
干旱胁迫下γ-氨基丁酸喷施对燕麦幼苗生长及叶绿素荧光参数的影响
2025-03-06 00:00:00魏臣杰李舒琦邓飞刘静任健
草地学报 2025年2期





摘要:燕麦(Avena sativa L.)作为一种世界性的重要饲草,其生长受到水分胁迫的影响和激素代谢调节。本研究利用PEG-6000模拟了中度干旱(15%)和重度干旱(25%),在不同干旱胁迫下于燕麦幼苗期喷施0.5和1 mmol·L-1 γ-氨基丁酸(γ-aminobutyric acid,GABA),探究外源GABA对干旱胁迫下燕麦幼苗生长和叶绿素荧光特性的影响。结果表明:(1)随干旱程度的加剧,燕麦幼苗的株高、叶长、叶宽、根长和地上生物量呈降低趋势,特别是重度干旱胁迫下显著降低(Plt;0.05),叶面喷施GABA后,燕麦的叶长、叶宽、根长和地上生物量则得到了显著提高(Plt;0.05);(2)干旱胁迫导致燕麦幼苗叶绿素含量、PSⅡ实际光能转换效率(Fv'/Fm'),光化学猝灭系数(Photochemical quenching coefficient,qP)和PSⅡ电子传递速率(Electron transport rate,ETR)的降低和天线热耗散百分率(Antenna thermal dissipation percentage,Hd)的升高,相反,叶面喷施GABA后,尤其是重度干旱胁迫下,叶绿素总量,Fv'/Fm',qP,ETR,Hd均显著增加(Plt;0.05),其中1 mmol·L-1的影响最为明显;(3)相关性分析表明,地上部生物量(Aboveground biomass per plant,ABP)与Fm',Fv'/Fm'呈显著正相关(Plt;0.05),根冠比(Root-shoot ratio,R-SR)却与之相反,两者均与Hd呈显著负相关(Plt;0.05)。因而,燕麦在遭受干旱的情况下,可以喷施GABA可以缓解干旱对燕麦幼苗光合和生长的抑制作用。
关键词:γ-氨基丁酸;干旱胁迫;燕麦;幼苗生长;叶绿素荧光参数
中图分类号:S512.6""""""" 文献标识码:A""""""" 文章编号:1007-0435(2025)02-0489-09
Effects of γ-aminobutyric Acid Application on the Growth and Chlorophyll Fluorescence Parameters of Oat Seedlings under Drought Stress
WEI Chen-jie1, LI Shu-qi1, DENG Fei1, LIU Jing-wen2, REN Jian1*
(1.Yunnan Agricultural University, Kunming, Yunnan Province 650201, China;
2.Yunnan Jinyu Ecological Engineering Consulting Co.Ltd, Kunming, Yunnan Province 650233,China)
Abstract:Oat (Avena sativa L.) is an important forage in the world, and its growth is affected by water stress and hormone metabolism regulation. In this study, polyethylene glycol 6000 (PEG-6000) was used to simulate moderate drought(15% PEG)and severe drought(25% PEG),and 0.5 and 1 mmol·L-1 γ-aminobutyric acid (GABA)were sprayed at the seedling stage of oats under different drought stresses, respectively. The aims were to investigate the effects of exogenous GABA on the growth and chlorophyll fluorescence characteristics of oat seedlings under drought stress. The results showed that:(1)with the aggravation of drought,the plant height,leaf length,leaf width,root length and aboveground biomass of oat seedlings decreased,especially under severe drought stress(Plt;0.05),while the leaf length,leaf width,root length and aboveground biomass of oats increased significantly after foliar application of GABA(Plt;0.05);(2)drought stress led to the decrease of chlorophyll content,PSII actual photoenergy conversion efficiency(Fv'/Fm'),photochemical quenching coefficient(qP)and PSII electron transport rate(ETR)and increase the percentage of antenna thermal dissipation percentage (Hd)of oat seedlings,on the contrary,the total amount of chlorophyll,Fv'/Fm',qP,ETR and Hd increased significantly after foliar application of GABA,especially under severe drought stress(Plt;0.05),the effect of 1 mmol·L-1 application is most pronounced;(3)correlation analysis showed that aboveground biomass per plant(ABP)was significantly positively correlated with Fm',Fv'/Fm'(Plt;0.05),while root-shoot ration(R-SR) was negatively correlated with Hd(Plt;0.05).Therefore,in the case of drought,GABA can be sprayed to alleviate the inhibition of drought on the photosynthesis and growth of oat seedlings.
Key words:γ-aminobutyric acid;Drought stress;Oats;Seedling growth;Chlorophyll fluorescence parameters
叶绿素荧光参数是植物叶片光合生理特性的重要指标,对于研究植物与环境之间关系有着重要的价值[1]。干旱作为一种长期且反复出现的极端自然气候,干旱程度的加剧会导致植物体内的水分代谢过程紊乱,植物的叶绿体结构被破坏以及叶绿素的分解,进而使叶绿素含量以及光合电子传递链传递效率降低,从表征的叶绿素荧光参数来看,下降明显[2],这在鹰嘴紫云(Astragalus cicer)幼苗中得到了进一步的证实[3];在干旱胁迫下,花生(Arachis hypogaea L.)的光合系统PSⅡ反应中心遭到破坏,花生对吸收光能后的利用能力降低[4]。而干旱不仅影响植物的叶绿素荧光参数,还使植物蒸腾速率加快、叶片萎蔫黄化、卷曲和植株低矮[5]。韩喆等[6]研究发现,干旱胁迫下伊犁绢蒿(Seriphidium transiliense)地上部分生长减缓,株高以及地上生物量降低,根长和根冠比逐渐增加。随着全球气候变暖以及干旱程度的加剧,研究植物的生存策略和适应性机制势在必行。
γ-氨基丁酸(γ-aminobutyric acid, GABA)作为一种重要的非蛋白质氨基酸,具有维持植物体内碳氮代谢平衡、作为信号分子、改善抗氧化酶系统活性、提高渗透调节物质含量等功能,对提高作物抗旱性具有重要作用[7-8]。有研究发现,喷施外源GABA可以有效地促进甘薯(Ipomoea batatas (L.) Lam.)地上部和地下部的生长发育以及干物质的积累,提高根冠比,缓解干旱对甘薯叶片的损伤[9]。GABA能够提高燕麦幼苗的抗氧化酶活性和光合能力,降低有害物质MDA和渗透压调节物质的含量[10],还可以提高辣椒(Capsicum annuum L.)叶绿素的含量,使叶片叶绿素荧光参数受胁迫的影响程度降低[11]。
燕麦(Avena sativa L.)是全球性的一年生禾本科牧草,营养丰富、适口性好、抗寒和抗旱性较强,是适合大面积推广种植的高产优质、粮饲兼用的作物之一[12-13],对全球范围内牛奶和肉类生产均具有重要意义[14]。近年来,随着人们生活水平的日益提高,对畜牧业产品的需求量增加,燕麦种植面积和区域急剧扩张[15]。然而,干旱限制了燕麦的推广和种植[16]。干旱导致燕麦株高和根冠比降低[17],叶片数减少,叶面积变窄,叶片开始失绿发黄,降低了籽粒产量及其构成因子[18]。因此,研究干旱下燕麦的反应机理,将为提高燕麦抗旱性提供理论依据。
为此,针对燕麦生长前期容易存在的干旱问题,本研究以燕麦为实验材料,利用PEG-6000模拟干旱胁迫,对叶片喷施不同浓度的GABA,研究幼苗生长和光合特性等指标,以期为利用外源GABA提高燕麦抗旱性提供理论依据。
1 材料与方法
1.1 试验材料
燕麦种子(品种:‘太阳神’)购自北京正道种业有限公司;外源GABA由国药集团化学试剂有限公司生产。
1.2 试验设计
本试验于人工气候箱内进行,白昼温度为25/20℃,每天光照和黑暗均12 h,光强为4000 lx,湿度为70%。首先选取大小一致且饱满的种子,用0.1%次氯酸钠溶液消毒15 min后蒸馏水冲洗干净,点播于珍珠岩量相同的育苗盆(19 cm×14.5 cm×11 cm)。根据燕麦的田间播种量和育苗盆的大小,试验中用的种子数量为36粒,每盆种子用量相同。2022年8月6日于人工气候箱中发芽,发芽后用Hoagland营养液培养,当平均株高为10 cm时(幼苗期)开展干旱胁迫。
试验因素为干旱胁迫和不同浓度的GABA,采用随机区组设计。干旱胁迫是利用聚乙二醇(PEG-6000)进行模拟,不同的水分梯度分别是:正常供水(PEG浓度为0)、中度干旱(15%PEG-6000溶液)和重度干旱(25%PEG-6000溶液)。试验中PEG溶解于Hoagland营养液中进行浇灌,每3 d更换一次(200 mL),对照中只含Hoagland营养液;GABA浓度梯度为:0,0.5 mmol·L-1和1 mmol·L-1,采用叶面喷施,每2 d喷施一次(15 mL),确保GABA更好附着叶片加入2滴吐温-80℃。胁迫时间为15 d。试验共有9个处理,每个处理重复3次。
1.3 测定指标
1.3.1 形态指标 每个处理随机选取10株幼苗,用卷尺测量株高、叶长、根长;游标卡尺测叶宽;单株地上生物量在烘箱内105℃杀青30 min,之后80℃烘干至恒重。
1.3.2 叶绿素含量 选取植株中部叶片,称取0.2 g后浸入10 mL 80%丙酮,在黑暗中浸提24 h。用紫外分光光度法测定470,649和665 nm下的吸光度值,并计算叶绿素a(Chlorophyll a,Chla)、叶绿素b(Chlorophyll b,Chlb),总叶绿素(Chlorophyll,Chla+b)以及叶绿素a/b的含量[19]。
1.3.3 叶绿素荧光测定 采用LI-6400-40叶绿素荧光系统,每次测定时选取无病虫害且完全展开的叶片。按照仪器操作说明并参照周艳虹等[20]的方法测定各荧光指标,测定指标包括叶片光适应下最小荧光(Fo'),最大荧光(Fm'),可变荧光(Fv'),PSⅡ最大光化学效率(Fv'/Fm'),光化学猝灭系数(qP),非光化学淬灭系数(Non-photochemical quenching,NPQ),PSII电子传递速率(PSⅡ electron transport rate,ETR),并计算叶片ΦPSⅡ天线热耗散百分率(Hd),非光化学耗散百分率(Non-photochemical dissipation percentage,Ex),光化学反应百分率(Photochemical reaction percentage,Pc)[21]。
Hd=1-Fv'/Fm'
Ex=(1-qP)×Fv'/Fm'
Pc=qP×Fv'/Fm'
1.4 数据统计分析
利用SPSS25.0统计分析软件对数据进行单因子方差分析,多重比较采用Duncan法,检验水平α=0.05,比较各个处理之间的区别;利用两因素方差分析(Two-way ANOVA)比较外源GABA和干旱的响应。用Origin2021软件绘图,Excel 365制表。
2 结果与分析
2.1 外源GABA对干旱胁迫下燕麦幼苗形态的影响
由表1可知,干旱胁迫对株高、叶长、叶宽、根长和地上生物量均有显著影响(Plt;0.05)。随着干旱程度的增加,燕麦幼苗株高、叶长、叶宽、根长和地上生物量均呈现下降趋势。与CK相比,中度、重度干旱胁迫下植株高度和叶宽分别降低23.8%,37%和17.5%,25%(Plt;0.05),且干旱胁迫之间差异显著(Plt;0.05),表明干旱胁迫对燕麦幼苗生长具有抑制作用,且随着胁迫程度增加而加剧。正常供水时,喷施GABA使燕麦幼苗株高、叶长、叶宽、根长、地上生物量呈现上升趋势。与CK相比,0.5 mmol·L-1 GABA处理下株高、叶长、根长和地上生物量分别增加了4.3%,5.4%,15.5%和4.8%,根冠比降低15.2%;1 mmol·L-1 GABA处理下燕麦幼苗株高和叶长显著增加11.4%,12.6%(Plt;0.05),表明一定浓度外源GABA能够促进燕麦幼苗生长。中度干旱胁迫下,燕麦幼苗的株高、叶长和地上生物量随GABA浓度增加呈现出先升后降的趋势,GABA浓度为0.5 mmol·L-1时达到最大值,与CK相比,0.5 mmol·L-1 GABA处理下燕麦幼苗株高和地上生物量显著增加8%,51%(Plt;0.05),根冠比显著降低32%(Plt;0.05)。1 mmol·L-1 GABA处理后,燕麦幼苗株高、叶长、叶宽、根长和地上生物量均高于CK,但未达显著水平,相反,根冠比发生了显著降低(Plt;0.05)。重度干旱胁迫下,0.5 mmol·L-1 GABA处理后燕麦幼苗株高、叶宽显著高于CK(Plt;0.05)。与CK相比,1 mmol·L-1 GABA处理下株高、根长和地上生物量有增加的趋势,但未达显著水平。说明一定浓度外源GABA能够缓解干旱对植株的生长抑制,其中0.5 mmol·L-1 GABA缓解效果最好。
2.2 外源GABA对叶绿素含量的影响
干旱胁迫显著影响了叶绿素含量(图1)(Plt;0.05)。随干旱胁迫程度的增加,叶绿素a、叶绿素b、总叶绿素含量以及叶绿素a/b呈现下降趋势,前三者均在重度干旱胁迫下达到最低值。正常供水时,外源GABA对叶绿素的影响不大。在中度干旱胁迫下,与CK相比,0.5 mmol·L-1和1 mmol·L-1 GABA处理下叶绿素a和总叶绿素含量升高,但未达显著水平;喷施1 mmol·L-1 GABA处理下叶绿素a/b显著升高(Plt;0.05)。而在重度干旱胁迫下,与CK相比,喷施0.5 mmol·L-1 GABA显著提高叶绿素a、叶绿素b和总叶绿素含量,分别提高53.5%,10.4%和42.5%(Plt;0.05),喷施1 mmol·L-1 GABA显著提高叶绿素a和总叶绿素含量,分别提高39.1%和22.0%(Plt;0.05)而在0.5 mmol·L-1和1 mmol·L-1 GABA处理下叶绿素a/b均显著升高(Plt;0.05)。就叶绿素a和整个叶绿素含量而言,干旱胁迫与GABA之间存在显著的交互作用(Plt;0.05)。结果表明:干旱胁迫使燕麦幼苗叶片叶绿素含量降低,但GABA降低了干旱胁迫的抑制作用,表现为叶绿素含量增加。
2.3 外源GABA对干旱胁迫下燕麦幼苗叶绿素荧光的影响
2.3.1 外源GABA对干旱胁迫下燕麦幼苗Fo',Fm',Fv'和Fv'/Fm'的影响 随着胁迫程度的增加,燕麦叶片的Fo',Fm',Fv'和Fv'/Fm'均逐渐降低(Plt;0.05)(图2)。中度干旱胁迫下,燕麦叶片Fo',Fm',Fv'和Fv'/Fm'分别比CK降低14.4%,26.4%,32.6%和7.7%(Plt;0.05)(图2);重度干旱胁迫Fo',Fm',Fv'和Fv'/Fm'分别比CK降低45.9%,63.5%,72.6%和25.2%(Plt;0.05)。说明干旱胁迫使燕麦幼苗叶片光系统反应中心捕获激发能效率下降,光系统功能受到抑制。
正常供水时,与CK相比,喷施GABA显著降低燕麦叶片Fo',Fm',Fv'和Fv'/Fm',0.5 mmol·L-1 GABA处理下Fo',Fm',Fv'和Fv'/Fm'分别降低了16.5%,29.6%,36.3%和8.6%;1 mmol·L-1 GABA处理下分别降低7.8%,19.4%,25.4%和8.4%(Plt;0.05)。而与CK相比,中度干旱胁迫下,1 mmol·L-1 GABA 处理下Fo',Fm',Fv'和Fv'/Fm'呈上升趋势,但未达显著水平;0.5 mmol·L-1 GABA处理下显著降低Fv'/Fm'(Plt;0.05)。说明外源GABA不能缓解中度干旱胁迫对燕麦叶片PSⅡ功能的破坏。重度干旱胁迫下,1 mmol·L-1 GABA与CK相比,燕麦叶片Fo',Fm',Fv'和Fv'/Fm'显著增加37.8%,52.5%,67.4%和11.2%(Plt;0.05),0.5 mmol·L-1 GABA作用不明显。
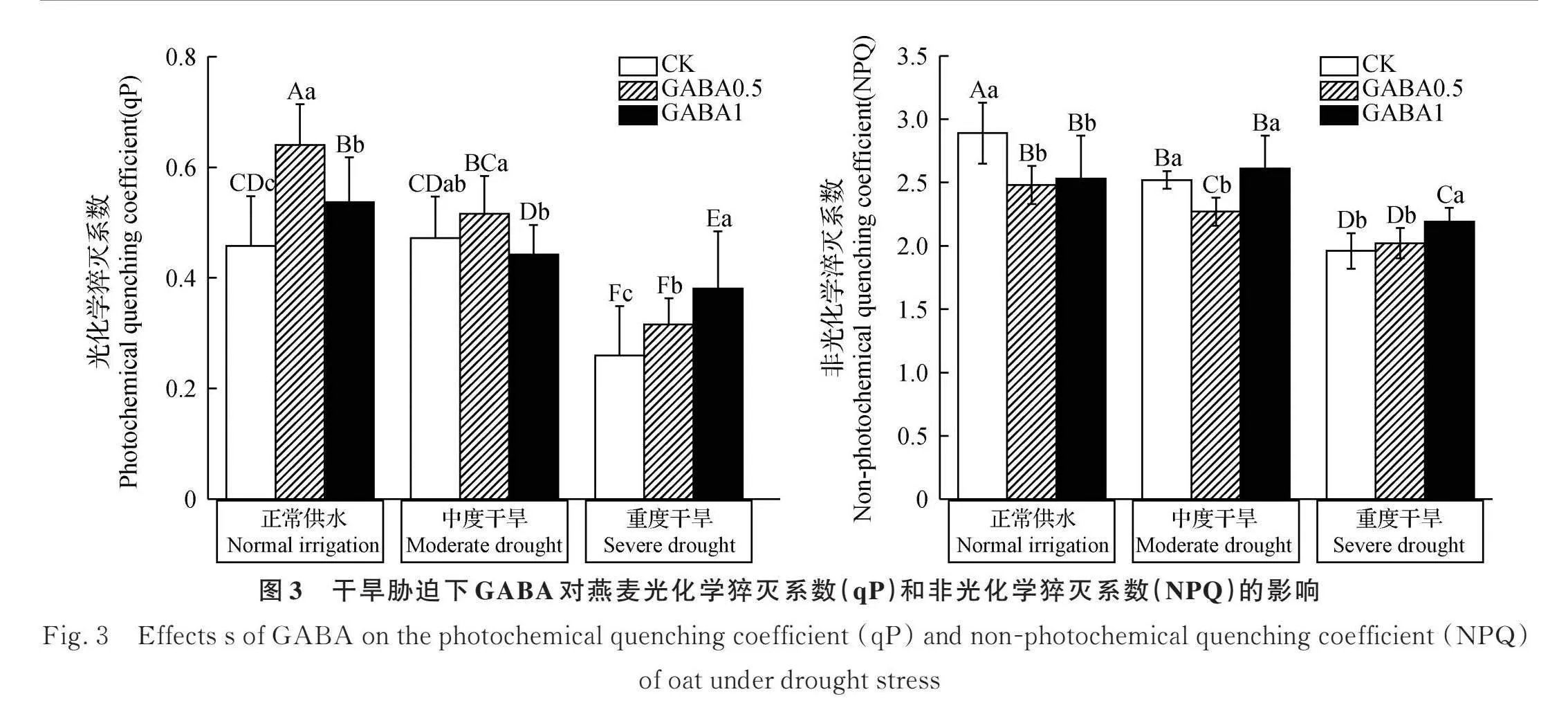
2.3.2 外源GABA对干旱胁迫下燕麦幼苗qP和NPQ的影响 光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)随着干旱程度的增加呈现相反趋势(图3)。正常供水时,与CK相比,喷施GABA可以显著提高qP(Plt;0.05),但显著降低了NPQ(Plt;0.05)。在中度干旱胁迫下,与CK相比,0.5 mmol·L-1 GABA提高了qP,但差异不显著,相反,NPQ显著降低(Plt;0.05),而1 mmol·L-1 GABA无显著效果。重度干旱胁迫下,与CK相比,喷施0.5 mmol·L-1 GABA使qP显著升高21.8%(Plt;0.05)。
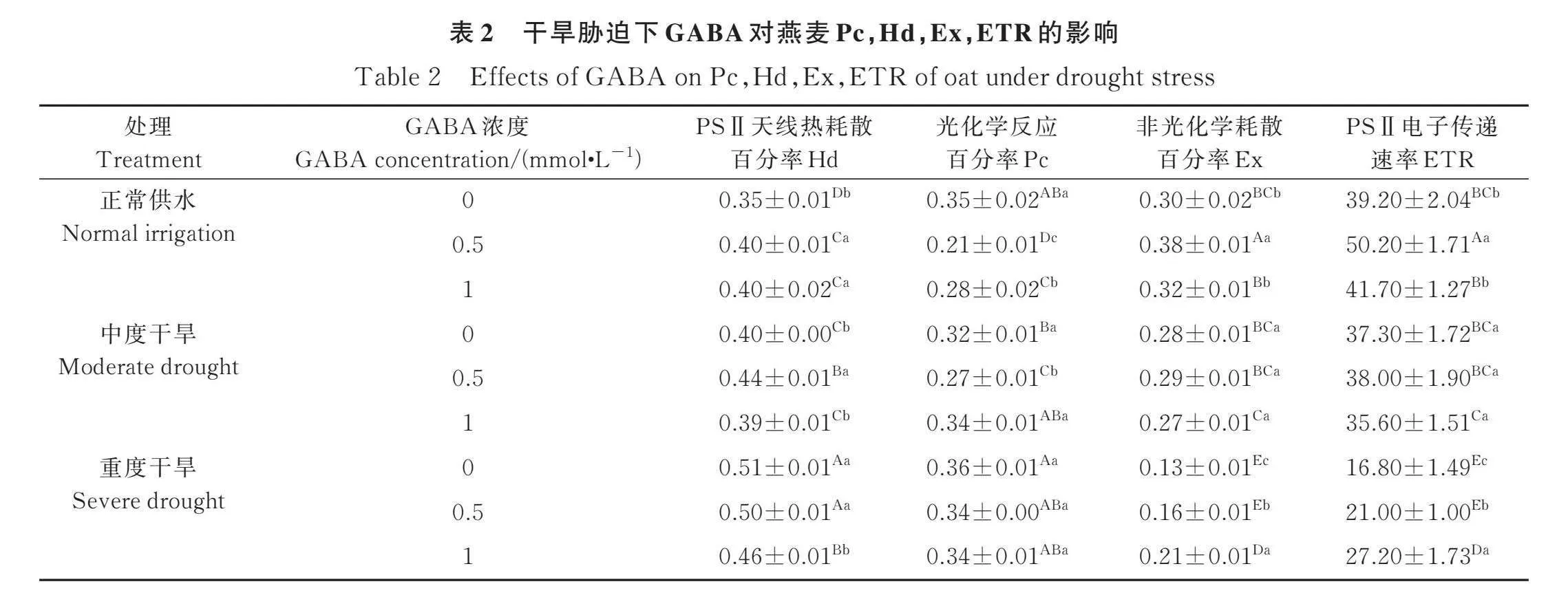
2.3.3 外源GABA对Pc,Hd,PSⅡ的Ex,ETR的影响 随着干旱程度的增加,燕麦幼苗叶片天线热耗散的百分率(Hd)呈逐渐升高趋势,中度、重度干旱胁迫分别比CK增加14.3%,45.7%(Plt;0.05)(表2)。相反,表观光合电子传递速率(ETR)和非光化学耗散百分率(Ex)呈逐渐降低的趋势,中度和重度干旱胁迫下Ex,ETR,分别比CK降低6.7%,4.8%;56.7%(Plt;0.05),57.1%(Plt;0.05)。正常供水时,与CK相比,喷施0.5 mmol·L-1 GABA显著提高Hd,Ex和ETR,Pc显著降低(Plt;0.05);喷施1 mmol·L-1 GABA 显著提高Hd,显著降低Pc(Plt;0.05)。说明正常条件下,喷施0.5 mmol·L-1 GABA能提高电子传递速率。中度干旱胁迫下,与CK相比,喷施0.5 mmol·L-1 GABA叶片Hd显著提高,Pc显著降低(Plt;0.05),喷施1 mmol·L-1 GABA效果不显著。重度干旱胁迫下,喷施GABA能够显著提高Ex和ETR,0.5 mmol·L-1和1 mmol·L-1 GABA较CK分别提高23.1%和25%;61.5%和61.9%(Plt;0.05)。说明重度干旱胁迫下外源GABA有利于促进PSⅡ反应中心的电子传递活跃,提高光合活性,缓解干旱胁迫对燕麦幼苗叶片光合功能的破坏。
2.4 外源GABA对干旱胁迫下燕麦生长与叶绿素荧光参数相关分析
对燕麦的生长指标和叶绿素荧光参数进行相关性分析(图4)。图中可知,PH与LL呈显著正相关(Plt;0.05);ABP与Fo',Fm',Fv'/Fm'呈显著正相关(Plt;0.05),与Hd呈显著负相关(Plt;0.05);除Fo'外,R-SR与Pc呈极显著正相关(Plt;0.01),与Fm',Fv', Fv'/Fm'和Hd呈显著负相关(Plt;0.05),而与qP,NPQ和ETR呈极显著负相关(Plt;0.01)。
3 讨论
干旱环境下,植物的生长受到了抑制,表现为株高、叶长和叶宽的降低,地上生物量的减少[21],外源GABA的施用影响了植物的抗旱性。有研究发现,外源GABA显著改善了干旱胁迫下番茄(Lycopersicon esculentum Mill.)、黄瓜(Cucumis sativus L.)和甜瓜(Cucumis melo L.)幼苗的根长、株高等,提高了植物应对胁迫的能力[22-24]。本试验中,干旱处理下燕麦的株高、叶长、叶宽和根长以及地上生物量显著低于CK,相反,喷施GABA后株高、叶长、叶宽和根长以及地上生物量显著增加。当GABA浓度为0.5 mmol·L-1处理时对干旱胁迫抑制燕麦幼苗株高、叶长、叶宽和根长以及地上生物的缓解效果最佳,结果表明添加适宜浓度的GABA有利于缓解逆境下植物幼苗生长抑制,提高植物的抗逆性。这是因为GABA是一种极易被吸收的氨基酸态氮源,有利于植物对硝酸盐的吸收,刺激根系生长,促进了植物中多胺合成,缓解环境胁迫带来的损伤[25]。
叶绿素是植物体内能进行光合作用的一类色素,植物的抗逆性强弱在很大程度上取决于其含量的高低。干旱胁迫下,植物叶绿体结构受到破坏,导致叶绿素含量下降和光合作用的降低[26]。在干旱胁迫下匍匐剪股颖(Agrostis stoloniferaL.)的叶绿素总量减低,相反,叶面喷施GABA后,叶绿素总量得到提高,改善了植物的光合效率[27]。同样,Iqbal B等[28]研究表明,干旱胁迫下甜椒(Capsicum annuumL.)幼苗叶绿素a、b和总叶绿素含量均显著降低,叶面喷施GABA后,其叶绿素含量均显著提高,且4 mmol·L-1效果最佳。本试验发现,随着干旱程度的增加,燕麦叶绿素a、叶绿素b、叶绿素总量含量以及叶绿素a/b均在降低,在喷施不同浓度的GABA后均显著提高,表明燕麦在经过干旱胁迫后喷施GABA促进植物自身调控机制,使叶绿素含量增加。这与徐溶蔓等[29-30]的研究结果一致。可见,经过外源GABA的处理,植物能够显著改善自身在干旱胁迫下的叶绿素含量,进而提高植物的光合效率。
叶绿素荧光参数作为植物光合作用机理和光合生理状况的关键指标,能够有效地反映植物在不同环境中光合反应的内在过程[31]。在植物的光合作用中,Fv'/Fm'被用作评估PSII光化学效能量子产量,是光能捕获效率的表达,在不同干旱程度下Fv'/Fm'值逐渐降低,表明干旱胁迫使叶绿素光系统Ⅱ反应中心内的光能转化效率下降,使光能利用能力下降[26]。ETR是植物体内光合作用电子传递速率快慢的重要指标,qP在一定程度上反映了PSⅡ中心的开放程度[32]。随着干旱程度的增加ETR呈下降趋势,表明干旱胁迫下植物体内PSⅡ中心电子传导受阻,导致光合功能下降,进而引起叶片光合速率降低[33]。本研究发现,随着干旱胁迫程度的增加,燕麦的Fo',Fm',Fv',Fv'/Fm',ETR以及qP大体上呈降低趋势,表明PSⅡ中心被破坏,光合电子的传递速率减慢,进而影响了光合电子的传递过程。据报道,随干旱程度的加剧,燕麦叶片Fm,Fv,Fv/Fm,Fv/Fo和qP显著降低,这与本试验结果一致[34]。随后在喷施GABA后,燕麦的Fo',Fm',Fv',Fv'/Fm', ETR以及qP均呈上升趋势,表明在干旱后喷施GABA可以缓解叶片光合系统的损伤,PSⅡ中心得到了修复。NPQ揭示了植物具有光保护的特性,可以将多余的光能转化为热能,对植物光合系统免受破坏提供保护作用[35]。本研究发现,燕麦的NPQ随着干旱程度的增加逐渐降低,在喷施GABA后总体呈上升趋势,说明燕麦在干旱胁迫后喷施GABA可以使PSⅡ反应中心的活性增强,提高了叶片中过剩光能的及时耗散,起到了保护光合结构的作用。
激发能的耗散方式主要包括PSⅡ天线热耗散百分率(Hd)、光化学反应百分率(Pc)、非光化学耗散百分率(Ex)。过剩的激发能是抑制植物光合的重要因素,随着胁迫程度的加剧,天线热耗散百分率逐渐升高,光化学反应百分率逐渐降低,非光化学耗散百分率所占百分率比例较[36]。本研究中,干旱处理下叶片Pc和Hd显著增加,Ex逐渐降低,在喷施GABA后,Pc,Hd和Ex总体呈上升趋势,这说明在干旱胁迫后喷施GABA使得植物吸收光能向光化学反应方向的转化率提高,这与光合、荧光参数的变化有关。而Ex的增加可防止PSⅡ和光合电子传递被还原,避免了光能过剩对光合传递结构的过度破坏,证明了喷施GABA可以提高燕麦抗旱的能力。该结论与沈小雪等[37-38]的研究结果一致。
4 结论
干旱胁迫下,喷施GABA提高了燕麦幼苗叶片中叶绿素含量,以及Fo',Fm',Fv',Fv'/Fm',qP,qN和ETR等荧光参数,有效改善了光能利用效率,使得幼苗的株高、叶长、叶宽、根长及地上单株生物量较对照增加。
参考文献
[1]"""""" 金祎婷,刘文辉,刘凯强,等. 全生育期干旱胁迫对‘青燕1号’燕麦叶绿素荧光参数的影响[J]. 草业学报,2022,31(6):112-126
[2]"""""" 冯蕊,周琪,吴令上,等. PEG6000模拟干旱胁迫对铁皮石斛幼苗生理和叶绿素荧光特性的影响[J]. 浙江农林大学学报,2024,41(1):132-144
[3]"""""" 戴海根,吴宏伟,吴尚卫,等. 干旱胁迫对鹰嘴紫云英种子萌发及幼苗叶绿素荧光特性的影响[J]. 草原与草坪,2022,42(5):95-105
[4]"""""" 韩同进,吴继华. 干旱胁迫对花生光合色素含量、叶绿素荧光参数及根系活力的影响[J]. 现代农业科技,2023(9):4-6
[5]"""""" 周笑,步园园,陈建桦,等. 外源褪黑素对干旱胁迫下越南槐幼苗生长及生理特性的影响[J]. 中药材,2023,46(10):2408-2413
[6]"""""" 韩喆,张永强,张浩浩,等. 干旱胁迫对伊犁绢蒿幼苗生长及叶片解剖结构的影响[J]. 草地学报,2024,32(1):105-112
[7]"""""" KOLUPAEV Y E,SHAKHOV I V,KOKOREV A I,et al. The role of reactive oxygen species and calcium ions in implementing the stress-protective effect of γ-aminobutyric acid on wheat seedlings under heat stress conditions[J]. Cytology and Genetics,2024,58(2):81-91
[8]"""""" LIGTHART K,FRYGANAS C,CONSTANTINOS P,et al. Physiology of γ-aminobutyric acid production by Akkermansia muciniphila[J]. Applied and Environmental Microbiology,2024,90(1):e01121-e01123
[9]"""""" 石彩玲,王沛潇,陈碧伟,等. γ-氨基丁酸对干旱胁迫下甘薯苗期渗透调节和抗氧化酶活性的影响[J]. 江苏师范大学学报(自然科学版),2023,41(3):30-35
[10]"""" 张碧茹,米俊珍,赵宝平,等. 外源γ-氨基丁酸缓解燕麦幼苗盐碱胁迫的生理效应[J]. 麦类作物学报,2024,44(2):222-229
[11]"""" 林欣琪,魏茜雅,梁腊梅,等. γ-氨基丁酸(GABA)种子引发缓解辣椒盐胁迫的效果及生理生化的变化[J]. 广西植物,2023,43(12):2338-2351
[12]"""" 陈彩锦,师尚礼,张尚沛,等. 饲用燕麦对干旱胁迫的响应及抗旱性评价[J]. 种子,2023,42(5):118-125
[13]"""" 赵小娜,高志昊,王斌,等. 15份饲用型燕麦种质材料苗期抗旱性比较与评价[J]. 草地学报,2023,31(12):3734-3743
[14]"""" ORTEGA-ORTEGA A,JIMÉNEZ-GALINDO J C,PARRA-QUEZADA R Á,et al. Osmotic stress tolerance in forage oat varieties(Avena Sativa L.)based on osmotic potential trials[J]. Agronomy Research, 2023,21(S1):335-346
[15]"""" 张国辉,陈彩锦,张尚沛,等. 不同饲用燕麦品种苗期生理抗旱性鉴定评价[J]. 饲料研究,2022,45(23):62-67
[16]"""" 王星宇,葛军勇,程静,等. 不同浓度PEG模拟干旱胁迫对燕麦种子胚根和胚芽生长的影响[J]. 陕西农业科学,2023,69(6):1-8,19
[17]"""" 李晓婷,李立军,张永平,等. 干旱胁迫下外源内生真菌对燕麦种子萌发及幼苗生长的影响[J]. 灌溉排水学报,2023,42(7):10-17,59
[18]"""" 李英浩,刘景辉,赵宝平,等. 干旱胁迫对燕麦生长及叶片光系统Ⅱ活性的影响[J]. 西北植物学报,2020,40(10):1706-1713
[19]"""" INSKEEP W P,BLOOM P R. Extinction coefficients of chlorophyll a and b in n,n-dimethylformamide and 80% acetone[J]. Plant Physiology,1985,77(2):483-485
[20]"""" 周艳虹,黄黎锋,喻景权. 持续低温弱光对黄瓜叶片气体交换、叶绿素荧光猝灭和吸收光能分配的影响[J]. 植物生理与分子生物学学报,2004,30(2):153-160
[21]"""" 李海霞,米银法,陈双臣. 外源钙对干旱胁迫下不同抗性牡丹PSⅡ功能和光能分配的影响[J]. 江苏农业科学,2022,50(16):120-127
[22]"""" 史建硕,潘丽佳,蒋龙刚,等. 外源γ-氨基丁酸对高温胁迫下番茄幼苗生长、抗氧化酶活性及光合特性的影响[J]. 华北农学报,2023,38(S1):188-193
[23]"""" 罗洁. 叶面喷施γ-氨基丁酸对黄瓜幼苗耐旱性的影响[J]. 农业技术与装备,2023(5):20-22,25
[24]"""" 于立尧,姚琪,李彭丽,等. 外源γ-氨基丁酸对干旱胁迫下甜瓜幼苗生长的影响[J]. 上海交通大学学报(农业科学版),2018,36(5):22-27
[25]"""" 李颖,鱼小军,李珍,等. 环境胁迫下γ-氨基丁酸对植物生长调控的研究进展[J]. 草地学报,2022,30(4):835-840
[26]"""" 南思睿,罗永忠,于思敏,等. 干旱胁迫后复水对新疆大叶苜蓿幼苗光合和叶绿素荧光的影响[J]. 草地学报,2022,30(5):1141-1149
[27]"""" LI Z,PENG Y,HUANG B R. Alteration of transcripts of stress-protective genes and transcriptional factors by γ-aminobutyric acid (GABA) associated with improved heat and drought tolerance in creeping bentgrass(Agrostis stolonifera)[J]. International Journal of Molecular Sciences,2018,19(6):1623
[28]"""" IQBAL B,HUSSAIN F,KHAN M S,et al. Physiology of gamma-aminobutyric acid treated capsicum annuum L. (sweet pepper) under induced drought stress[J]. Plos One,2023,18(8):e0289900
[29]"""" 徐溶蔓,侯典云,李春霞,等. 外源壳寡糖喷施对干旱胁迫下‘科大绿2号’幼苗生理特性及产量的影响[J]. 现代园艺,2023,46(23):1-6
[30]"""" 王晶,伏兵哲,李淑霞,等. 外源褪黑素对干旱胁迫下沙芦草幼苗生长和生理特性的影响[J]. 应用生态学报,2023,34(11):2947-2957
[31]"""" 刘军,齐广平,康燕霞,等. 不同灌溉处理下紫花苜蓿光合特性、叶绿素荧光参数及生物量的变化[J]. 草地学报,2019,27(6):1569-1576
[32]"""" 吴甘霖,段仁燕,王志高,等. 干旱和复水对草莓叶片叶绿素荧光特性的影响[J]. 生态学报,2010,30(14):3941-3946
[33]"""" 王方琳,柴成武,赵鹏,等. 3种荒漠 植物光合及叶绿素荧光对干旱胁迫的响应及抗旱性评价[J]. 西北植物学报,2021,41(10):1755-1765
[34]"""" 李英浩,刘景辉,赵宝平,等. 干旱胁迫下腐植酸对燕麦叶绿素荧光特性的调控效应[J]. 灌溉排水学报,2020,39(4):26-33
[35]"""" PAŁASZ A,PAŁKA M,FILIPCZYK Ł,et al. Effect of long-term treatment with classical neuroleptics on NPQ/spexin,kisspeptin and POMC mRNA expression in the male rat amygdala[J]. Journal of Neural Transmission(Vienna,Austria,2018,125(7):1099-1105
[36]"""" 王国莉,郭振飞. 低温对水稻不同耐冷品种幼苗光合速率和叶绿素荧光参数的影响[J]. 中国水稻科学,2005,19(4):381-383
[37]"""" 沈小雪,姜立得,陶佳伟,等. 退化秋茄林的补水修复实验研究[J]. 北京大学学报(自然科学版),2019,55(4):699-708
[38]"""" 铁得祥,胡红玲,喻秀艳,等. 桢楠幼树光合特性对镉胁迫的响应[J]. 生态学报,2020,40(11):3738-3746
(责任编辑" 刘婷婷)
猜你喜欢
现代畜牧科技(2021年6期)2021-07-16 05:49:54
中国粮油学报(2019年4期)2019-07-12 09:06:28
江苏农业科学(2016年8期)2017-02-15 19:40:24
江苏农业科学(2016年8期)2017-02-15 19:39:47
江苏农业科学(2016年8期)2017-02-15 19:38:57
安徽农学通报(2016年24期)2017-01-12 21:06:57
安徽农学通报(2016年21期)2016-12-22 16:15:06
山东农业科学(2016年11期)2016-12-17 21:16:16
中国科技博览(2016年14期)2016-04-25 02:04:42
现代园艺(2016年2期)2016-03-15 16:05:02
