
皮肤创伤愈合过程中基质细胞衍生因子-1表达及其影响因素
2010-08-06 11:42黄宏郭敏徐祥杨策孙宏振戴卉冯帅南王莎丽张波
成都医学院学报 2010年3期
黄宏,郭敏,徐祥,杨策,孙宏振,戴卉,冯帅南,王莎丽,张波
(1.第三军医大学第三附属医院野战外科研究所,创伤、烧伤与复合伤国家重点实验室,重庆 400042;2.重庆医科大学第二附属医院创伤烧伤科,重庆 400010;3.重庆医科大学生理教研室,重庆 400016)
皮肤创伤修复/愈合是一个复杂而有序的过程。创伤部位释放的多种细胞因子和生长因子严格调控创伤愈合过程,这些细胞因子在局部表达水平、表达类型以及表达规律决定了参与修复细胞的活性及功能状态,从而对组织修复速率、纤维化程度以及修复结局产生极其重要的影响[1]。基质细胞衍生因子-1α(stromal cell-derived factor-1,SDF-1)是一种由基质细胞合成分泌、缺血缺氧诱导表达、对人类CD34+祖细胞等有显著趋化作用的CXC类强效趋化因子[2,3],能与CXCR4特异性结合构成SDF-1/CXCR4生物轴,参与并调节器官发生、干细胞归巢、癌症转移和重要脏器损伤修复等病理生理过程[4-8]。皮肤损伤必然发生缺血缺氧,存在明显诱导创面SDF-1表达的诱因,而大量文献证实SDF-1是参与调节组织/器官损伤修复的关键细胞因子[9-11],由此我们推测,SDF-1可能参与皮肤创伤愈合过程,但是对其在皮肤创伤修复过程中的表达及作用了解甚少。为此,本实验旨在探讨SDF-1在皮肤创伤修复过程中的表达规律及其可能作用,为进一步丰富创面愈合的分子机制提供新的实验依据。
1 材料与方法
1.1 主要试剂
一抗:SDF-1α抗体(购于美国Santa Cruz Biotechnology公司);RT-PCR试剂盒(购于 TaKaRa宝生物公司);二氨基联苯胺(……DAB)、SP-9000通用免疫组化试剂盒(购于北京中衫金桥公司);SDF-1α及GAPDH 引物(由上海生工合成)。
1.2 实验动物模型的建立
清洁级雄性昆明小鼠32只(第三军医大学实验动物教研室提供),体重25-30 g.小鼠背部剃毛后,应用速眠新Ⅱ(1 ml·kg-1)肌肉麻醉,常规消毒,于背部近颈侧以脊柱为中线切除约1.5 cm ×1.5cm的正方形全层皮肤,创面以消毒纱布止血后暴露,不用任何药物,常规饲养,观察伤口的自然愈合过程。分别于伤后第 1、2、3、4、5、7、10 和 14 天随机取 4 只小鼠,麻醉后取材,切取创面组织。部分组织4%中性多聚甲醛缓冲固定液固定标本,常规脱水,包埋,切成5 μ m切片,用于组化染色。
同窝新生1 d和2周龄的昆明种小鼠各12只,清洁级,雌雄不拘。新生组小鼠于背部切除0.5 cm×0.5 cm的全层皮肤,创面以无菌纱布止血后暴露,母乳喂养;2周龄小鼠用适量戊巴比妥钠(30 mg/kg)腹腔注射麻醉后,背部剃毛,常规消毒,于背部近颈部以脊柱为中线切除约1.0 cm×1.0 cm的全层皮肤,创面以消毒纱布止血后暴露,不用任何药物,常规饲养,于伤后1、2和3 d均随机处死4只小鼠,切取创面及创缘正常侧0.3 cm组织,切取正常小鼠皮肤组织作为对照组,所有标本液氮冻存,用RT-PCR法检测SDF-1α基因表达。
1.3 小鼠皮肤成纤维细胞的分离培养
清洁级6-8周龄雄性昆明种小鼠,体重(28±2)g.3%戊巴比妥钠30 mg/kg腹腔注射麻醉,背部剃毛后常规消毒,取背侧皮下组织。采用组织块法分离培养小鼠皮肤成纤维细胞,分别用 0.1、1、10、100 ng/ml浓度的肿瘤坏死因子α(tumor necrosis factor α,TNF-α)和白细胞介素1(interleukin 1,IL-1)作用于成纤维细胞,48 h后收获细胞,对照组仅加培养基,不加任何刺激物。应用RT-PCR法检测细胞SDF-1α的基因表达。
1.4 免疫组化染色
取创面以及周围0.5 cm宽的皮肤组织,室温下4%中性多聚甲醛缓冲固定液固定标本48 h,逐级脱水,石蜡包埋,切成5 μ m 厚度切片。切片脱蜡至水化;经0.3%过氧化氢-甲醇去除内源性过氧化物酶,0.1%Tritox-100 37℃孵育 8 min;以 10%正常山羊血清封闭,滴加一抗SDF-1α(1∶300)4℃过夜;次日,PBS洗涤后,加入生物素化二抗和辣根过氧化物酶标记链酶卵白素分别在37℃孵育30 min,最后DAB显色,苏木素复染后,脱水、透明、中性树胶封片。显微镜下观察,胞浆内有棕黄色颗粒的为阳性细胞。免疫组化阴性对照以PBS代替一抗。
1.5 RT-PCR检测SDF-1α mRNA 表达
采用异硫氢酸胍一步法提取皮肤创面组织中的总RNA,紫外分光光度计测定RNA的浓度,凝胶电泳分析RNA结构。采用半定量逆转录聚合酶链反应(RT-PCR)技术对转录产物进行扩增,以三磷酸甘油醛脱氢酶(……,GAPDH)作为内参对照,所用引物为:SDF-1α,上游引物:5'-AAA CTG TGC CCT TCA GAT TGT T-3',下游引物:5'-CGG GGA ACT AGT T TT TCC T TT T-3',产物大小为209碱基对(bp);GAPDH,上游引物:5'-TGA CAT CAA GAA GGT GGT GAA G-3',下游引物:5'-ATC CTG T TG CTG TAG CCG TAT T-3',产物大小470 bp.取 PCR反应产物 10 μ l在1×TAE制备2.0%琼脂糖凝胶中电泳,加入浓度为0.5 μ g/ml的溴化乙啶(……,EB)染色,用DL2000为标准来判断PCR产物片段大小,恒压5 V/cm电泳,然后用凝胶扫描仪观察并扫描。实验结果以检测条带与GAPDH条带的总灰度值的比值作为检测目的片段的相对表达水平。
1.6 统计学处理
2 结果
2.1 免疫组织化学染色
正常皮肤组织表达力量SDF-1,主要表达在皮肤表皮基底层和真皮层成纤维细胞(图1),正常皮肤表皮基底层细胞呈SDF-1阳性表达,而真皮间质也可见少量阳性表达的细胞,主要是梭形的成纤维细胞。而创伤后第1天真皮组织内SDF-1阳性细胞显著增加,主要位于创缘表皮基底层、毛囊外周、血管上皮细胞以及间质细胞胞浆内;伤后第3天,阳性表达细胞显著减少,而血管周围仍有一些SDF-1阳性细胞;伤后第5、7天,SDF-1阳性细胞又显著增加,主要是位于真皮的基质细胞、创沿表皮细胞、血管周围和皮脂腺的一些细胞;伤后14天在真皮层成纤维细胞表达SDF-1,基底层未见有表达。
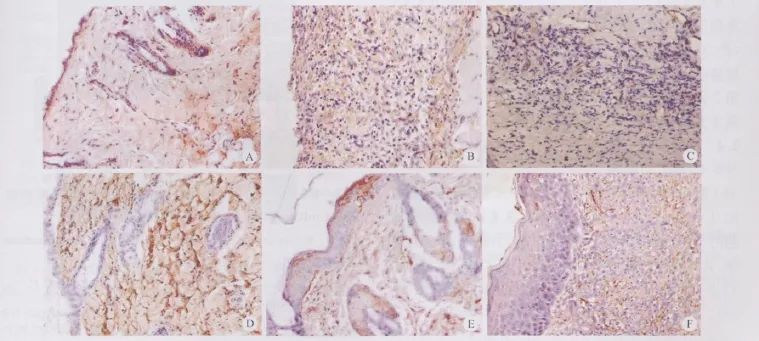
图1 损伤前后SDF-1α在皮肤创面的表达(S-P,×400)Fig.1 Expression of SDF-1αat normal skin and wound skin after injury(S-P,×400)

表1 各组小鼠损伤后不同时间的SDF-1α的半定量结果(%,n=4)Tab.1 Semiquantitative analyses of SDF-1αgene amplification expressed by each group during initial stage of wound healing(%,n=4)
2.2 SDF-1 mRNA表达变化
正常皮肤组织存在SDF-1 mRNA表达,当皮肤组织发生缺损伤后1 d,SDF-1 mRNA表达较正常组织明显增加(P<0.01)。随后,伤后2、3 d,SDF-1 mRNA表达减少,2 d时基本恢复到正常组织水平,3 d时表达量最低,低于正常组织水平(P<0.05)。伤后4、5 d SDF-1 mRNA表达又开始回升(P<0.01),5 d时达峰值(P<0.01),以后又逐渐下降,14 d时基本恢复至正常水平。小鼠全层皮肤缺损伤创伤愈合过程中创面SDF-1 mRNA表达呈双峰(图 2)。
2.3 新生组和2周龄组创面愈合过程中SDF-1α基因的表达规律
新生组和2周龄组小鼠皮肤均构成性表达SDF-1 mRNA。新生组的SDF-1 mRNA在创伤后表达呈逐渐增加趋势,于第3天表达达峰值,显著高于正常皮肤(P<0.05);2周龄组的SDF-1 mRNA在创伤后第1天表达即增加,于第2天表达达峰值(P<0.05),在第3天时表达虽减少,但仍高于正常皮肤(P<0.05)。与2周龄小鼠比较,新生鼠伤后第2天SDF-1 mRNA表达显著下降(P<0.05),而第3天显著升高(P<0.05)(图3,表1)。
2.4 不同浓度TNF-α和IL-1对成纤维细胞表达SDF-1α基因的影响
成纤维细胞构成性表达SDF-1基因。低浓度(0.1 ng/ml)TNF-α和IL-1能够显著抑制成纤维细胞的SDF-1α基因表达(P<0.05),并随着浓度的增加,其抑制作用逐渐增强(图4、5,表2),相同浓度的TNF-α组和IL-1组间差异没有统计学意义(P>0.05)。
3 讨论
趋化因子是一类对细胞具有定向趋化、迁移和募集作用的细胞因子。机体组织发生创伤后,创面局部产生的趋化因子不仅能趋化白细胞以及修复细胞聚集到创面,而且还能激活这些细胞并增强它们的功能,因此,趋化因子在创伤修复中的作用十分重要。新近研究证实,SDF-1作为迄今发现的对骨髓细胞趋化效应最强的趋化因子,在组织/器官损伤修复过程中发挥着极其重要的作用[2]。目前对SDF-1在重要脏器如心、肾等损伤修复过程中的作用研究报道较多,但其在皮肤创伤愈合过程中的作用及机制尚不清楚[7,8]。为此,本实验欲探讨皮肤创伤后SDF-1在创伤愈合过程中的表达特点及其可能的作用。

图2 RT-PCR检测小鼠皮肤创伤后各时间点创面组织SDF-1 mRNA的表达Fig.2 Semiquantitative analyses of SDF-1 gene amplifications at different time after trauma by RT-PCR
SDF-1是以成纤维细胞和内皮细胞为主的基质细胞合成的趋化因子,并非由炎症细胞分泌表达,在多种器官中持续性表达,SDF-1有6种同分异构体:SDF-1、SDF-1β、SDF-1γ、SDF-1δ、SDF-1ε及 SDF-1φ,但它们在功能上没有显著差异,并且SDF-1α是其在体内的主要存在形式[12],因此,本实验主要观察了SDF-1在创面的表达特征。本实验结果显示,正常皮肤组织构成性表达SDF-1,主要表达于表皮基底层和真皮的成纤维细胞胞浆内。在皮肤创伤愈合过程中,创缘SDF-1 mRNA表达呈双峰改变,分别位于创伤后第1天和第5天。伤后第1天SDF-1 mRNA表达显著增加,很可能与皮肤组织缺损造成的局部缺血缺氧有关。Daniel等[2]研究发现,组织缺血缺氧能够快速诱导缺氧诱导因子-1α表达,而后者能够在转录水平上促进SDF-1表达上调,SDF-1表达量与缺氧程度呈明显的正相关。伤后第2、3天是创伤愈合炎症反应最为严重的阶段,大量巨噬细胞募集创面,合成分泌大量促炎细胞因子,如IL-1和 TNF-α,通常在伤后第3天创面IL-1和 TNF-α表达达峰值[13]。有研究显示,IL-1和 TNF-α能强烈抑制成纤维细胞合成和分泌SDF-1[14],由此可见,创面愈合的炎症反应期大量的炎症细胞浸润和炎症细胞因子产生,可能对SDF-1 mRNA表达呈负性调控作用,因此,这可能是伤后第3天 SDF-1 mRNA表达显著减少的重要原因。
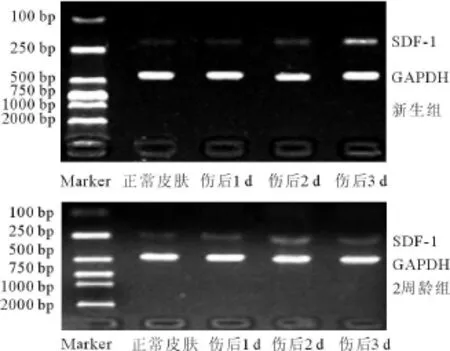
图3 皮肤创面愈合初期SDF-1和GAPDH的电泳结果Fig.3 Gel electrophoresis of SDF-1 and GAPDH during initial stage of wound healing

图4 不同浓度TNF-α作用成纤维细胞后SDF-1和 GAPDH的电泳结果Fig.4 Gel electrophoresis of SDF-1 and GAPDH expressed by fibroblast after treated with different concentrations of TNF-α
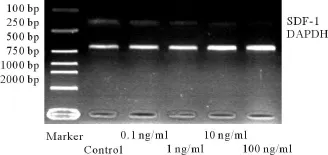
图5 不同浓度IL-1作用成纤维细胞后SDF-1和GAPDH的电泳结果Fig.5 Gel electrophoresis of SDF-1 and GAPDH expressed by fibroblast after treatment with different concentrations of IL-1
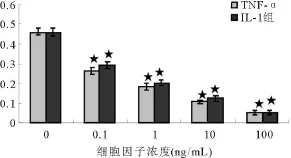
图6 不同浓度TNF-α和IL-1作用成纤维细胞后SDF-1的RT-PCR结果Fig.6 RT-PCR analyses of SDF-1 expressed by fibroblast treated with different concentrations of TNF-αand IL-1
创伤后4-14 d进入创伤愈合的增殖期,此期可见明显的肉芽组织形成,大量的新生血管形成使局部缺氧得以改善,同时可见大量增生的成纤维细胞。此时新生血管内皮细胞和成纤维细胞构成了肉芽组织的主要细胞成分,而它们均是SDF-1的重要来源细胞[2,14]。这可能是伤后第5天出现SDF-1表达再次达峰值的主要原因,随后一直维持在较高水平。
本实验还观察了不同年龄小鼠皮肤创伤后SDF-1 mRNA的表达变化,结果显示:不同年龄小鼠皮肤创面的SDF-1 mRNA表达规律不同,特别是在伤后前3天的炎症反应期,与成年小鼠完全不同,新生组和2周龄组小鼠皮肤SDF-1 mRNA表达水平是增加。以往研究认为,新生小鼠免疫系统发育尚未成熟,炎症细胞聚集功能不足,缺少炎症介质,创面的炎症反应(2-3 d)程度低[15,16],提示创面的炎症反应程度可能对SDF-1 mRNA表达产生重要影响,新生鼠和2周龄组小鼠创面低炎症反应状态可能是SDF-1 mRNA表达持续高水平的原因,进一步提示炎症反应对SDF-1 mRNA表达起着负调节作用。
成纤维细胞是创伤愈合和纤维化瘢痕形成的主要效应细胞,不仅是损伤局部分泌的细胞因子和生长因子作用的主要靶细胞,也是一些细胞因子和SDF-1的重要来源。我们通过体外实验观察促炎细胞因子TNF-α和IL-1对于小鼠皮肤成纤维细胞SDF-1 mRNA表达的影响,结果显示:炎性细胞因子能够强烈抑制成纤维细胞SDF-1 mRNA表达,并且随着细胞因子浓度的增加,其抑制作用显著增强。本实验结果与文献报道是一致的[17]。
SDF-1是一个与新生血管生成以及促进干细胞募集密切相关的趋化因子[6,7],而炎性细胞因子对其表达具有负调控作用,炎症反应越重,SDF-1合成分泌越少,将严重影响创面愈合速度。这可能是感染创面愈合速度慢和愈合不良的重要原因之一。抑制创面炎症反应,促进SDF-1合成分泌或外源性给予SDF-1治疗,可能成为提高创面愈合速度、改善创面愈合质量的新的治疗手段,也暗示促炎细胞因子对于创面愈合与组织修复是不利的。
已有研究显示SDF-1不仅是第一个报道的对人类CD34+祖细胞有强烈趋化作用的CXC趋化因子,也是参与新生血管形成的关键细胞因子[14,18]。有研究显示,糖尿病小鼠皮肤创面的SDF-1表达减少,循环中的内皮祖细胞数目显著减少,其增殖、黏附和形成血管的能力减弱,导致创面毛细血管密度显著降低,即使增加外周循环的内皮祖细胞数目,这些细胞也不能有效地到达到皮肤创面,但是增加创面的SDF-1表达,能显著增加内皮祖细胞募集到达创面,参与新血管形成,加速创面愈合,改善愈合质量[19,20]。本研究结果提示,SDF-1参与了皮肤创伤愈合的炎症期和增生期,其在创面的表达特征提示炎症反应(炎性细胞和炎性细胞因子)可能抑制SDF-1表达,其结果可能干扰创面新生血管形成,抑制骨髓来源的修复细胞在创面的募集,从而影响修复速度和愈合质量,造成延迟愈合或愈合不良。因此,外源性SDF-1对创面愈合尤其是难愈创面可能具有潜在的临床应用价值。
[1]Mescher A L,Neff AW.Regenerative capacity and the developing immune sy stem[J].Adv Biochem Eng Biotechnol,2005,93:39-66.
[2]Ceradini JD,Kulkarni AR,Callaghan MJ,et al.Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1[J].Nat Med,2004,10(8):858-864.
[3]Aiuti A,Webb IJ,Bleul C,et al.The chemokine SDF-1 is a chemoattractant for human CD34+hematopoietic progenitor cells and provides a new mechanism to ex plain the mobilization of CD34+progenitors to peripheral blood[J].J Ex p Med,1997,185(1):111-120.
[4]Ratajczak MZ,Zuba-Surma E,Kucia M,et al.The pleiotropic effects of the SDF-1-CXCR4 axis in organogenesis,regeneration and tumorigenesis[J].Leukemia,2006,20(11):1915-1924.
[5]Kucia M,Reca R,Miekus K,et al.Trafficking of normal stem cells and metastasis of cancer stem cells involve similar mechanisms:pivotal role of the SDF-1-CXCR4 axis[J].Stem Cells,2005,23(7):879-894.
[6]Jiang YP,Wu XH,Xing HY,et al.Role of CXCL12 in metastasis of human ovarian cancer[J].Chin Med J(Engl),2007,120(14):1251-1255.
[7]Tang YL,Qian K,Zhang YC,et al.M obilizing of haematopoietic stem cells to ischemic myocardium by plasmid mediated stromal-cell-derived factor-1α(SDF-1α)treatment[J].Regul Pept,2005,125(1-3):1-8.
[8]Tö gel F,Isaac J,Hu Z,et al.Renal SDF-1 signals mobilization and homing of CXCR4 positive cells to the kidney after ischemic injury[J].Kidney Int,2005,67(5):1772-1784.
[9]Kucia M,Ratajczak J,Ratajczak MZ.Bone marrow as a source of circulating CXCR4+tissue-committed stem cells[J].Biol Cell,2005,97(2):133-146.
[10]Magda K,Ratajczak J,Reca R,et al.Tissue-specific muscle,neural and liver stem/progenitor cells reside in the bone marrow,respond to an SDF-1 gradient and are mobilized into peripheral blood during stress and tissue injury[J].Blood Cells Mol Dis,2004,32(1):52-57.
[11]Falanga V.Wound healing and its impairment in the diabetic foot[J].Lancet,2005,66(9498):1736-1743.
[12]林建银,郑志竑,章涛,等.分子医学技术[M].北京:科学出版社,2000:49-51.
[13]Efron PA,Moldawer LL.Cy tokines and wound healing:the role of cytokine and anticy tokine therapy in the repair response[J].J Burn Care Rehabil,2004,25:149-160.
[14]Yu L,Cecil J,Peng SB,et al.Identification and expression of novel isoforms of human stromal cell-derived factor 1[J].Gene,2006,374:174-179.
[15]Adzick NS,Lorenz HP.Cells,matrix,growth factors and the surgeon:the biology of scarless wound repair[J].Ann Surg,1994,220(1):10-18.
[16]Olutoye OO,Yager DR,Cohen IK,et al.Lower cytokine release by fetal porcine platelets:a possible explaination for reduced inflammation after fetal wounding[J].J Pediatr Surg,1996,31(1):91-95.
[17]Fedyk ER,Jones D,Critchley HO,et al.Expression of stromal-derived factor-1 is decreased by IL-1 and TNF in dermal wound healing[J].J Immunol,2001,166(9):5749-5754.
[18]Gallagher KA,Liu ZJ,Xiao M,et al.Diabetic impairments in NO-mediated endothelial progenitorcell mobilization and homing are reversed by hyperoxia and SDF-1 alpha[J].J Clin Invest,2007,117(5):1249-1259.
[19]Gallagher KA,Liu ZJ,Xiao M,et al.Diabetic impairments in NO-mediated endothelial progenitorcell mobilization and homing are reversed by hyperoxia and SDF-1 alpha[J].J Clin Invest,2007,117(5):1249-1259.
[20]Stellos K,Langer H,Daub K,et al.Platelet-derived stromal cell-derived factor-1 regulates adhesion and promotes differentiation of human CD34+cells to endothelial progenitor cells[J].Circulation,2008,117(2):206-215.
猜你喜欢
动物营养学报(2022年11期)2022-12-08
特产研究(2021年6期)2021-12-08
昆明医科大学学报(2021年8期)2021-08-13
云南医药(2021年3期)2021-07-21
河南农业科学(2019年9期)2019-09-24
农村百事通(2016年6期)2016-05-14
中国医药生物技术(2015年4期)2015-12-26
中国现代医学杂志(2015年26期)2015-12-23
中国当代医药(2015年33期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06
