。扩增产物经1.5%琼脂糖凝胶电泳鉴定,并将扩增产物送上海英骏生物技术公司测序。
1.2.3 CO I 基因序列分析
将测序结果先用ClustalX 1.81软件进行多序列比对,再在 MEGA 4.0软件中的 Kimura 双参数法(Kimura 2-Parameter)计算遗传距离,分别用邻接法(NJ)和最大简约法(MP) 构建系统发生树,Bootstrap进行检验。
2 结果
2.1 CO I 基因PCR 扩增结果
以 8个不同地理株湖北钉螺基因组 DNA 为模板,PCR 特异性扩增线粒体DNA CO I 基因,均在长度约700 bp 附近均扩增出一目的条带,与预期目的片段大小一致。
2.2 CO I基因序列分析
经过 ClustalX 1.81 软件比对,删除多变区后,得到701个同源位点,其中有保守位点663个,可变位点38个,简约信息位点26个。A、T、C和G的碱基平均含量分别为22.8%、37.6%、18.8%、20.8%,A+T平均含量为 60.4%,C+G平均含量为 39.6%,A/T含量明显偏高。表1给出了基于Kimura双参数模型序列分歧矩阵。最终得出所有序列间的平均遗传距离为0.025(表1)。

表1 本研究中不同地理株间的遗传距离(基于Kimura双参数模型)Tab.1 The genetic distances based on the formula of Kimura 2 parameters among snails from different areas in this study
2.3 基于CO I基因序列的分子系统树
本研究采用邻接法和最大简约法共构建了 2种分子系统树:NJ树和 MP树。NJ树中的分支检验置信值和后验值低于 50%的则未显示。NJ进化树显示,江苏丹徒、湖北武昌、江西余干及安徽安庆和贵池5个地理株的钉螺形成一个分支,而浙江平湖、江苏邗江和江西彭泽3个地理株钉螺形成另一个分支; 但两种方法构建的系统发生树存在差异。在内部分支检验置信值在大于 80%时很相近(图 1、图 2)。

图1 基于CO I基因部分序列构建的NJ树(各分支上数字为1 000次内部分支检验置信值)Fig.1 The NJ tree based on CO I gene sequence data(Numbers on each node correspond to their bootstrap values for 1000 replicates.)
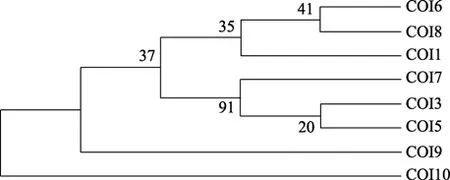
图2 基于CO I基因部分序列构建的MP树各分支上的数字为bootstrap1 000个循环的自举检验值Fig.2 The MP tree based on partial of CO I gene sequences data with confident values of Bootstrap 1 000 indicated above each branch
3 讨论
随着分子生物学技术的迅速发展,线粒体DNA、核糖体DNA和RNA、微卫星DNA、单核苷酸多态性(SNP)和限制性片段长度多态性(RFLP)、随机扩增多态性(RAPD)等多种分子遗传研究方法和遗传标记的出现,为湖北钉螺遗传多态性及其分子系统学的研究提供了更丰富的手段[21-22]。线粒体DNA作为一种核外遗传物质,具有以母性遗传为主,进化速率快,较易发生突变,其突变频率约为核基因组的 5~10 倍,且基因重组率极低,同一个体组织中的线粒体 DNA 具有一致性等特点,使它成为生物进化过程中谱系发生和迁移流动的有效遗传标记,已经广泛应用于寄生虫属、种及种群水平相互关系的研究[19,23-26]。
本研究通过对长江中、下游地区 8个不同地理株湖北钉螺线粒体 DNA CO I 基因进行序列分析,从分子水平上探明其种群内的遗传差异。研究发现,不同地域株湖北钉螺遗传差异显著,平均遗传距离为 0.025。进化树显示,江苏丹徒、湖北武昌、江西余干及安徽安庆和贵池 5个地理株钉螺形成一个分支,而浙江平湖、江苏邗江和江西彭泽3个地域钉螺形成另一个分支。这可能与长江中、下游地区形成的天然地理隔离和生态环境差异有关,支持 Davis等[27]对湖北钉螺的分类结果。Wilke等[17]对钉螺cox1基因序列研究发现,长江中下游地区光壳和肋壳钉螺之间遗传变异很小,同属指名亚种。石朝辉等[28]发现同一地区光壳与肋壳钉螺 cox1 基因序列之间有较高的同源性。本实验结果表明,长江中、下游地区湖北钉螺种群内存在较高的核苷酸多态性,遗传差异较显著,且两种方法构建的进化树存在差异。由于线粒体涉及绝大多数真核生物的呼吸代谢,线粒体DNA 含有12 或13 种呼吸链酶原复合物中重要蛋白的编码基因,具有丰富的遗传特征,在生物进化过程中受到的选择压力较大。因此,线粒体 DNA序列的进化要快于细胞核基因[29]。这一遗传差异的原因及其会否导致螺宿主对日本血吸虫易感性差异有待于进一步研究。
本研究采用线粒体DNA CO I基因分子标记获得了长江中、下游血吸虫病流行区湖北钉螺种群CO I基因序列差异,为进一步构建中国大陆湖北钉螺遗传多态性数据库和DNA 指纹检测平台奠定了基础。建立的分子技术可对现场采集到的钉螺进行地理种株和亲缘关系鉴定。
[1]O’Connell D.Neglected Diseases[J].Nature,2007,449:157-182.
[2]Steinmann P,Keiser J,Bos R,et al.Schistosomiasis and water resources development:systematic review,meta-analysis,and estimates of people at risk[J].Lancet Infect Dis,2006,6:411-425.
[3]周晓农,姜庆五,汪天平,等.我国血吸虫病防治研究现状与发展战略思考[J].中国血吸虫病防治杂志,2004,17(1):1-3.
[4]Zhou X N,Guo J G,Wu X H,et al.Epidemiology of schistosomiasis in the People’s Republic of China,2004[J].Emerg Infect Dis,2007,13:1470-1476.
[5]Wang L D,Utzinger J,Zhou X N.Schistosomiasis control:experiences and lessons from China[J].Lancet,2008,372:1793-1795.
[6]郝阳,郑浩,朱蓉,等.2009年全国血吸虫病疫情通报[J].中国血吸虫病防治杂志,2010,22(6):521-527.
[7]Li S Z,Luz A,Wang X H,et al.Schistosomiasis in China:acute infections during 2005-2008[J].Chin Med J (Engl),2009,122:1009-1014.
[8]郝阳,易冬华,张险峰,等.2008年全国血吸虫病疫情控制考核评估报告[J].中国血吸虫病防治杂志,2009,21(6):457-463.
[9]Zhao G M,Zhao Q,Jiang QW,et al.Surveillance for schistosomiasis japonica in China from 2000 to 2003[J].Acta Trop,2005,96:288-295.
[10]朱蓉,党辉,张利娟,等.2005-2008年全国血吸虫病疫情监测[J].中国血吸虫病防治杂志,2009,21(5):358-362.
[11]中华人民共和国卫生部疾病控制司.血吸虫病防治手册[M].3版.上海:上海科学技术出版社,2000:23.
[12]袁鸿昌,张绍基,姜庆五.血吸虫病防治理论与实践[M].上海:复旦大学出版社,2003:3-4.
[13]郭家钢.中国血吸虫病综合治理的历史与现状[J].中华预防医学杂志,2006,40(4):225-228.
[14]Yuan Y,Xu X J,Dong H F,et al.Transmission control of schistosmiasis japonica:implementation and evaluation of different snail control interventions[J].Acta Trop,2005,96:191-197.
[15]刘月英,楼子康,王耀先,等.钉螺亚种分化[J].动物分类学报,1981,6(3):253-266.
[16]周晓农,孙乐平,洪青标,等.中国大陆钉螺种群遗传学研究 I 种群遗传差异[J].中国血吸虫病防治杂志,1995,7(2):67-71.
[17]Wilker T,Davis G M,Cui E C,et al.Oncomelania hupensis(Gastropoda:Rissooidea) in eastern China:molecular phylogeny,population structure,and ecology[J].Acta Trop,2000,77:215-227.
[18]周艺彪,赵根明,韦建国,等.湖北钉螺种群内AFLP分子标记遗传变异分析[J].中国寄生虫学与寄生虫病杂志,2006,24(1):27-31.
[19]Shi C H,Wilke T,Davis G M,et al.Population genetics,microphylogeography,ecology and susceptibility to schistosome infection of ChineseOncomelania hupensis hupensis(Gastropoda:Rissooidea:Pomatiopsidae)in the Miao River System[J].Malacologia,2002,44:333-338.
[20]陈琳,张锡林,何谐,等.三峡库区上、下游血吸虫病流行区钉螺线粒体cox1基因遗传变异研究[J].热带医学杂志,2010,10(1):5-8.
[21]周艺彪,姜庆五,赵根明.湖北钉螺遗传多样性及其分子系统学研究进展[J].中国血吸虫病防治杂志,2005,17(5):391-396.
[22]孙恩涛,张锡林.湖北钉螺遗传标记研究进展[J].中国血吸虫病防治杂志,2010,22(1):90-95.
[23]Moro P L,Nakao M,Ito A,et al.Molecular identification ofEchinococcusisolates from Peru[J].Parasitol Int,2009,58:184-186.
[24]Nguyen T G,Van De N,Vercruysse J,et al.Genotypic characterization and species identification ofFasciolaspp.with implications regarding the isolates infecting goats in Vietnam[J].Exp Parasitol,2009,123:354-361.
[25]Jeon H K,Kim K H,Huh S,et al.Morphologic and genetic identification of Diphyllobothrium nihonkaiense in Korea[J].Korean J Parasitol,2009,47:369-375.
[26]Semyenova S K,Morozova E V,Chrisanfova G G,et al.Genetic differentiation in eastern European and western Asian populations of the liver fluke,Fasciola hepatica,as revealed by mitochondrial nad1 and cox1 genes[J].J Parasitol,2006,92:525-530.
[27]Davis G M,Wilke T,Zhang Yi,et al.Snails-Schistosoma,paragonimus interactions in China:population ecology,genetic diversity,coevolution and emerging diseases[J].Malacologia,1999,41:355-377.
[28]石朝辉,邱持平,夏明仪,等.湖北省庙河地区钉螺细胞色素C 氧化酶1 基因差异的研究[J].中国寄生虫学与寄生虫病杂志,2001,19(1):41-44.
[29]Le T H,Blair D,McManus D P.Mitochondrial genomes of parasitic flatworms [J].Trends Parasitol,2002,18:206-213.
猜你喜欢
热带病与寄生虫学(2022年4期)2022-09-20
世界科学技术-中医药现代化(2022年3期)2022-08-22
热带病与寄生虫学(2022年6期)2022-02-11
现代畜牧科技(2021年2期)2021-03-19
湖北大学学报(自然科学版)(2021年1期)2021-01-06
湖南畜牧兽医(2016年3期)2016-06-05
兽医导刊(2016年6期)2016-05-17
西南农业学报(2016年6期)2016-04-16
河南畜牧兽医(2015年13期)2015-11-28
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
