
桂花种子休眠和萌发的初步研究
2010-12-05 02:49杨秀莲郝其梅
浙江农林大学学报 2010年2期
杨秀莲,郝其梅
(1.南京林业大学 风景园林学院,江苏 南京 210037;2.江苏省新沂市建设局,江苏 新沂 221400)
种子休眠是指具有生活力的种子处于适宜的环境条件下仍不能正常萌发的一种生理现象。种子休眠是植物在长期演化过程中为了种的生存而不断适应环境的结果[1-2]。对植物本身来说,种子的休眠对植物个体的生存、物种的延续和进化具有积极的作用,但对林木的生产带来较大的影响。通过观察,桂花Osmanthus fragrans种子具有一定的休眠现象。当年4-5月成熟的种子需层积至翌年2-3月才能萌发。张义等[3]利用赤霉素浸种和低温层积相结合的方法进行桂花种子的催芽试验,证明桂花种子具有休眠特性,低温层积和赤霉素(GA3)浸种可有效促进种子的萌发。袁王俊等[4]利用离体培养技术对层积3个月的桂花种子进行胚培养,打破了种子的休眠。对于桂花种子的休眠原因,目前尚未见报道。本研究通过对桂花种子的休眠和萌发的研究,旨在探讨桂花种子的休眠机制,以期寻找解除休眠的最佳方法,缩短休眠时间,提高播种繁殖率,同时也为品种间杂交和培育新品种积累经验,提供理论指导。
1 材料与方法
1.1 试验材料
紫梗籽银桂Osmanthus fragrans‘Zigeng Ziyin’种子:2007年5月初采于南京林业大学教五楼前,去除外种皮后保存于(3±1)℃冰箱中。紫梗籽银桂为银桂品种群中的一个品种,结实量较大,种子采收容易,故作为桂花种子休眠的研究对象。
白菜Brassica campestris种子:南京市蔬菜种子公司的绿优1号白菜种子,纯度95%,净度98%,发芽率85%以上。
1.2 试验方法
1.2.1 种皮、种实浸提液的提取 取内种皮(以下称种皮)和种实(包含胚和胚乳两部分,下文同)各10 g,分别置于250 mL容量瓶中,加入200 mL体积分数为80.0%甲醇,混匀后放入冰箱内,在2~4℃的恒温条件下密闭浸提,期间多次取出摇匀使其充分浸提,48 h后过滤。所得滤液在35℃下减压浓缩后定容到0.10 g·mL-1(以1 mL浓缩液中含有原材料的量计算)。
1.2.2 白菜种子生物测定方法 在培养皿中放置滤纸,分别加入一定量的甲醇浸提原液(0.10 g·mL-1),60%原液(0.06 g·mL-1),30%原液(0.03 g·mL-1),设置 3 个重复·处理-1,100 粒·重复-1白菜种子,于30℃恒温全光照条件下培养,48 h测定发芽率,72 h测量苗高和根长,用含体积分数为0.2%甲醇的水溶液处理的白菜种子作对照。
1.2.3 催芽方法 采用蒸馏水(记为 G0)和质量浓度为 500 mg·L-1(记为 G5),1 000 mg·L-1(记为G10),1 500 mg·L-1(记为G15)的赤霉素溶液浸种48 h,后分别与湿沙按1∶3混合放入塑料盆中,上面覆盖保鲜膜并用剪刀剪几个小洞,利于种子呼吸,将塑料盆置于2~4℃冰箱中进行低温层积处理。层积期间隔半个月取各处理种子4个重复,30粒·重复-1。在25℃恒温、24 h光照条件下做种子发芽试验,并统计种子的发芽率。
2 结果分析
2.1 甲醇浸提液对白菜籽发芽的影响
2.1.1 种皮、种实甲醇浸提液对白菜籽发芽率的影响 不同处理的种皮、种实甲醇浸提液对白菜籽发芽率的影响结果如图1所示。不同处理种实的甲醇浸提液处理的白菜籽发芽率分别较对照降低了2.81%,20.83%和29.16%;不同处理种皮浸提液处理的白菜籽发芽率分别较对照降低了1.35%,6.98%和11.15%。总的来说,随着浸提液质量浓度的增加,所处理的白菜籽发芽率呈逐渐降低的趋势,高质量浓度甲醇浸提液对白菜籽的抑制作用显著。
2.1.2 种皮、种实甲醇浸提液对白菜苗苗高生长的影响 不同处理的种皮、种实甲醇浸提液对白菜苗的苗高生长量的影响结果见图2。与对照相比,种实各处理浸提液对白菜苗高生长分别降低了18.0%,40.0%和58.0%;而不同处理种皮浸提液处理的白菜苗高却比对照分别增加7.4%,7.4%和1.6%。以上结果表明:胚乳浸提液处理的白菜籽苗高随着浸提液质量浓度的增加,苗高呈现出逐渐降低的趋势,说明胚乳浸提液对白菜苗高有抑制作用;而种皮浸提液对苗高生长的抑制作用不显著。
2.1.3 种皮、种实甲醇浸提液对白菜苗的根长生长量的影响 不同处理的种皮、种实甲醇浸提液对白菜苗的根长生长量的影响结果见图3。与对照相比,种实各处理浸提液处理的白菜根长分别降低了76.4%,88.76%和 91.6%;而不同处理种皮浸提液处理的白菜根长在 0.03 g·mL-1和 0.06 g·mL-1时比对照增加了34.79%和18.53%,质量浓度为0.10 g·mL-1时,根长比对照降低48.77%。以上结果表明:种实浸提液处理的白菜苗根长随着浸提液质量浓度的增加,根长呈现出急剧降低的趋势,说明种实浸提液对白菜苗根长有很强的抑制作用;而低质量浓度的种皮浸提液处理的白菜苗根长较对照有一定的促进作用(其原因有待进一步研究),随着质量浓度的增大,逐渐转为抑制作用。

图1 甲醇浸提液对白菜籽发芽率的影响Figure 1 Effect of methanol extracted from different seed parts on germination rate of Brassica campestris

图2 甲醇浸提液对白菜苗高的影响Figure 2 Effect of methanol extracted from different seed parts on the height of Brassica campestris
综上所述,紫梗籽银桂种子的种皮和种实的甲醇浸提液中含有某些抑制物质,这些物质对白菜籽的发芽率、苗高和根长生长量均有一定的抑制作用,并随着浸提液质量浓度的提高,抑制作用逐渐加强。相比而言,种实的甲醇浸提液对白菜籽发芽率的抑制作用比种皮浸提液的抑制作用更大。
2.2 不同质量浓度赤霉素处理的紫梗籽银桂种子发芽率的变化
未经低温层积处理的紫梗籽银桂种子置床30 d仍未见种子萌发,后将部分种子的种皮和胚乳剥除成离体胚,再置床58 d,仅见胚体子叶部分转为绿色,而胚轴仍为白色;而未剥除的种子仍无萌发迹象。这充分证明前面的研究结果:紫梗籽银桂种子中含有发芽抑制物质,种子存在生理休眠现象。
经不同质量浓度赤霉素处理并经低温层积的紫梗籽银桂种子发芽率见图4。从4图中可以看出,不同处理的紫梗籽银桂种子在层积60 d后开始发芽,且以 1 000 mg·L-1赤霉素处理的发芽率最高,随着层积时间的延续,各处理发芽率逐渐增加,至层积105 d时发芽率达最高,其中G10在75 d时发芽率为71.11%,93 d时为67.78%,105 d时为72.37%,基本上保持稳定。G5和G15发芽率的提高滞后于G10,105 d时分别为63.82%和70.32%。而未经赤霉素处理的种子在层积105 d后其发芽率仅为26.67%。方差分析结果表明:不同质量浓度赤霉素、不同层积时间以及处理 ×时间的种子的发芽率差异均达到极显著水平。由此可见,用赤霉素处理种子后再进行低温层积可显著解除休眠,其中以 1 000 mg·L-1为最佳,只要75 d左右就可打破桂花种子的休眠。

图3 甲醇浸提液对白菜根长生长的影响Figure 3 Effect of methanol extracted from different seed parts on the root height of Brassica campestris
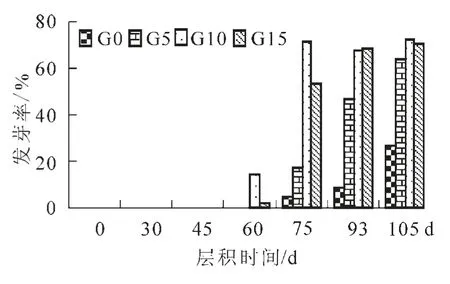
图4 紫梗籽银桂种子层积过程中发芽率的变化Figure 4 Changes on germination percentage of‘Zigeng Ziyin’seeds during stratification
3 结论与讨论
对紫梗籽银桂种子的种皮、种实的甲醇浸提液进行生物测定。结果表明:种皮、种实的甲醇浸提液对白菜籽的发芽率、白菜幼苗高生长和根生长量均有一定程度的抑制。这种抑制作用随着质量浓度的增加而增大,并且胚乳浸提液的抑制作用比种皮的大。在实验中还观察到白菜苗根系出现膨大、扭曲、向上生长等畸形现象。由此可见,紫梗籽银桂种子中存在一些抑制白菜籽萌发和影响生长的物质。相同的实验在南方红豆杉 Taxus chinensis var.mairei[5],青钱柳 Cyclocarya paliurus[6],洋白蜡Fraxinus pennsylvanica[7]等树种上均已得到证实。但紫梗籽银桂种子中究竟含有什么物质,多大质量浓度才会对种子休眠起作用仍需进一步研究。
种子休眠一般分为外源休眠、内源休眠和综合休眠。种子物理透性常引起外源休眠,休眠程度与种皮的不透性或硬实程度有关。如红松Pinus koraiensis种子外种皮坚硬,阻碍了种胚与外界的气体交换,进而引起种子休眠[8]。内源休眠有的因胚发育不完全所致,有的因种子萌发的生理抑制造成。如导致银杏Ginkgo biloba[9]种子深休眠的主要原因是胚发育不完全;玫瑰Rosa rugosa种子种皮中存在的抑制物脱落酸是引起其休眠的主要原因。南京椴Tilia mqueliana[10],对节白蜡Fraxinus hupensis[11]种子的果皮、种皮及胚乳中都存在导致休眠的抑制物质。综合休眠则是内外源因素互相联系共同控制着种子的休眠,从而使种子休眠问题的研究复杂化[12]。秤锤树 Sinojackia xylocarpa[13]和青钱柳[14]种子的休眠,除了种皮机械束缚和透气性差的原因外,种胚还需要一段低温的生理后熟过程,属于综合休眠类型。本研究中,紫梗籽银桂种子的种皮虽然较为坚硬,但种皮较薄,种子萌发时种胚可以顺利突破种皮,表明种皮不会对种胚的萌发造成机械性阻碍;对种子吸水速率的观察也表明紫梗籽银桂种皮具有良好的透性[15]。紫梗籽银桂种子的甲醇浸提液对白菜籽的发芽率、白菜苗高和根长等均有一定的抑制作用,说明种子中含有抑制物质可能是导致桂花种子休眠的主要原因,也即桂花种子的休眠属生理(内源)休眠。
种子休眠的原因各不相同,因而解除休眠的方法也各不相同。层积处理是一种在生产上应用很广泛的方法,尤其是对解除由于生理后熟引起的种子休眠很有效。张义等[3]研究发现,紫梗籽银桂种子胚具有生理后熟现象,低温层积能够增进种子萌发,在采收后一般需经沙藏后播种。本研究用赤霉素处理浸种后低温层积,60 d时就有部分种子萌发,并且75 d后发芽率基本达到最大值,之后种子的发芽率保持稳定,表明层积处理对解除桂花种子休眠很有效,层积75 d左右即可解除休眠。
[1]LIU Zhen.Studies on the dormancy in Idesia polycarpa distributing in the subtropical zone[J].Bull Mie Univ For,2000,24:107-161.
[2]NAGATA H,NAKASHIMA A,YURUGI Y.Bud dormancy in woody plants[J].Bull Mie Univ For,1994,18:17-42.
[3]张义,宋春燕.赤霉素浸种与低温层积对桂花种子发芽的影响[J].中国林副特产,2005(6):9-10.ZHANG Yi,SONG Chunyan.Impact of Osmanthus fragrans seed gibberellin processing and low temperature stratification on its seed germination performan[J].Q For By-prod Spec China,2005(6):9-10.
[4]袁王俊,董美芳,尚富德.利用离体培养技术打破桂花种子休眠的试验[J].南京林业大学学报:自然科学版,2004,28(增刊):91-93.YUAN Wangjun,DONG Meifang,SHANG Fude.Preliminary study on breaking the seed dormancy of Osmanthus fragrans by in vitro culture[J].J Nanjing For Univ Nat Sci Ed,2004,28(supp):91-93.
[5]张艳杰,高捍东,鲁顺保.南方红豆杉种子中发芽抑制物的研究[J].南京林业大学学报:自然科学版,2007,31(4):51-56.ZHANG Yanjie,GAO Handong,LU Shunbao.Germination inhibitors in methanol extract from Taxus chinensis var.mairei seed[J].J Nanjing For Univ Nat Sci Ed,2007,31(4):51-56.
[6]杨万霞,方升佐.青钱柳种皮甲醇浸提液的生物测定[J].植物资源与环境学报,2005,14(4):11-14.YANG Wanxia,FANG Shengzuo.The bioassay of the methanol extract from Cyclocarya paliurus seed coat[J].J Plant Resour Environ,2005,14(4):11-14.
[7]郑彩霞,高荣孚.脱落酸和内源抑制物对洋白蜡种子休眠的影响[J].北京林业大学学报,1991,13(4):19-22.ZHENG Caixia,GAO Rongfu.Effect of ABA and other endogenous inhibitors on Fraxinus pennsylvanica Marsh.seeds dormancy[J].J Beijing For Univ,1991,13(4):19-22.
[8]郑彩霞,王九龄,智信.国内外红松种子休眠及催芽问题研究动态[J].世界林业研究,1997,10(5):3-9.CHEN Caixia,WANG Jiuling,ZHI Xin.Research state of the dormancy and pregermination of Pinus koraiensis seed in China and aboard[J].World For Res,1997,10(5):3-9.
[9]蔡春菊,曹帮华,许景伟,等.银杏种子后熟生理的研究[J].山东林业科技,2002(6):7-9.CAI Chunju,CAO Banghua,XU jinghua,et al.Studies on delayed-ripening physiology of Ginkgo seeds[J].J Shandong For Sci Technol,2002(6):7-9.
[10]史锋厚,沈永宝,施季森.南京椴种子发芽抑制物研究[J].福建林学院学报,2007,27(3):222-225.SHI Fenghou,SHEN Yongbao,SHI Jisen.Study on germination inhibitor of Nanjing linden seeds[J].J Fujian Coll For,2007,27(3):222-225.
[11]叶要妹,王彩云,史银莲.对节白蜡种子休眠原因的探讨[J].湖北农业科学,1999,78(4):45-47.YE Yaomei,WANG Caiyun,SHI Yinlian.Preliminary study on cause of seed dormancy of Fraxinus hupensis[J].Hubei Agric Sci,1999,78(4):45-47.
[12]KHAN A A.Quantification of plant dormancy:introduction to the workshop[J].HortScience,1997,32(4):608-609.
[13]史晓华,黎念林,金玲,等.秤锤树种子休眠与萌发的初步研究[J].浙江林学院学报,1999,16(3):228-233.SHI Xiaohua,LI Nianlin,JIN Ling,et al.Seed dormancy and germination of Sinojackia xylocarpa[J].J Zhejiang For Coll,1999,16(3):228-233.
[14]史晓华,徐本美,黎念林,等.青钱柳种子休眠与萌发的研究[J].种子,2002(5):5-8.SHI Xiaohua,XU Benmei,LI Nianlin,et al.Study on dormancy and germination of Cyclocarya paliurus(Batal.)Iljinskaja[J].Seed,2002(5):5-8.
[15]杨秀莲,丁彦芬,甘习华.‘紫梗籽银桂’种子休眠原因的初步探讨[J].江苏林业科技,2007,34(6):18-19,45.YANG Xiulian,DING Yanfen,GAN Xihua.Primary research on dormancy mechanism of seeds of Zigengziyin osmanthus[J].J Jiangsu For Sci Technol,2007,34(6):18-19,45.
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24
安徽农业科学(2022年18期)2022-10-13
中国油料作物学报(2022年2期)2022-05-13
黑龙江大学工程学报(2021年2期)2021-06-22
花卉(2020年24期)2021-01-04
防护林科技(2020年11期)2020-12-30
东北农业大学学报(2020年11期)2020-12-17
潍坊学院学报(2020年6期)2020-11-22
中国食品学报(2019年3期)2019-01-13
天津农业科学(2018年3期)2018-03-24
