
CALHM1与阿尔茨海默病
2010-05-31 03:41:44孙素霞李文军
中国老年学杂志 2010年11期
孙素霞 李文军 邹 飞
(南方医科大学公共卫生与热带医学学院,广东 广州 510515)
阿尔茨海默病(Alzheimer′s disease,AD)作为老年性痴呆的主要类型,是老年人常见的中枢神经系统退行性疾病,临床表现为持续性、进行性的记忆、认知障碍。显著的病理特征是以 β-淀粉样蛋白(amyloid-β,Aβ)为核心成分的老年斑(senile plaques,SP)和以异常过度磷酸化的 tau蛋白为核心成分的神经纤维缠结(neurofibrillary tangles,NFTs)〔1,2〕。 AD的病因复杂,涉及遗传、环境、代谢、病毒感染等多种因素,其中遗传因素是主要原因之一。目前已确定与AD相关的基因有 4种,分别是淀粉样前体蛋白(amyloid precursor protein,APP)基因、早老素 1(presenilin 1,PS1)基因、早老素 2(presenilin 2,PS2)基因和载脂蛋白 E(apolipoprotein E,ApoE)基因。基于分子生物学及遗传学的研究表明,上述 4种基因只能解释AD中 50%与其相关,提示还有其他与 AD相关的基因〔3,4〕。最近发现一种新的基因-钙平衡调节基因 1(calcium homeostasis modulator 1,CALHM1),该基因可通过调控细胞内 Ca2+浓度来调节 Aβ的生成,从而促进 AD的发生〔5〕。本文主要阐述 CALHM1基因的特性及其与 AD发生之间的关系。
1 CALHM1特性
Dreses-Werringloer等利用 TissueInfo方法筛选到位于海马区、与 AD发生相关的基因 -FAM26C,并为其命名为 CALHM1。CALHM1基因定位于 10号染色体 q24.33位点,编码一个开放阅读框架(open reading frame,ORF)、包含四个疏水区和两个N端糖基化(分别位于 N74和 N140位点)序列。CALHM1基因表达的糖基化蛋白主要表达于成人脑组织中,其他组织器官表达量非常低或未检出。免疫荧光检测结果显示 CALHM1蛋白主要定位于内质网膜和细胞膜〔5〕。
CALHM1蛋白单体不能构成功能性的离子通道,但可在细胞内以非还原状态形成二聚体和三聚体,进而构成离子通道的孔径。CALHM1蛋白与 N-甲基 D-门冬氨酸(N-methy-D-aspatrate,NMDA)受体非常相似,通过比较他们的序列,发现二者都包含一个高度保守的、位于C端第二个疏水区的N72位点,该位点发生突变后,CALHM1蛋白形成的通道对Ca2+的通透性将会被抑制。在爪蟾卵母细胞中发现CALHM1蛋白形成的通道可渗透 Na+,在中国仓鼠卵巢细胞(CHO)中发现,CALHM1蛋白形成一种构成性的Ca2+选择性阳离子通道,这些结果表明CALHM1是一种与其他 Ca2+通道不同的、新型的 Ca2+通道〔5〕。
2 CALHM1可调控胞质中Ca2+浓度
在结构上 CALHM1与 NMDA受体存在许多相似性,二者都包含三次跨膜区和一个回流环,这些结构可帮助他们形成离子通道的孔径。因为部分 NMDA受体是一种膜蛋白,可介导Ca2+通过细胞膜,调节胞内 Ca2+浓度,所以推测 CALHM1也可能形成细胞质膜和内质网膜上的 Ca2+通道,控制胞质中 Ca2+的水平,Dreses-Werringloer等研究的结果证明该假设存在〔5〕。过表达胞内 CALHM1蛋白可促进胞外Ca2+流入胞内,通过使用相关通道工具药的研究结果显示,通过 CALHM1的胞外Ca2+内流不能被已知的钙通道的阻断剂(如:Nifedipine,SNX-482,DTL等)所阻断,但能被非特异性的钙通道阻断剂(Co2+,Ni2+)所阻断;证明 CALHM1可在细胞膜上形成离子通道介导Ca2+内流,而且 CALHM1所形成的离子通道有别于以前所知的电压门控钙离子通道(voltage gated Ca2+channel,VGCC)和钙池调控钙离子进入(store-operated Ca2+entry,SOCE)〔5,6〕。
3 细胞内Ca2+浓度与 AD发生之间的关系
早在 1987年,就有研究认为细胞内 Ca2+稳态失衡与AD发生之间存在一定关系〔7〕。随着研究的深入,多数学者认为细胞内Ca2+稳态失衡主要通过两个方面促进AD的发生,一方面Ca2+稳态失衡可影响与 AD发生密切相关的 Aβ的产生和代谢,另一方面Ca2+稳态失衡可影响tau蛋白的产生和代谢;反之,这些与AD发生相关的病理蛋白的过度积聚又可促进细胞内Ca2+稳态的进一步失衡,进而促进神经元功能的进一步紊乱和神经元的退行性疾病,诱导 AD的发生。随后研究进一步证实了该假设,如 PS1/PS2基因的突变可影响内质网中 Ca2+的释放和胞外 Ca2+通过钙池操纵性钙通道(store-operated channels,SOC)进入细胞内,从而诱导家族性AD(familial AD,FAD)的发生〔8〕。尽管缺乏 AD病人脑细胞中确实存在 Ca2+稳态失衡的直接证据,但病人尸体的检验结果证明脑细胞中钙依赖的蛋白酶含量显著增加〔9〕。
4 CALHM1与胞内 Ca2+浓度及 AD发生之间的关系
随着 AD病因的研究,细胞内 Ca2+稳态失衡诱导Aβ增加的假说得到了越来越多的证据,该假说认为,AD是一种由于基因缺陷直接或间接改变APP基因表达或其酶解过程从而影响Aβ聚集稳定性的病理综合征,Aβ产生与清除之间的平衡逐渐改变,聚集态的 Aβ积累形成 SP和过度磷酸化 tau蛋白为核心的 NFTs〔10〕。在已发现的 4个 AD基因中,APP、PS1、PS2的变异都导致 Aβ生成增加〔11,12〕,载脂蛋白(Apo)E4则影响 Aβ的沉积。细胞中 Ca2+浓度也可显著影响 APP的加工过程,促进Aβ的生成〔13〕。在 N2a细胞中过表达 CALHM1可显著抑制细胞外 Aβ的产生,同时增加 sAPPα的含量,但不影响 APP。
对晚发型 AD(LOAD)病人和正常人的基因多态性研究发现〔5〕,CALHM1基因的单核苷酸序列存在两个非同义突变rs2986017(+394 C/T;P86L)和rs17853566(+927 C/A;H264N),其中 rs17853566多态性是指氨基酸序列中的第 86位点的脯氨酸被亮氨酸所取代,导致CALHM1蛋白功能丢失,减少细胞膜对 Ca2+的渗透性,胞内 Ca2+浓度下降,进而影响 APP的加工过程,促进Aβ的产生和聚集,最终诱导AD的发生。病例对照研究结果显示,若有一个CALHM1副本可使 AD的发病率由 20%增加到 44%,若是有两个 CALHM1副本则发生AD的可能性增加为 77%。其中 rs17853566多态性在 AD病人与对照组间差异没有显著性。
相对于细胞外和内质网等钙库,胞质中Ca2+浓度相对较低,胞质中 Ca2+浓度的稳定受控于胞膜上的 VGCC、SOCE、CALHM1和 Ca-ATP酶通道和内质网膜上的钙泵(SERCA)、兰尼定受体(RyR)、三磷酸肌醇受体(IP3R)介导的受体调控的钙通道(receptor operated calcium channel,ROC)和 CALHM1;PS蛋白发生突变后可增加 IP3R和RyR介导的内质网中Ca2+释放,并抑制 SERCA将胞质中 Ca2+泵入内质网,从而升高胞质中Ca2+浓度;CALHM1基因发生突变后功能丢失,引起内质网中Ca2+的被动释放,同样升高胞质中 Ca2+浓度。胞质中 Ca2+浓度的升高可促进 β-淀粉酶和 γ-淀粉酶对APP的剪切过程,从而增加 Aβ的生成。反之,若CALHM1基因为发生突变,Ca2+可正常通过 CALHM1通道,从而促进 α-淀粉酶对 APP的剪切过程,减少 Aβ的生成。因此,细胞内 Ca2+稳态失衡可影响 Aβ的产生和代谢,提示细胞内Ca2+稳态失衡可能是AD的病因之一〔13〕(图 1所示)。但也有文献报道,CALHM1基因的多态性与 AD发生之间不存在相互关系〔14~16〕。
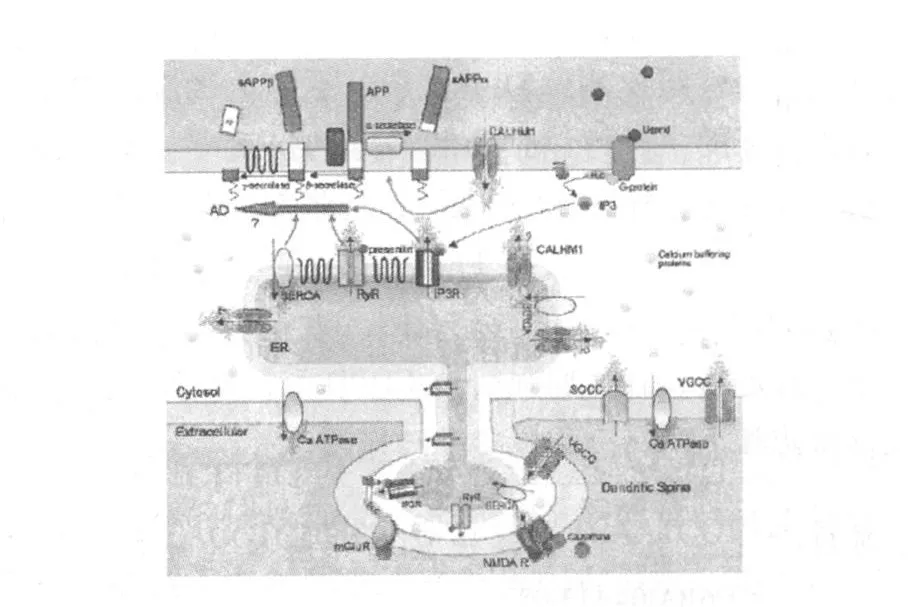
图1 钙信号通路〔13〕
5 结 语
CALHM1是表达于成人脑组织细胞膜和内质网膜、分子特性尚未完全清楚、构成性的Ca2+选择性阳离子通道。与NMDA受体具有许多相似性,可调节胞质中 Ca2+浓度,但与VGCC和SOCE等通道明显不同。CALHM1发生突变(P86L)后可通过改变胞内 Ca2+浓度调节Aβ的产生和聚集,从而增加 AD发生的危险性。这些结果为脑神经系统中Ca2+稳态失衡和APP代谢异常参与 AD的病理生理过程提供更多的证据,也提示 CALHM1是否可作为一种潜在的、诊断和治疗AD的分子靶点。
1 Mattson MP.Pathways towards and away from Alzheimer′s disease〔J〕.Nature,2004;430(7000):631-9.
2 Hardy J,Selkoe D J.The amyloid hypothesisof Alzheimer′s disease:progress and problems on the road to therapeutics〔J〕.Science,2002;297(5800):353-6.
3 Serretti A,Olgiati P,De Ronchi D.Genetics of Alzheimer′s disease.A rapidly evolving field〔J〕.J Alzheimers Dis,2007;12(1):73-92.
4 Cacabelos R,Fernandez-Novoa L,Lombardi V,et al.Molecular genetics of Alzheimer′s diseaseand aging〔J〕.Methods Find Exp Clin Pharmacol,2005(Suppl A);27:1-573.
5 Dreses-Werringloer U,Lambert JC,Vingtdeux V,et al.A Polymorphism in c ALHM1 influences Ca2+Homeostasis,Aβ Levels,and alzheimer′s disease risk〔J〕.Cell,2008;133(27):1149-61.
6 Tanzi RE,Bertram L.Alzheimer′s disease:the latest suspect〔J〕.Nature,2008;454(7205):706-8.
7 Khachaturian ZS.Hypothesis on theregulation of cytosol calcium concentration and the aging brain〔J〕.Neurobiol Aging,1987;8(4):345-6.
8 LaFerla FM.Calcium dyshomeostasis and intracellular signalling in Alzheimer′s disease〔J〕.Nat Rev Neurosci,2002;3(11):862-72.
9 Green KN,Smith IF,Laferla FM.Role of calcium in the pathogenesis of Alzheimer′s disease and transgenic models〔J〕.Subcell Biochem,2007;45:507-21.
10 Bertram L,McQueen MB,Mullin K,et al.Systematic meta-analyses of Alzheimer disease genetic association studies:the AlzGene database〔J〕.Nat Genet,2007;39(1):17-23.
11 Citron M,Westaway D,Xia W,et al.Mutant presenilins of Alzheimer′s disease increase production of 42-residue amyloid beta-protein in both transfected cells and transgenic mice〔J〕.Nat Med,1997;3(1):67-72.
12 Scheuner D,Eckman C,Jensen M,et al.Secreted amyloid beta-protein similar to that in the senile plaques of Alzheimer′s disease is increased in vivo by the presenilin 1 and 2 and APPmutations linked to familial Alzheimer#s disease〔J〕.Nat Med,1996;2(8):864-70.
13 Green KN,Laferlal FM.Linking calcium to Abeta and Alzheimer′s disease〔J〕.Neuron,2008;59(2):190-4.
14 Sleegers K,Brouwers N,Bettens K,et al.No association between CALHM1 and risk for Alzheimer dementia in a Belgian population〔J〕.Hum Mutat,2009;30(4):570-4.
15 Minster RL,Demirci FY,DeKosky ST,et al.No association between CALHM1 variation and risk of Alzheimer disease〔J〕.Hum Mutat,2009;30(4):566-9.
16 Bertram L,Schjeide BM,Hooli B,et al.No association between CALHM1 and Alzheimer′s disease risk〔J〕.Cell,2008;135(6):993-4.
猜你喜欢
世界科学技术-中医药现代化(2022年2期)2022-05-25 13:17:14
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:31:36
传染病信息(2021年6期)2021-02-12 01:52:08
中成药(2017年12期)2018-01-19 02:06:27
福建轻纺(2017年12期)2017-04-10 12:56:39
天津医药(2016年9期)2016-10-20 03:19:40
中国老年学杂志(2015年16期)2015-03-05 06:19:02
癌变·畸变·突变(2015年4期)2015-02-27 06:15:22
现代检验医学杂志(2014年3期)2014-02-02 02:42:21
西南军医(2014年6期)2014-01-22 06:57:43
