
结核分枝杆菌热休克蛋白GroEL1的研究进展
2010-01-26 06:38何磊张鹭彭超王洪海
微生物与感染 2010年3期
何磊,张鹭,彭超,王洪海
复旦大学生命科学学院遗传学研究所遗传工程国家重点实验室,上海 200433
各种生物有机体,从低等的大肠埃希菌(Escherichiacoli,E.coli)、酵母(Saccharomycescerevisiae)、果蝇(Drosophilamelanogaster)到高等的植物、哺乳动物,在受周围环境不利因素影响下,会通过快速、短暂的细胞代谢调节产生一系列适应性改变,在此期间细胞内一些正常基因的表达受抑制,某些特殊基因被激活,表现为暂时性下调或选择性上调表达一组蛋白,这一现象称为热休克反应。该反应本质上是细胞对外界作出的一种自身保护性反应,所产生的蛋白就是热休克蛋白(heat shock protein, HSP)。HSP是一种高度保守且广泛表达的蛋白,最早在高温培养果蝇时发现。正常情况下,HSP作为新生蛋白、非正常折叠蛋白和突变蛋白的分子伴侣,阻止蛋白质变性和聚集,协助蛋白质折叠、伸展、组装和转运,抑制错误折叠的蛋白质分泌,并参与细胞内对应激刺激的自我保护[1]。
HSP按相对分子质量大小可分为不同家族,主要划分为泛素 ( ubiqutin )、HSP20、HSP60、HSP70、HSP90及大分子HSP家族。其中HSP60 和 HSP70 作为HSP的2个重要家族,具有高度的物种保守性,但不同家族的HSP无明显序列同源性。HSP70已被深入研究,而 HSP60 的研究相对较少。HSP60在核酸序列及蛋白质组成上具有高度的保守性。不同生物间HSP60具有50%~90%同源性。原核生物中 HSP60 类似物是大肠埃希菌的伴侣蛋白 GroEL。GroEL伴侣分子在生物体内帮助折叠10%~15%细胞内蛋白[2,3]。虽然在大部分细菌中只有1组GroEL基因,但结核分枝杆菌(Mycobacteriumtuberculosis)基因组携带2组groEL基因,即groEL1和groEL2,分别编码GroEL1(Hsp60、Cpn60.1)和 GroEL2 (Hsp65、Cpn60.2)蛋白。结核分枝杆菌GroEL1和GroEL2具有61%同源性,与大肠埃希菌分别有53%和59%同源性,与人Hsp60也有47%同源性[4]。
结核分枝杆菌中GroEL2具有20多个 T细胞抗原决定簇,其中1个是结核分枝杆菌和牛型结核分枝杆菌(Mycobacteriumbovis)特异性的,具有很好的免疫保护与免疫治疗作用[5],是机体对抗结核分枝杆菌入侵的重要免疫保护性抗原之一,为细胞生存所必需的。研究发现,结核分枝杆菌感染的小鼠化学治疗后接种groEL2 DNA,能清除残余细菌,提示GroEL2不仅可作为预防性疫苗的候选组分,也可作为治疗性疫苗的候选组分[6,7]。
GroEL1蛋白不是细胞生存所必需的,而是一种高度保守的细胞内抗原。机体感染结核分枝杆菌后会产生针对GroEL1的交叉免疫反应,可引起实验动物自身免疫性疾病。有研究显示,GroEL1比GroEL2具有更强诱导细胞因子产生的能力,使人外周血单核细胞(peripheral blood mononuclear cell,PBMC)产生促炎症反应细胞因子,如白细胞介素1β(interleukin 1β,IL-1β)、肿瘤坏死因子α(tumor necrosis factor α,TNF α)、IL-6、IL-8、IL-10、IL-12、粒细胞-巨噬细胞集落刺激因子(granulocyte-macrophage colony-stimulating factor,GM-CSF),但不能刺激产生保护性细胞因子γ干扰素(interferon γ,IFN γ)和 IL-4[8]。这种分子伴侣和免疫优势抗原的双重功能,以及在自身免疫性疾病中的作用,使得GroEL1越来越受到广泛关注。
1 结核分枝杆菌GroEL1的发现
GroEL也称HSP60,是典型的分子伴侣。人们对分子伴侣及其在蛋白折叠中功能的了解来自于对这种蛋白及其共伴侣分子HSP10(在原核生物中为GroES)的研究。GroES称为辅分子伴侣(co-chaperonin),因为GroEL在指导折叠过程中起重要作用,而GroES 辅助其活性功能。
在大肠埃希菌中,由于GroEL/GroES伴侣系统对至少13种重要蛋白的折叠是必需的,所以这些寡聚体蛋白对生长极其重要(GroEL是四聚体,GroES是七聚体)[9]。大部分细菌在单个操纵子中只有1份groES和groEL基因拷贝[10]。然而,基因组序列分析发现,大约15%的细菌拥有不止1份groEL基因,这些细菌主要是放线菌,结核分枝杆菌也属此类。GroEL2在很久前就因其强免疫原性而被关注,被称之为普遍抗原[11]。1993年,Kong等发现一个同源基因,命名为groEL1[12]。结核分枝杆菌中groEL1基因位于groES基因下游,表明它编码这种生物主要的60kD蛋白(图1)。
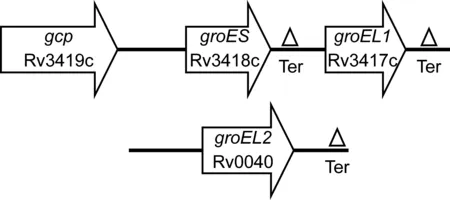
groES, codes for the Cpn10 antigen (HSP10);groEL1, codes for the Cpn60.1 antigen (HSP60);groEL2, codes for the Cpn60.2 antigen (HSP65).
图1groES-groEL1顺式操纵子和groEL2基因在基因组上的结构[13]
Fig.1GenomicorganizationofthegroES-groEL1operonandthegroEL2gene
2 结核分枝杆菌 GroEL1独特的结构特点
通过对结核分枝杆菌GroEL结构的测定,研究者发现GroEL以二聚体形式存在[14]。每个亚单位由顶部(apical domain)、赤道部(equatorial domain)和中间部(intermediate domain)3个区域组成。GroEL A链由447个氨基酸残基组成,B链由445个残基组成,拥有与大肠埃希菌相似的折叠。在GroEL表面存在疏水区。GroEL1的面积比GroEL2大,赤道部出现一个疏水区与N端残基电子密度有关。巧妙的构象变化影响不对称单位中所有分子N端残基螺旋的方向,可保护赤道区的疏水残基[14]。
结核分枝杆菌GroEL的2个蛋白具有如下特点:低序列性的寡聚体形式而非四聚体,非常弱的ATP酶活性,在底物折叠方面非常弱的活性[15]。因此,人们开始怀疑GroEL的2个蛋白是否具有助折叠功能,质疑被感染宿主中结核分枝杆菌GroEL蛋白生物功能的本质[16]。
3 结核分枝杆菌GroEL1的功能
3.1 分子伴侣功能
许多分子伴侣是HSP家族的成员。分子伴侣通过结合和释放的方式来识别非天然状态的蛋白,帮助变性、不可溶的凝聚蛋白正确折叠,重新恢复天然构象, 从而提高细胞的应急耐受能力[17]。groEL1和groES编码基因共同排列于groESL操纵子上,而groEL2单独排列于染色体上。所有分枝杆菌的GroEL在热休克、氧化应激反应、巨噬细胞感染时大量表达,比平时增加1%~10%[18-20]。
一般的细菌如大肠埃希菌在行使分子伴侣功能时均需要结合其辅助蛋白GroES和水解ATP供能。分枝杆菌却不需要辅助蛋白和ATP的协助即能完成蛋白质重新折叠、组装及转运,通过赤道部疏水区结合未折叠的蛋白,防止错误折叠和聚集。辅助蛋白折叠需要耗能而使细菌新陈代谢缓慢,结核分枝杆菌改变了以往蛋白质折叠需要耗能的途径,将ATP提供给更需要的环节。在柠檬酸合成酶体外折叠的实验中证实,GroEL具有促进其重新折叠的能力,且这种能力不受辅助蛋白和ATP的影响[21]。
3.2 抗原
对致死率很高的结核病而言,GroEL1可作为一种接种抗原使用,并有望成为治疗结核病的一种特效药。GroEL1作为一种免疫刺激物,能刺激机体产生一定效价的抗体。通过双向电泳和质谱鉴定所得蛋白质组分分析结果表明,在分别被牛型结核分枝杆菌和结核分枝杆菌感染后,众多HSP(包括GroEL1)在宿主巨噬细胞中表达量增加。已确认HSP有细胞间信号转导活性[22,23],可能促成这种免疫原性在感染宿主中高表达,不同种类HSP高度的序列同源性也导致它们之间存在交叉反应的抗原表位,会产生较强的交叉反应。免疫系统一旦接触特殊病原体的HSP,便可引起免疫记忆,诱导与其他生物体包括宿主HSP的交叉反应,当免疫系统再次感染时可启动快速应答。在人和小鼠模型中,GroEL1是体液和T细胞应答的靶点[24]。在结核病、麻风病患者和感染结核的小鼠体内可检测到GroEL1特异性抗体[25,26]。大约20%分枝杆菌反应性CD4+αβT细胞能识别结核分枝杆菌GroEL1[27]。
3.3 在抗原呈递中的作用
除直接的抗原作用,HSP对免疫系统还有间接作用。体内病原体的传染过程会经过一次热休克反应,引起HSP含量增加,这些HSP被加工和作为外源因子可结合大量细胞内蛋白降解产生的肽。GroEL 作为HSP的一员,也会与这些肽段结合,从而形成GroEL-肽复合物。这些复合物被抗原呈递细胞加工处理之后,肽段被传递到主要组织相容性复合物 (major histocompatibility complex,MHC)编码的Ⅰ类或Ⅱ类分子,随后这些MHC 多肽复合物被 T细胞识别。CD4(辅助性T细胞)和CD8(杀伤性T 细胞)等多种不同类型的 T 细胞识别HSP决定簇,有利于病原体清除[28]。
新近研究显示,用结核分枝杆菌GroEL1转染肿瘤小鼠,可减少致肿瘤性细胞。GroEL1有呈递肿瘤免疫显性抗原到细胞表面的作用[29],显示HSP有呈递和携带抗原的能力。
4 新功能的发现及研究进展
4.1 蛋白伴侣分子
结核分枝杆菌中GroEL2是细胞生存所必需的,但无论是在肉汤培养基还是巨噬细胞中,groEL1突变体与野生型菌株都没有区别,表明GroEL1对细胞生存是非必需的。但鼠感染研究表明,无论是在小鼠还是豚鼠中,groEL1突变体都不能产生肉芽肿炎症,这与groEL1突变引起细胞因子表达减少有关[30]。因此,GroEL1可能是结核分枝杆菌感染中控制细胞因子依赖性肉芽肿产生的关键因子。也就是说,GroEL1激活原发性感染的细胞因子产物,维持感染的疾病状态。然而,这种免疫调控功能是否代表分枝杆菌属中GroEL1的主要作用仍悬而未决,因为这种蛋白也存在于非典型性分枝杆菌如耻垢分枝杆菌(Mycobacteriumsmegmatis)或非致病性物种中。
在条件致病性物种如耻垢分枝杆菌中,groEL1包含了整合噬菌体Bxb1的attB位点[31],而且对于正常的浮游生长不可或缺。也有研究指出,GroEL1在耻垢分枝杆菌生物膜形成方面的作用:通过整合噬菌体Bxb1,耻垢分枝杆菌的groEL1基因失活,细菌呈现正常的浮游生长,但不形成成熟的生物膜。研究表明,GroEL1在KasA蛋白(一种参与分枝菌酸合成的Ⅱ型脂肪酸合成酶的关键成分)的帮助下,特别是在生物膜形成过程中,调控分枝菌酸-分枝杆菌细胞壁长链脂肪酸成分的合成[16]。虽然结核分枝杆菌和耻垢分枝杆菌的GroEL1直系同源物,显示高度的保守序列,但功能却不相似。很可能分枝杆菌GroEL蛋白序列或者结构上的细微变化对其生物学活性产生了深远影响,或者蛋白可能经历了不同的翻译后修饰。
最近研究表明,GroEL1保守蛋白的磷酸化是物种特异性的。在结核分枝杆菌中,GroEL1是一种Ser/Thr 蛋白激酶的底物,可以在Thr25和Thr54位点磷酸化,其中Thr25是关键的磷酸化接受位点。环境信号可引发GroEL1磷酸化,并可能影响其生物活性。Thr25磷酸化也许是导致病原体和非病原体物种GroEL1功能不同的原因。要证实Thr25磷酸化与诱导感染动物炎症反应的联系,需拥有GroEL1(T25A)等位基因的结核分枝杆菌同源菌株[32]。
相反,耻垢分枝杆菌中GroEL1不是Ser/Thr蛋白激酶F(serine/threonine protein kinase F, PknF)的底物。GroEL1通过物理性结合KasA和SMEG4308,完成主要的翻译后修饰[16]。耻垢分枝杆菌中GroEL1(T25A)不同的磷酸化模式是否与KasA相互作用,继而影响成熟生物膜形成,有待证实[32]。
GroEL1蛋白的不同表现表明这些分子并不是统一的,不同来源的GroEL1蛋白可以表达不同的生物活性。
4.2 DNA伴侣分子
研究表明,结核分枝杆菌GroEL1蛋白在进化过程中,获得一种新功能,即以低特异性、高DNA亲和力来结合DNA的能力,从而成为核酸相关蛋白(nucleoid-associated protein,NAP)的一员。GroEL1可有效保护DNA免受DNase Ⅰ和氢氧自由基的损害。原子粒显微镜研究表明,GroEL1可将一条大的DNA浓缩为一个紧密的结构。体内实验证明,通过DNA杂交和免疫荧光图像发现纯核酸中存在GroEL1时两者会发生相互作用,说明自然环境中GroEL1和DNA可相互作用[33]。
产生这一功能的原因可能在于GroEL1底物识别顶端区域的改变导致底物特异性从多肽转变为多核苷酸。根据其二聚体而非四聚体的结构,推测这种蛋白寡聚状态的改变可能来自于细微突变,具体原因未知[34]。
5 结语
分枝杆菌GroEL1作为基因复制的产物,不仅具有原始的助蛋白折叠功能,还可作为一种重要抗原被免疫系统所识别,在结核病的防治中具有重要的应用价值,而且在不同分枝杆菌中呈现出多种不同的生物学功能。结核分枝杆菌的GroEL1作为蛋白分子伴侣,帮助控制细胞因子依赖性肉芽肿的形成[35];在耻垢分枝杆菌中,GroEL1作为蛋白分子伴侣,与分枝菌酸的生物合成有关。这对结核分枝杆菌在体内尤其在巨噬细胞内的生存有重要意义,基因敲除实验结果进一步证实了这一点。GroEL1包含了整合噬菌体Bxb1的attB位点,对于耻垢分枝杆菌正常的浮游生长是不可或缺的,且其C端富含组氨酸残基,对生物膜形成很重要。研究还发现它具有 DNA 分子伴侣功能,可保护DNA免受自由基侵害。GroEL蛋白序列或者结构上的细微变化对其功能可能产生深远的影响,致使不同来源的GroEL1蛋白表达不同的生物活性[33]。
未来的研究将对致病菌和条件致病菌GroEL蛋白的不同功能进行分析,找出产生这些差异的具体原因,从而有助于找到合适的药物靶点和(或)疫苗,为人类结核病的发生及进化机制的研究提供坚实的基础和广阔的视角。另外,致病菌GroEL1蛋白在进化过程中产生了新的功能,这种功能变化的具体原因也有待继续研究。
[1] Gething MJ, Sambrook J. Protein folding in the cell [J]. Nature, 1992, 355(1): 33-45.
[2] Ewalt KL, Hendrick JP, Houry WA, Hartl FU. In vivo observation of polypeptide flux through the bacterial chaperonin system [J]. Cell, 1997, 90(3): 491-500.
[3] Houry WA, Frishman D, Eckerskorn C, Lottspeich F, Hartl FU. Identification of in vivo substrates of the chaperonin GroEL [J]. Nature, 1999,402(6758): 147-154.
[4] Goyal K, Qamra R, Mande SC. Multiple gene duplication and rapid evolution in the groEL gene: functional implications [J]. J Mol Evol, 2006, 63(6): 781-787.
[5] Silva CL, Bonato VL, Coelho-Castelo AA, Souza1 AD, Santos1 SA, Lima1 KM, Faccioli LH, Rodrigues JM. Immunotherapy with plasmid DNA encoding mycobacterial hsp65 in association with chemotherapy is a more rapid and efficient form of treatment for tuberculosis in mice [J]. Gene Ther, 2005, 12(3): 281-287.
[6] Vordermeier HM, Lowrie DB, Hewinson RG. Improved immunogenicity of DNA vaccination with mycobacterial HSP65 against bovine tuberculosis by protein boosting [J]. Vet Microbiol, 2003, 93(4): 349-359.
[7] Yang BF, Zhao HL, Xue C, Xiong XU, Zhang W, Yao XQ, Liu Z. Recombinant heat shock protein 65 carrying hepatitis B core antigen induces HBcAg-specific CTL response [J]. Vaccine, 2007, 25(22): 4478-4486.
[8] Lewthwaite JC, Coates AR, Tormay P, Singh M, Mascagni P, Poole S, Roberts M, Sharp L, Henderson B. Mycobacterium tuberculosis chaperonin 60.1 is a more potent cytokine stimulator than chaperonin 60.2 (Hsp 65) and contains a CD14-binding domain [J]. Infect Immun, 2001, 69(12): 7349-7355.
[9] Kerner MJ, Naylor DJ, Ishihama Y, Maier T, Chang HC, Stines AP, Georgopoulos C, Frishman D, Hartl MH, Mann M, Hartl FU. Proteome-wide analysis of chaperonin-dependent protein folding in Escherichia coli [J]. Cell, 2005, 122(2): 209-220.
[10] Lund PA. Microbial molecular chaperones [J]. Adv Microb Physiol, 2001, 44: 93-140.
[11] Young DB, Ivanyi J, Cox JH, Lamb JR. The 65 kDa antigen of mycobacteria—a common bacterial protein [J]. Immunol Today, 1987, 8(7-8): 215-219.
[12] Kong TH, Coates AR, Butcher PD, Hickman CJ, Shinnick TM. Mycobacterium tuberculosis expresses two chaperonin-60 homologs [J]. Proc Natl Acad Sci USA, 1993, 90(7): 2608-2612.
[13] Aravindhan V, Christy AJ, Roy S, Ajitkumar P, Narayanan PR, Narayanan S. Mycobacterium tuberculosis groE promoter controls the expression of the bicistronic groESL1 operon and shows differential regulation under stress conditions [J]. FEMS Microbiol Lett, 2009, 292(1): 42-49.
[14] Qamra R, Mande SC. Crystal structure of the 65-kilodalton heat shock protein, chaperonin 60.2, of Mycobacterium tuberculosis [J]. J Bacteriol, 2004, 186(23): 8105-8113.
[15] Qamra R, Srinivas V, Mande SC. Mycobacterium tuberculosis GroEL homologues unusually exist as lower oligomers and retain the ability to suppress aggregation of substrate proteins [J]. J Bacteriol, 2004, 342(2): 605-617.
[16] Ojha A, Anand M, Bhatt A, Kreme L, Jacobs WR, Hatfull GF. GroEL1: a dedicated chaperone involved in mycolic acid biosynthesis during biofilm formation in mycobacteria [J]. Cell, 2005, 123(5): 861-873.
[17] Hartl FU, Martin J. Molecular chaperones in cellular protein folding [J]. Nature, 1996, 381(6583): 571-579.
[18] Stewart GR, Wernisch L, Stabler R, Mangan JA, Hinds J, Laing KG, Youn DB, Butcher PD. Dissection of the heat-shock response in Mycobacterium tuberculosis using mutants and microarrays [J]. Microbiology, 2002, 148(10): 3129-3138.
[19] Dosanjh NS, Rawat M, Chung JH, Yossef AG. Thiol specific oxidative stress response in Mycobacteria [J]. FEMS Microbiol Lett, 2005, 249(1): 87-94.
[20] Ellis RJ. Do molecular chaperones have to be proteins [J]? Biochem Biophys Res Commun, 1997, 238(3): 687-692.
[21] Qamra R, Mande SC, Coates AR, Hendersonc B. The unusual chaperonins of Mycobacterium tuberculosis [J]. Tuberculosis (Edinb), 2005, 85(5-6): 385-394.
[22] Monahan IM, Betts J, Banerjee DK, Butcher PD. Differential expression of mycobacterial proteins following phagocytosis by macrophages [J]. Microbiology, 2001, 147(2): 459-471.
[23] Panayi GS, Corrigall VM, Henderson B. Stress cytokines: pivotal proteins in immune regulatory networks [J]. Curr Opin Immunol, 2004, 16(4): 531-534.
[24] Tormay P, Coates AR, Henderson B. The intercellular signaling activity of the Mycobacterium tuberculosis chaperonin 60.1 protein resides in the equatorial domain [J]. J Biol Chem, 2005, 280(14): 14272-14277.
[25] Young RA. Stress proteins and immunology [J]. Annu Rev Immunol, 1990, 8: 401-420.
[26] Young D, Lathigra R, Hendrix R, Sweetser D, Young RA. Stress proteins are immune targets in leprosy and tuberculosis [J]. Proc Natl Acad Sci USA, 1988, 85(12): 4267-4270.
[27] Hochrein H, Shortman K, Vremec D, Scott B, Hertzog P, O’Keeffe M. Differential production of IL-12, IFN-α, and IFN-γ by mouse dendritic cell subsets [J]. J Immunol, 2001, 166(9): 5448-5455.
[28] Mustafa AS, Lundin KE, Oftung F. Human T cells recognize mycobacterial heat shock proteins in the context of multiple HLA-DR molecules: studies with healthy subjects vaccinated with Mycobacterium bovis BCG and Mycobacterium leprae [J]. Infect Immun, 1993, 61(12): 5294-5301.
[29] Kaufmann SH, Vath U, Thole JE, Embden JD, Emmrich F. Enumeration of T cells reactive with Mycobacterium tuberculosis organisms and specific for the recombinant mycobacterial 64-kDa protein [J]. Eur J Immunol, 1987, 17(3): 351-357.
[30] Lamb JR, Bal V, Mendez-Samperio P, Mehlert A, So A, Rothbard J, Jindal S, Young RA,Young DB. Stress proteins may provide a link between the immune response to infection and autoimmunity [J]. Int Immunol, 1989, 1(2): 191-196.
[31] Kim AI,Ghosh P, Aaron MA, Bibb LA, Jain S, Hatfull GF. Mycobacteriophage Bxb1 integrates into the Mycobacterium smegmatis groEL1 gene [J]. Mol Microbiol, 2003(50): 463-473.
[32] Lukacs KV, Lowrie DB, Stokes RW, Colston MJ. Tumor cells transfected with a bacterial heat-shock gene lose tumorigenicity and induce protection against tumors [J]. J Exp Med, 1993, 178(1): 343-348.
[33] Basu D, Khare G, Singh S, Tyagi A, Khosla S, Mande1 SC. A novel nucleoid-associated protein of Mycobacterium tuberculosis is a sequence homolog of GroEL [J]. Nucleic Acids Res, 2009, 37(15): 4944-4954.
[34] Hendrick JP, Hartl FU. Molecular chaperone function of heat shock proteins [J]. Annu Rev Biochem, 1993, 62(2): 349-384.
[35] Bhatt A, Molle V, Besra GS, Jacobs WR, Kremer L. The Mycobacterium tuberculosis FAS-II condensing enzymes: their role in mycolic acid biosynthesis, acid-fastness, pathogenesis and in future drug development [J]. Mol Microbiol, 2007(64): 1442-1454.
猜你喜欢
海峡姐妹(2020年6期)2020-07-25
海峡姐妹(2019年12期)2020-01-14
海峡姐妹(2019年9期)2019-10-08
好日子(2018年9期)2018-10-12
特别健康(2018年4期)2018-07-03
中国医学装备(2016年6期)2016-12-01
西南医科大学学报(2015年1期)2015-08-22
医学研究杂志(2015年6期)2015-07-01
中国当代医药(2015年36期)2015-03-11
癌变·畸变·突变(2015年3期)2015-02-27
