
大菱鲆不同家系生长性能的比较
2011-03-14 06:06王新安马爱军雷霁霖曲江波黄智慧薛宝贵
海洋科学 2011年4期
王新安,马爱军,雷霁霖,杨 志,曲江波,黄智慧,薛宝贵,3
(1.中国水产科学研究院黄海水产研究所 青岛市海水鱼类种子工程与生物技术重点实验室,山东 青岛266071;2.烟台市开发区天源水产有限公司,山东 烟台 264003;3.上海海洋大学 上海 201306)
大菱鲆(Scophthalmus maximusL.)为原产于欧洲的著名海水养殖特有良种,具有生长迅速、肉味鲜美、经济价值高等优点,自然分布于大西洋东北部,北起冰岛,南至摩洛哥附近的欧洲沿海[1-2]。中国水产科学研究院黄海水产研究所于1992年“跨洋引种”将其引入中国,突破苗种繁育的关键技术,并创建了“温室大棚+深井海水”的开放式工厂化养殖模式,使其成为中国北方沿海工厂化养殖业的主导品种之一[3-4]。近年来,由于引进原种数量较少,群体种类相对单一,并且育种过程中采用的亲鱼未经过专门选优处理,加之累代养殖和近亲交配,以致造成种质退化现象比较严重,结果导致孵化率、成活率降低、生长速度减慢、抗逆性差、白化严重等系列种质退化现象不断发生[5-7]。因此,有必要对大菱鲆进行遗传改良,以选育出具有生长快、抗逆性强等性状的新品种(系),为大菱鲆养殖业健康、可持续发展提供重要保证。
基于当前国际上运用大规模家系选育技术在水产养殖动物进行遗传改良已取得重大成效[8-10],运用家系选育技术对大菱鲆进行遗传改良,同样具有重要意义。本课题组选取不同地理群体的大菱鲆,于2007年4月,在烟台天源水产有限公司根据巢式设计已完成家系的构建和培育[5-11]。关于水产动物生长性能的比较研究,国内外文献报道较多,如李思发[12-13]、Eknath[14]、罗相忠[15]、文春根[16]、王炳谦[17]等分别对不同品系或种群的鲢(Hypophthalnichthys molitrix)、罗非鱼(Tilapia niloticaLinnaeus)、鲟鱼(Acipenser gueldenstaedtiBrandt)、虹鳟(Oncorhynchus mykiss)等的生长性能进行了比较研究,Overturf[18]等对几个品系虹鳟的生长性能、遗传杂合度等进行了比较。有关大菱鲆不同群体后代生长性能的研究已有报道,于飞等[19]比较了群体间家系的生长性能。但其构建家系的基础群体与本研究所用的基础群体并不完全相同,即所研究的家系是不同的,其生长性能也必然存在差异。本试验通过对所构建家系生长性能的比较,分析群体间及群体内家系的生长性能的差异,为大菱鲆家系选育提供基础数据和理论依据。
1 材料与方法
1.1 亲本来源
实验地点在烟台天源水产有限公司。实验亲本为烟台天源水产有限公司所培育的英国(E)、法国(F)、丹麦(D)和挪威(N)4个不同地理群体的大菱鲆,其中英国群体为2002年8月11日从英国进海捕亲本孵化成的约5cm的苗种;法国群体为2003年9月22日从法国进海捕亲本孵化成的约5cm的苗种;丹麦群体为2003年4月4日从丹麦进口海捕亲本孵化而成的约5cm的苗种;挪威群体为2002年6月10日从挪威进海捕亲本孵化成的约5cm的苗种。
1.2 人工受精及孵化
各家系构建于2007年4月12日至7月19日在烟台天源水产有限公司进行。选取体格健壮、体形完整、色泽正常、性腺发育成熟、状态良好大菱鲆作为亲鱼,在群体内和群体间,根据巢式设计,按照1雄配2雌的原则,在产卵盛期,采用人工采卵受精的方法进行定向交配。受精卵放置在0.8 m×0.6 m×0.6 m长方形孵化箱(80~100目)内孵化,孵化温度13~14℃。实验采用了12种群体间组合及4种群体内组合的 32个全同胞家系进行实验研究(见表2)。每一群体选择10尾雄鱼、20尾雌鱼作为备选亲本提前 1个月进行体外液氮金属数字标记,以避免亲本重复使用。
1.3 苗种培育
从仔鱼孵化后到标记混养前,各家系分池培育,为尽量减小环境条件的差异,对家系个体采用环境条件标准化和数量标准化的方式进行培育。
1.3.1 环境条件标准化
从仔鱼孵化后到标记混养前这一阶段,尽量使每个家系在各阶段的培育条件保持一致。主要包括各阶段养殖空间的大小、水的盐度、水温、光照、充气和饵料等条件。按照不同发育阶段,各家系可放置在同一车间养殖管理。
1.3.2 数量标准化
在大菱鲆早期发育的不同阶段,为消除放养密度的影响,对其进行 3次标准化处理(第一次标准化后,仍会有一定数量的苗种死亡,使放养密度发生变化,因此,需要进行第二次、第三次数量标准化)。依据本试验点育苗车间培育池的规格,每次标准化的数量标准如下:
第1次数量标准化:即分苗一期标准化,孵化后第15天,选取生长状态良好的仔鱼10 000尾放置在4 m×1.5 m×1 m的水泥池内培育。
第2次数量标准化:即分苗二期标准化,孵化后第30天,选取生长状态良好的稚鱼5 000尾放置在4 m×1.5 m×1 m的水泥池内培育。
第3次数量标准化:即分苗三期标准化,孵化后第60天,选取最大等级且生长状态良好的幼鱼2 000尾放置在5 m×3 m×1 m的水泥池内进行中间培育。中间培育约需 1个月后,对幼鱼进行荧光(VIE)标记,进行家系混养,以消除环境因素对各交配组合所造成的影响。
1.4 数据测量
从仔鱼孵化后开始,每隔 3个月进行一次鱼体质量测量(测量时同时进行混养池倒池),每个全同胞家系随机取样45尾。本实验采用数据为3月龄至12月龄的体质量。
1.5 统计分析方法
本文数据用SPSS13.0软件及Excel软件对试验数据进行处理。对不同生长阶段全同胞家系间的体质量差异进行方差分析和Duncan多重比较分析,差异的显著性设置为P<0.05。对不同生长阶段全同胞家系进行瞬时体质量增长率和绝对体质量增长率的计算。对不同生长时期家系的体质量以及不同生长阶段家系的生长速度进行相关性分析。
生长率的计算公式[20]:

式中,W1,W2分别为时间t1和t2时的体质量;RIGW,RAGW分别表示瞬时体质量增长率、绝对体质量增长率。
相关系数的计算公式为:
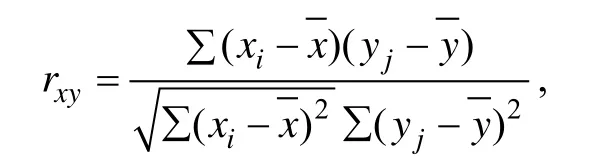
式中,xi,yj为不同生长时期每一家系的体质量或不同生长阶段每一家系的生长速度,,为相对应所有家系指标的平均值,i≠j,且i=3,6,9,12;j=3,6,9,12。
2 结果与分析
2.1 不同生长时期家系间体质量的差异分析
对不同生长时期各家系体质量进行方差分析的结果表明(表1),在3月龄、6月龄、9月龄和12月龄,家系间体质量差异均较大,F检验均为极显著水平,4个生长时期家系间生长的一致性均较差。进一步进行多重比较分析结果表明(表2),3月龄:平均体质量较高的家系F♂1×F♀3,F♂1×F♀4 和F♂2×E♀3,与其他家系均达到差异显著水平(P<0.05),平均体质量较低的是 N♂1×N♀8,N♂2×E♀8,N♂1×N♀7三个家系,平均体质量最高的家系比最低的家系高出约478.12%;6月龄:平均体质量较高的家系是F♂1×F♀3,E♂1×E♀2,E♂4×N♀2,F♂4×N♀3,与其他大多数家系差异显著(P<0.05),平均体质量较低的是N♂2×E♀8,N♂1×N♀7,D♂4×N♀6,N♂4×D♀7,N♂4×D♀8五个家系,平均体质量最高的家系比最低的家系高出约167.10%;9月龄:平均体质量较高的家系是E♂1×E♀2,F♂4×N♀3,F♂1×F♀4,F♂2×E♀3,与其他大多数家系差异显著(P<0.05),平均体质量较低的家系是N♂2×E♀8,N♂1×N♀7,D♂4×N♀6,N♂4×D♀7,平均体质量最高的家系比最低的家系高出约185.24%;12月龄:平均体质量较高的家系是 E♂1×E♀2,F♂4×N♀3,F♂2×E♀4,E♂2×F♀1,与其他大多数家系差异显著(P<0.05),平均体质量较低的家系是N♂2×E♀8,N♂1×N♀7,N♂4×D♀7,N♂3×F♀7,D♂4×N♀6,F♂3×D♀4,平均体质量最高的家系比最低的家系高出约168.62%。通过上述结果发现,家系在 3,6,9,12月龄体质量排序上存在不一致性,为此对 4个生长时期的体质量进行了相关分析,结果表明(表3),除3月龄体质量和6月龄体质量间的相关系数显著性检验达到显著水平外(P<0.05),其他月龄间的相关系数的显著性检验均达到了极其显著水平(P<0.01),显然,在3,6,9,12月龄,尽管各家系在体质量排序上并不完全一致,但对于大多数家系而言,家系的初始平均体质量对试验终值体质量仍然有较大的影响。由表3还可以看出,在 1龄鱼阶段内,生长时期越长,家系的体质量对下一生长时期家系体质量的影响越大,相邻两个时期家系排序的一致性越好。
2.2 不同生长阶段家系的生长速度分析
大菱鲆不同生长阶段家系的生长速度分析见(表4,图1)。结果表明,在3~6月龄,绝对体质量增长率较高的家系是 E♂1×E♀2,E♂4×N♀2,F♂1×F♀3,F♂4×N♀3,F♂4×N♀4,绝对体质量增长率较低的家系是N♂2×E♀8,N♂1×N♀7,D♂4×N♀6;在6~9月龄,绝对体质量增长率较高的家系是E♂1×E♀2,F♂4×N♀3,F♂2×E♀3,F♂1×F♀4,绝对体质量增长率较低的家系是 D♂2×E♀6,N♂1×N♀7,N♂2×E♀8,D♂3×F♀5;在9~12月龄,绝对体质量增长率较高的家系是 E♂2×F♀1,E♂1×E♀2,F♂2×E♀4,F♂4×N♀3,绝对体质量增长率较低的家系是N♂3×F♀7,N♂1×N♀7,N♂2×E♀8,F♂3×D♀4。上述三个不同生长阶段,绝对体质量增长率最高的家系比绝对体质量增长率最低的家系分别快 163.16%,209.52%,176.20%。从表4中还可以看出,在每一生长阶段,家系的瞬时体质量增长率的排序与绝对体质量增长率并不一致,显然,使用绝对体质量增长率和瞬时体质量增长率来表达鱼体的生长快慢有时会产生不一致的结果,但这两种方法的同时使用和综合分析有助于得出较为准确的结论。通过表4还发现,家系在3~6月龄、6~9月龄及9~12月龄绝对体质量增长率排序上存在不一致性,为此对 3个生长阶段的生长速度(绝对体质量增长率)进行了相关分析(表5),结果表明,3~6月龄生长速度与 6~9月龄和 9~12月龄生长速度的相关系数分别为 0.392和 0.390,相关系数的显著性检验均达到显著水平(P<0.05),6~9月龄与9~12月龄生长速度的相关系数为0.887,相关系数的显著性检验达到极显著水平(P<0.01)。进一步进行3~6月、6~9月龄生长速度对9~12月龄生长速度影响的通径分析(表6),结果表明,6~9月龄生长速度的通径系数具有统计学意义(P=0.000<0.05),对9~12月龄生长速度的效应大,综合决定系数为0.759,3~6月生长速度的通径系数不具有统计学意义(P=0.596>0.05),对9~12月龄生长速度的效应小,综合决定系数仅为 0.005,因此,3~6月龄生长速度对 9~12月龄生长速度的效应可以忽略不计。从表6还可以看出,3~6月龄生长速度和9~12月龄生长速度的方差膨胀因子都没有超过经验值(方差膨胀因子,VIF<10),表明模型中的复共线性程度很小,拟合的线性统计模型及相应参数的估计可靠。显然,尽管家系在 3~6月龄、6~9月龄及 9~12月龄的绝对体质量增长率在排序上存在不一致性,但对于大多数家系而言,不同生长阶段家系的生长速度仍然具有一定的关系,其中6~9月龄家系的生长速度对 9~12月龄家系的生长速度的影响较 3~6月龄家系的生长速度对6~9月龄家系的生长速度大,9~12月龄家系的生长速度主要取决于6~9月龄家系的生长速度,即9~12月龄家系生长速度与6~9月龄家系生长速度在排序上的一致性比6~9月龄家系生长速度与3~6月龄家系生长速度在排序上的一致性好得多。

表1 大菱鲆32个全同胞家系组合不同生长阶段的体质量方差分析Tab.1 Variance analysis of body weight among 32 full-sib families combinations at different stages

表2 大菱鲆不同生长阶段全同胞家系体质量平均数的Duncan多重比较Tab.2 Duncan multiple range test for comparisons of multi-average of body weight at different stages

表3 大菱鲆不同生长阶段家系体质量间的相关系数Tab.3 Correlation coefficients of body weight of families of Scophthalmus maximus at different growth stages
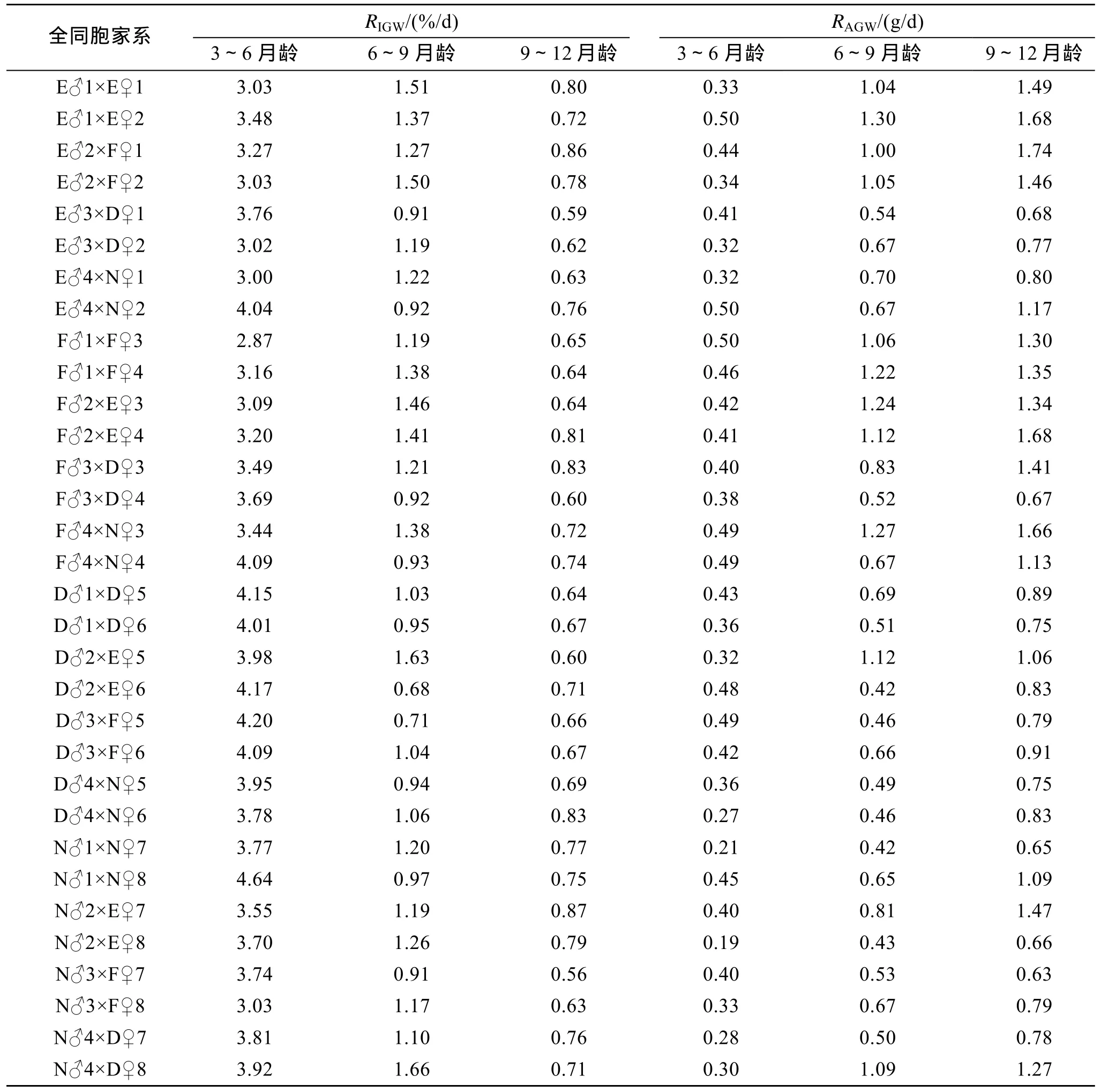
表4 大菱鲆不同生长阶段32个全同胞家系的体质量增长率Tab.4 Growth rates of body weight of 32 families of Scophthalmus maximus at different stages

图1 各月龄大菱鲆不同家系生长速度离差Fig.1 Residuals of growing rate of different full-sib families of scophthalmus maximus of 3~6 (above),6~9 (middle) and 9~12(below) months

表5 大菱鲆不同生长阶段家系生长速度间的相关系数Tab.5 Correlation coefficients of growing rate of families of Scophthalmus maximus at different growth stages
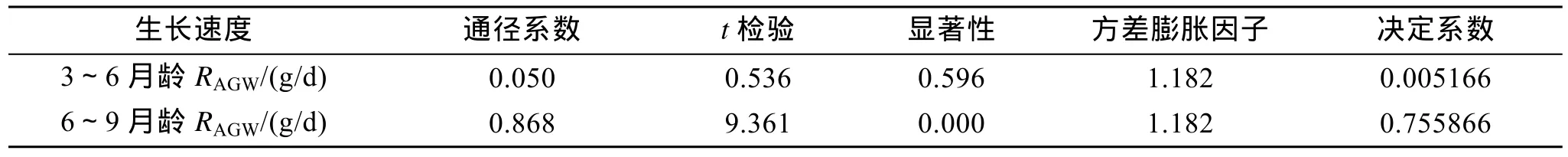
表6 3~6月龄、6~9月龄生长速度对9~12月龄生长速度影响的通径分析Tab.6 Path analysis of RAGW/(g/d) of 3~6 and 6~9 months on that of 9~12 months
3 讨论
本文对来自不同地理群体大菱鲆亲鱼的群体间家系和群体内家系进行了生长性能的比较和分析。不同生长时期家系体质量的差异分析和不同生长阶段家系的生长速度分析均表明,家系生长性能的排序存在不一致性,生长时期(生长阶段)越长,相邻两个生长时期(生长阶段)家系生长性能的相关性越高,家系排序的一致性越好。综合不同生长时期家系的体质量差异分析和不同生长阶段家系的生长速度分析,在1龄鱼阶段内,从整体上看来自英国群体和法国群体家系的生长性能比丹麦群体和挪威群体家系的生长性能更为优良。这表明在实际生产中应用法国群体和英国群体进行群体间和群体内交配进行繁育,可取得相对于应用丹麦群体和挪威群体较高的经济效益。然而,尽管丹麦群体和挪威群体群体内家系的生长性能较弱,但部分英国群体和法国群体与丹麦群体与挪威群体的群体间家系存在着较为优良的生长性能,如F♂4×N♀3、N♂2×E♀7等杂交家系,这可能是群体间杂种优势的体现。有关大菱鲆群体间杂种优势的研究已有报道[21],尽管未涉及本文所采用的大菱鲆群体,作者仍然认为英国、法国群体与丹麦、挪威群体间部分家系存在的优良生长性能,是不同群体间杂交优势的结果。不同生长时期(生长阶段)家系生长性能排序的不一致性,表明培育生长性能稳定的大菱鲆品种,实现大菱鲆种质改良,具有极其重要的经济和理论意义。不同生长时期(生长阶段)家系生长性能均有较大的差异,表明群体生长性状遗传变异丰富,这有利于富集优良的生长基因,从而为运用选择育种技术对大菱鲆进行遗传改良提供了重要保证。
大菱鲆群体间和群体内家系在早期的构建和培育过程中,家系未标记混养前,各家系分池培育,为消除环境条件的差异,对这一生长阶段的家系个体采用标准化(包括环境条件标准化和数量标准化)的方式进行培育,然后进入标记混养阶段。因此,大菱鲆不同生长时期(生长阶段)家系生长性能的差异及大多数家系排序的不一致性,主要应由家系自身的遗传基础决定,受环境因素的影响较小。由于大菱鲆不同生长时期体质量的相关系数中,9月龄和12月龄之间的相关性最强,相关系数为 0.965,不同生长阶段生长速度的相关系数中,6~9月龄和 9~12月龄的相关性最强,相关系数为 0.887,决定系数为 0.756,尽管这两个生长时期(生长阶段)大多数家系生长性能的排序仍然不一致,但相对于其他生长时期(生长阶段)家系生长性能排序的一致性较好。因此,在采用选择育种技术对大菱鲆进行遗传改良时,进行个体选择的时期应不低于9月龄,否则,选择的效率和可信度会十分有限。于飞等(2008)对其他大菱鲆不同群体杂交后代25 d和80 d的生长差异研究发现[19],个体的生长速度差异较大,不适合于早期选择,这与本文的研究相一致。本研究进一步发现,在对大菱鲆选择育种时,进行个体选择的时期不应低于9月龄。至于最为适合的选择时间点,有待于进一步研究。
[1]雷霁霖.英国养殖大菱鲆简况[J].水产科技情报,1983,2:26-27.
[2]雷霁霖.大菱鲆养殖技术[M].上海:上海科学技术出版社,2003.
[3]雷霁霖,门强,王印庚,等.大菱鲆“温室大棚+深井海水”工厂化养殖模式[J].海洋水产研究,2002,23(4):1-7.
[4]马爱军,雷霁霖,陈四清,等.大菱鲆不同产卵季节对卵子的生物学及生化特征的影响[J].海洋与湖沼,2002,33(1):75-82.
[5]马爱军,王新安,雷霁霖,等.大菱鲆(Scophthalmus maximus)四个不同地理群体数量形态特征比较[J].海洋与湖沼,2008,39(1):24-29.
[6]申雪艳,宫庆礼,雷霁霖,等.进口大菱鲆Scophthalmus maximusL.苗种的遗传结构分析[J].海洋与湖沼,2004,35(4):332-341.
[7]雷霁霖,马爱军,陈超,等.大菱鲆(Scophthalmus maximus)养殖现状与可持续发展[J].中国工程科学,2005,7(5):30-34.
[8]Gjerde B,Korsvoll A.Realised selection differentials forgrowth rate and early sexual maturity in Atlantic salmon[J].Aquac Eur,1999,99:73-74.
[9]李鸿鸣,孙效文.应用大规模家系选育技术促进辽宁海水养殖业的可持续发展[J].沈阳农业大学学报,2002,4(1):7-10.
[10]Gall G A E,Bakar Y.Application of mixed-model techniques to fish breed improvement:analysis of breeding-value selection to increase 98-day body weight in tilapia[J].Aquaculture,2002,212:93-113.
[11]王新安,马爱军,雷霁霖,等.大菱鲆早期阶段外部形态性状对体质量的影响效果分析 [J].动物学报,2008,54(3):540-545.
[12]李思发,蔡正纬,陆伟民,等.长江水系鲢鱼和珠江水系鲢鱼的生长差异[J].水产学报,1984,8(3):211-217.
[13]李思发,李晨虹,李家乐,等.尼罗罗非鱼五品系生长性能评估[J].水产学报,1998,22(4):314-321.
[14]Eknath A E,Tayamen M M,Palada-de Vera M S,et al.Genetic improvement of farmed tilapias:the growth performance of eight strains ofOreochromis niloticustested in different farm environments[J].Aquaculture,1993,111:171-188.
[15]罗相忠,邹桂伟.雌核发育鲢各系及与普通鲢生长的比较研究[J].集美大学学报,2003,98(3):203-207.
[16]文春根.俄罗斯鲟与杂交鲟鱼种的生长对比试验[J].水利渔业,2004,24(6):27-29.
[17]王炳谦,谷伟,贾钟贺,等.4个品系虹鳟生产性能的比较[J].大连水产学院学报,2007,22(3):170-174.
[18]Overturf K,Casten M T,Lapatra S E,et al.Comparison of growth performance,immunological response and genetic diversity of five strains of rainbow trout (Oncorhynchus mykiss) [J].Aquaculture,2003,217:93-106.
[19]于飞,张庆文,孔杰,等.大菱鲆不同进口群体杂交后代的早期生长差异[J].水产学报,2008,32(1):58-64.
[20]李思发.淡水鱼类种群生态学[M].北京:中国农业出版社,1990.
[21]季士治,雷霁霖,王伟继,等.双列杂交法分析 2个大菱鲆养殖群体的杂交效果[J].中国水产科学,2006,13(6):1001-1005.
猜你喜欢
黑龙江水产(2022年6期)2022-12-03
科学生活(2019年7期)2020-01-01
中国水产(2019年3期)2019-03-25
养殖与饲料(2019年10期)2019-02-25
山东畜牧兽医(2018年3期)2018-04-26
浙江农业学报(2016年7期)2016-06-15
广东海洋大学学报(2015年4期)2016-01-13
听力学及言语疾病杂志(2015年5期)2015-12-24
听力学及言语疾病杂志(2015年5期)2015-12-24
首都医科大学学报(2015年4期)2015-12-16
