
抗菌肽Lactoferricin B的原核表达及其纯化
2010-08-08 06:37:08赵晓宇冯兴军孟利强张淑梅王玉霞张先成
东北农业大学学报 2010年9期
赵晓宇,冯兴军,孟利强,张淑梅,王玉霞,张先成
(1.黑龙江省科学院微生物研究所,哈尔滨 150010;2.东北农业大学动物科学技术学院,哈尔滨 150030)
传统抗生素的应用所具有的负面效应日益严重。抗生素的滥用所产生的诸如对人类的危害、细菌耐药性的增强和耐药菌株的增加、对环境的潜在危害等问题已受到全世界的关注。抗菌肽是机体在受到病原体感染时产生的一类小分子多肽。它作为一种先天性免疫因子,在保护机体免受病原体侵害方面具有重要作用,且不易产生菌株耐药性[1]。牛乳铁蛋白活性多肽(LfcinB)最先是由Bellamy等于1992年在牛乳铁蛋白的酶解产物中分离得到的一段N端多肽,其抗菌活性比牛乳铁蛋白高400多倍[1-2]。LfcinB位于牛乳铁蛋白的17~41位氨基酸残基之间,由25个氨基酸残基组成,分子结构以两亲性的β-折叠为主。LfcinB具有广谱抗菌活性[3]、强大的抑菌杀菌作用[4-5]、、抗病毒[6]、抑制肿瘤生长[7-8]、调节免疫及消炎[9]、抗寄生虫[10]等多种生物学功能。LfcinB在医药、保健、食品保鲜等领域应用前景广泛。然而,来源上的限制阻碍了LfcinB的应用发展。基因工程技术为解决LfcinB来源问题提供了有效手段。本研究利用大肠杆菌表达系统高效表达了LfcinB,为重组表达LfcinB提供技术路线和理论依据。
1 材料与方法
1.1 材料
1.1.1 质粒和菌株
质粒pGEX-4T-2(购于Amersham公司);E.coli DH5α 感受态细胞和 E.coli Rosetta(DE3)(购于北京全式金公司);金黄色葡萄球菌(Staphylococcus aureus ATCC25923)由本实验室保存。
1.1.2 试剂和仪器
限制性内切酶Bam HⅠ和SalⅠ(购于NEB公司),T4DNA连接酶(购于宝生物公司);DNA分子质量标准、蛋白分子质量标准(购于北京全式金公司)。DNA酶(购于Sigma公司);异丙基-B-D-硫代半乳糖苷(IPTG)、氨苄青霉素(Amp)、还原型谷胱甘肽(L-glutathion reduced)(购于Sigma公司);质粒提取试剂盒(购于北京全式金公司);琼脂糖凝胶回收试剂盒(购于Qiagen公司);其他试剂均为国产分析纯。
仪器设备:UVP Auto Chemi System凝胶成像系统(美国),分析软件LabWorks 4.6,BIORADMINI电泳仪,魧KTApurifier蛋白纯化系统(美国),GSTrap FF 1 mL亲和层析柱(美国),McFarland Standard比浊管(法国)。
1.2 方法
1.2.1 基因的设计与合成
LfcinB的氨基酸序列为Phe-Lys-Cys-Arg-Arg-Trp-Gln-Trp-Arg-Met-Lys-Lys-Leu-Gly-Ala-Pro-Ser-Ile-Thr-Cys-Val-Arg-Arg-Ala-Phe,根据E.coli密码子偏爱性设计合成编码LfcinB 25个氨基酸的基因片段[11],同时在基因3'端加上终止密码子TAA,分别在两端加上Bam H I与Sal I的粘性末端。整个基因的正链为5'GATCCTTCAAATGCCGC CGTTGCAGTGCGTATGAAAAAACTGGGTCGCCGTC TATTACCTGCGTGCGTCGCGCGTTCTAAG 3',反链为5'TCGACTTAGAACGCGCGACGCACGCAGGTAA TAGACGGCGCACCCAGTTTTTTCATACGCCACTGC CAACGGCGGCATTTGAAG 3'。化学合成正反链,退火,连接到pGH的Bam H I与Sal I位点之间,构建克隆载体pGH-X-LCB[12]。
1.2.2 原核重组表达载体的构建
用Bam HⅠ和SalⅠ限制性内切酶切割质粒pGH-X-LCB,回收基因片段连接到原核表达载体pGEX-4T-2的Bam HⅠ和SalⅠ位点之间,构建重组表达质粒pGEX-4T-2-LCB,转化DH5α,经双酶切鉴定筛选得到阳性克隆,并将阳性克隆送至上海生工生物工程公司测序验证。
1.2.3 原核表达
重组表达载体pGEX-4T-2-LCB转化E.coli Rosetta(DE3),挑取阳性菌落转接于5 mL含50 μg·mL-1氨苄青霉素的LB培养基中,37℃摇床震荡培养过夜。同时接种含母本pGEX-4T-2载体的转化菌作对照。吸取500 μL菌液转接入50 mL含100 μg·mL-1氨苄青霉素的LB培养基中,37℃摇床培养至OD600=0.6左右,加IPTG至终浓度为0.3 mmol·L-1,在33℃诱导4 h。取1 mL菌液,加入等体积的上样缓冲液100℃煮沸5 min,12 000 r·min-1离心5 min,取上清,15%SDS-PAGE分析。考马斯亮蓝染色、脱色后,用UVP AutoChemi System凝胶成像系统拍照,LabWorks4.6软件测定表达蛋白的相对含量。
1.2.4 融合蛋白的纯化
1.2.4.1 样品制备
将含有阳性重组子pGEX-4T-2-LCB E.coli Rosetta(DE3)按照上述条件诱导后,5 000 r·min-1离心10 min收集菌体;按100 mL培养物的菌体沉淀加入4 mL PBS重悬菌体,-20℃冷冻后,冰浴融解;加入溶菌酶至终浓度1 mg·mL-1,冰上放置30 min;加入Triton X-100至0.1%,震荡混匀;加入DNA酶和RNA酶至终浓度5 μg·mL-1,4℃温浴10 min;加入DTT至终浓度1 mmol·L-1,4 ℃12 000 r·min-1离心30 min,收集上清备用。
1.2.4.2 GST-LfcinB融合蛋白纯化
上清液中的GST-LfcinB融合蛋白通过GST标签亲和层析进行分离纯化。上样体积:100 μL。纯化条件:流速 0.5 mL·min-1,压力 0.25 MPa,Binding Buffer(1× PBS :140 mmol·L-1NaCl,2.7 mmol·L-1KCl,10 mmol·L-1Na2HPO4,1.8 mmol·L-1KH2PO4,pH 7.3)。处理预装柱,UV280调零,基线稳定后上载样品,待流出峰经过,基线完全回归零点后更换 Elution Buffer(50 mmol·L-1Tris-HCl,10 mmol·L-1还原性谷胱苷肽,pH 8.0)进行洗脱。回收洗脱峰样品经SDS-PAGE电泳分析,具体方法参照文献[12]。样品制备参照文献[13]。
1.2.5 抗菌肽的活性测定
将金黄色葡萄球菌于MH液体培养基活化培养,比浊测定菌数固定在6×108~9×108之间,涂布在MHB固体培养基表面,待菌液吸干后取灭菌纸片放置于培养基上,每个试验纸片滴加10 μL纯化后的LfcinB重组蛋白,37℃过夜培养,观察是否有抑菌圈。对照为灭菌水。
2 结果与分析
2.1 质粒的提取及酶切
质粒pGH-X-LCB以及酶切后的产物经2%琼脂糖凝胶电泳后,得到清晰的DNA条带,大小与预计结果相同,凝胶回收试剂盒回收目的片段,与表达载体pGEX-4T-2连接后转化DH5α。
2.2 重组表达载体的构建
牙签挑取阳性克隆,在LB液体培养后,提取质粒,用Bam HⅠ和SalⅠ限制性内切酶双酶切,结果见图2。在经过双酶切后样品2中在84 bp处存在一清晰条带。经过测序验证,重组质粒中含有LfcinB基因片段,与设计的基因片段完全吻合。
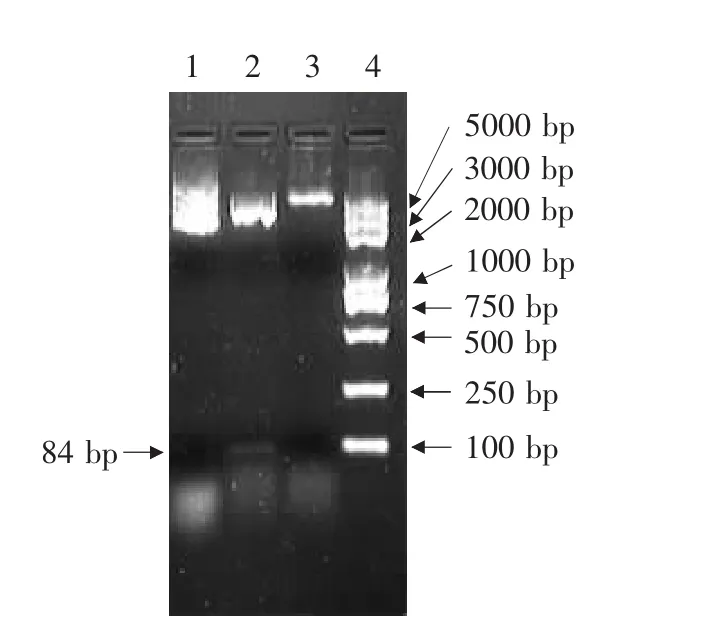
图1 质粒pGH-X-LCB、pGEX-4T-2质粒及其酶切电泳Fig.1 Electrophoresis analysis of plasmid pGH-X-LCB,pGEX-4T-2 and digested products
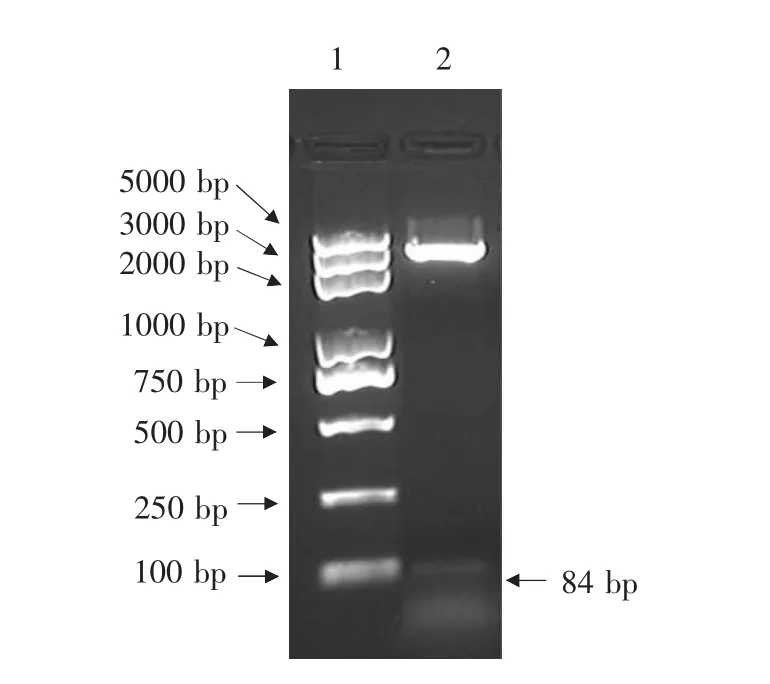
图2 质粒pGEX-4T-2-LCB的酶切验证Fig.2 Electrophoresis analysis of plasmid pGEX-4T-2-LCB digested by Bam H I and Sal I
2.3 原核表达
诱导表达后的样品经SDS-PAGE分析,如图3所示。样品1在29 ku处存在一条蛋白条带,为GST-LfcinB融合蛋白。通过凝胶分析软件Lab-Works 4.6对工程菌株裂解上清液中蛋白含量进行测定,结果表明所表达的融合蛋白占总蛋白含量的20%以上,并且所表达的蛋白多为可溶性形式。
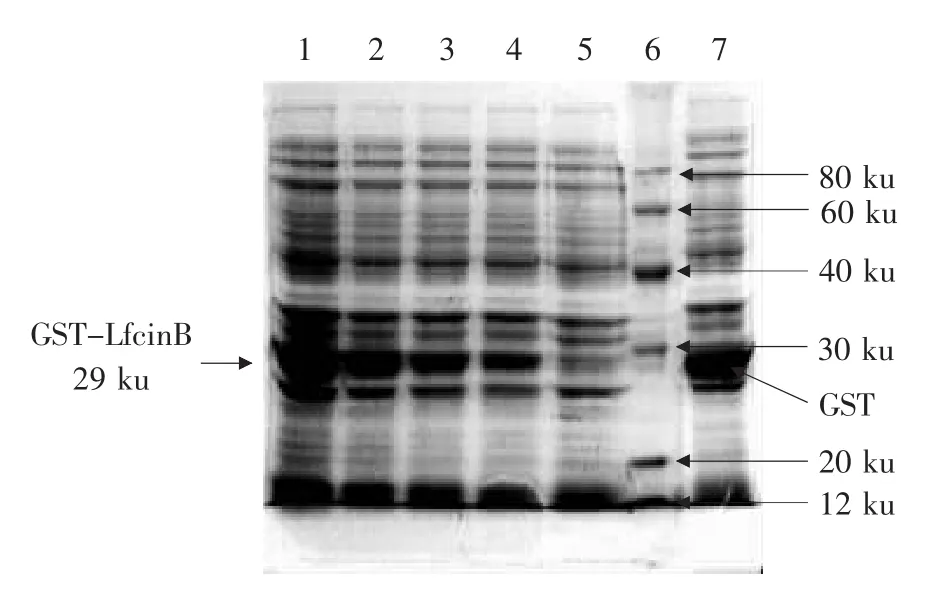
图3 LfcinB融合蛋白在E.coli Rosetta(DE3)中的诱导表达Fig.3 Induced expression of LfcinB fusion protein in E.coli Rosetta(DE3)
2.4 融合蛋白的纯化
利用GE魧KTA蛋白分离纯化系统对表达的融合蛋白GST-LfcinB进行亲和层析纯化,洗脱曲线如图4所示。共获得P1、P2和P33个洗脱峰,其中P1和P2为流出峰,P3则为洗脱峰。将洗脱峰进行SDS-PAGE分析,结果如图5所示。收集获得的样品在29 ku位置有一清晰条带,为纯化后的GST-LfcinB融合蛋白,其纯度高于95%。
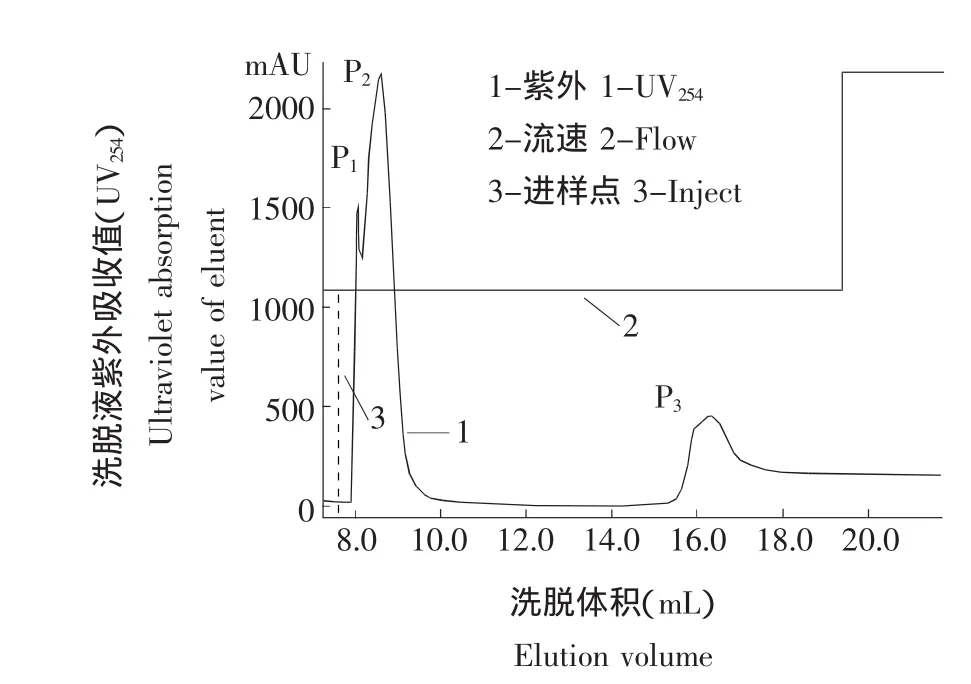
图4 融合蛋白GST-LfcinB的洗脱曲线Fig.4 Eluting curve of fusion protein GST-LfcinB
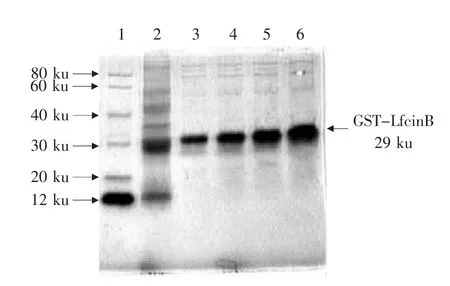
图5 纯化蛋白电泳Fig.5 Electrophoresis analysis of purified fusion protein GST-LfcinB
2.5 抗菌肽的活性测定
将经过GE魧KTA蛋白分离纯化系统纯化获得的融合蛋白GST-LfcinB,进行体外活性测定,结果如图6所示。利用纸片薄层琼脂糖平皿扩散法测定重组LfcinB抗菌活性。在金黄色葡萄球菌生长的平板上,试验孔明显出现抑菌圈,而对照孔菌株生长正常,表明重组的LfcinB具有抗菌活性。
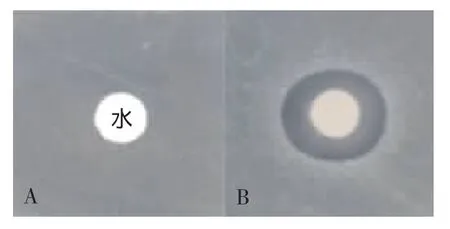
图6 重组LfcinB对金黄色葡萄球菌(S.aureas ATCC25923)的抑菌活性Fig.6 Antimicrobial activity of recombinant LfcinB to Staphylococcus aureas ATCC25923
3 讨论与结论
获得目的基因有多种方法,但多数费时耗力。目前DNA合成仪能够精确合成人们需要的目的核苷酸片段,为获得目的基因提供了一条新途径。因此,人们可以根据研究要求设计、合成具有更加实际应用价值与研究价值的编码基因。合成基因片段的长度从FcyRIIb1基因的几百碱基到IL-2基因的上千碱基不等[14-15],并且已经试验证明合成的基因能够成功表达。因此,人工合成基因的方法避免了以往传统基因克隆方法获得的目的基因片段容易产生的碱基错配以及获得目的基因费时、费力的弊端,可以快速获得任意核苷酸序列的目的基因片段。本研究获得的LfcinB编码基因长度仅为75 bp,使用人工合成的方法不仅能够快速获得该基因片段、节约试验耗材,而且通过该方法成功地优化了原有的密码子,使得该基因能够更好的表达,并且在大肠杆菌中不形成包涵体,主要以可溶性蛋白为主,为蛋白后期分离纯化打下了良好的基础。
诱导条件对重组蛋白表达有显著影响,诱导温度、IPTG浓度以及诱导表达时间是主要的影响条件,本研究针对三种影响因素对诱导条件进行了优化,获得了合适的诱导表达条件。在优化的诱导条件下,IPTG浓度0.3 mmol·L-1,表达温度33℃,诱导时间4 h,GST基因融合表达系统成功表达了LfcinB,获得了理想的表达效果,诱导蛋白表达量占细胞总蛋白的20%以上,并且所表达的融合蛋白大部分以可溶性形式存在,避免了繁琐复杂的包涵体处理操作过程,可溶性表达方式存在的蛋白可以直接利用亲和层析进行纯化获得融合蛋白,方便了进一步的分离纯化过程。
本研究利用GE魧KTApurifier蛋白纯化系统结合GST亲和层析预装柱成功从裂解液上清中纯化出LfcinB重组融合蛋白,分离纯化条件稳定,纯化效率高,回收率大,对目标蛋白无影响。纯化目标蛋白纯度在95%以上,达到了极好的分离效果。
*冯兴军为本文的同等贡献者。
[1] Bellamy W,Takase M,Wakabayashi H,et al.Antibacterial spectrum of lactoferricin B,a potent bactericidal peptide derived from the N-terminal region of bovine lactoferrin[J].J Appl Bacteriol,1992,73(6):472-479.
[2] Bellamy W,Takase M,Yamaudhim K,et al.Identification of the bactericidal domain of lactoferrin[J].Biophys Acta,1992,1121:130-136.
[3] 唐传核,曹劲松,彭志英.乳铁蛋白最新研究进展-活性多肽以及生理功能(Ⅱ)[J].中国乳品工业,2000,28(5):44-47.
[4] Wakabayashi H,Matsumoto H,Hashimoto K,et al.N-Acylated and D enantiomer derivatives of a nonamer core peptide of lactoferricin B showing improved antimicrobial activity[J].Antimicrob Agents Chemother,1999b,43(5):1267-1269.
[5] Wakabayashi H,Uchida K,Yamauchi K,et al.Lactoferrin given in food facilitates dermatophytosis cure inguinea pig models[J].J Antimicro Chemothera,2000,46:595-601.
[6] Anderson J H,Jenssen H,Gutteberg T J.Lactoferrin and lactoferricin inhibit Herpes simplexs 1 and 2 infection and exhibit synergy when combined with acyclovir[J].Antiviral Res,2003,58(3):209-215.
[7] Yoo Y C,Watanabe R,Koike Y,et al.Apoptosis in human leukemic cells induced by lactoferricin,a bovine milk protein-derived peptide:involvement of reactive oxygen species[J].Biochem Biophys Res Commun,1997,237(3):624-628.
[8] Yoo Y C,Watanabe S,Watanabe R,et al.Bovine lactoferrin and lactoferricin inhibit tumor metastasis in mice[J].Adv Exp Med Biol,1998,443:285-291.
[9] Krieg A M,Yi A K,Matson S,et al.CpG motifs in bacterial DNA trigger direct B cell action[J].Nature,1995,374(6522):546-549.
[10] Tanaka T,Omata Y,Saito A,et al.Toxoplasma gondii:parasiticidal effects of bovine lactoferricin against parasites[J].Exp Parasitol,1995,81(4):614-617.
[11] Hale R S,Thompson G.Codon optimization of the gene encoding a domain from human type 1 neurofibrot in expression min protein results in a threefold improvement level in Escherichia coli[J].Protein Expr Purif,1998,12:185-188.
[12] 萨姆布鲁克J,拉塞尔D W.分子克隆实验指南[M].黄培堂等译.3版.北京:科学出版社,2002:1228-1232,1713-1720.
[13] 马歇克D R,门永J T,布格斯R R,等.蛋白质纯化与鉴定实验指南[M].朱厚础等译.北京:科学出版社,1999:259-261.
[14] Chen H Y,Bierknes R,Kumar R,et al.Determination of the optimal aligned spacing between the SD sequencesand the translation initiation codon of E.coli mRNAs[J].J Nucleic Acids Res,1994,22:4953-4957.
[15] Williams D P,Regier D,Akiyoshi D,et al.Design,synthesis and expression of a human interleukin-2 gene incorporating the codon usage bias found in highly expressed Escherichia coli genes[J].Nucleic Acids Res,1988,17(24):10453-10467.
猜你喜欢
科学(2022年4期)2022-10-25 02:43:32
数学物理学报(2021年4期)2021-08-30 08:27:48
新世纪智能(数学备考)(2020年10期)2021-01-04 00:37:50
食品科学(2018年10期)2018-05-23 01:27:28
中成药(2017年12期)2018-01-19 02:06:52
中国交通信息化(2017年8期)2017-06-06 07:16:47
西南医科大学学报(2015年1期)2015-08-22 13:01:46
天津科技大学学报(2015年3期)2015-04-16 04:54:59
中国当代医药(2015年9期)2015-03-01 02:01:59
西南军医(2015年6期)2015-01-23 01:25:50
