
施用羊板粪对高寒矿区土壤微生物群落结构及其功能的影响
2025-03-06 00:00:00于中阳胡生斌安晓婷杨明春姚喜喜苏晓燕李长慧
草地学报 2025年2期
关键词:土壤理化性质




摘要:为探究施用羊板粪对高寒矿区土壤理化性质、微生物群落结构及其功能的影响,本研究设置了不同羊板粪施用量(0 m3·m-2,CK;0.04 m3·m-2,T1;0.05 m3·m-2,T2;0.06 m3·m-2,T3),对施用后第二年的土壤相关参数进行了测定和分析,发现施用羊板粪显著提高了土壤全氮、全磷、土壤有机质、速效氮和速效磷含量,并显著改变了细菌和真菌群落的Beta多样性、部分优势微生物菌群及功能微生物的相对丰度。冗余分析发现,土壤有机质、全氮和全磷是影响高寒矿区土壤细菌和真菌群落分布的关键土壤理化因子。综合而言,施用羊板粪对高寒矿区的土壤质量和微生物环境具有促进作用,建议木里矿区土壤修复可将羊板粪用量设置为0.06 m3·m-2。
关键词:高寒矿区;羊板粪;土壤理化性质;微生物群落结构;冗余分析
中图分类号:S154.36""""""" 文献标识码:""""""" 文章编号:1007-0435(2025)02-0654-11
Effects of Applying Sheep Slat Manure on the Structure of Soil Microbial Community and Its Function in Alpine Mining Area
YU Zhong-yang1,2, HU Sheng-bin1, AN Xiao-ting1,2, YANG Ming-chun1, YAO Xi-xi1, SU Xiao-yan1,2,
LI Chang-hui1*
(1.College of Agriculture and Animal Husbandry, Qinghai University, Xining, Qinghai Province 810016, China;
2.Veterinary Medicine and Academy of Animal Science, Qinghai University, Xining, Qinghai Province 810016, China)
Abstract:In order to investigate the effects of sheep slat manure application on soil physicochemical properties, microbial community structure and its function in alpine mining area, this study set different sheep slat manure application rates (0 m3·m-2, CK; 0.04 m3·m-2, T1; 0.05 m3·m-2, T2; 0.06 m3·m-2, T3), and measured and analyzed the soil-related parameters in the second year after application. The results showed that the application of sheep slat manure significantly increased soil total nitrogen, total phosphorus, soil organic matter, quick-acting nitrogen and quick-acting phosphorus contents, and significantly changed the Beta diversity of bacterial and fungal communities, the relative abundance of some dominant microflora and functional microorganisms. Redundancy analysis revealed that soil organic matter, total nitrogen and total phosphorus were the key soil physicochemical factors affecting the distribution of soil bacterial and fungal communities in alpine mining areas. In summary, the application of sheep slat manure had obvious promotion effects on soil quality and microbial environment in alpine mining areas, and it suggested that the dosage of sheep slat manure should be set to 0.06 m3·m-2 for soil remediation in the Muli mining area.
Key words:Alpine mining area;Sheep slat manure;Soil physicochemical properties;Microbial community structure;Redundancy analysis
木里矿区位于青藏高原东北部,是我国重要的煤炭开采基地,同时也是青藏高原典型的生态脆弱区[1-2]。经过多年的煤炭开采,对周边地区的生态环境造成了严重的污染,尤其是土壤污染和水土流失[3]。青海木里地处多年冻土区和草甸草原区,生态系统极其脆弱,退化趋势难以逆转[4]。矿区因煤炭开采形成了大量的泥砂岩,这种矿渣雨淋和风蚀的在作用下进一步形成了大量的渣土,然而,这种渣土相比于一般的土壤又存在很大的差异,其土壤基质很少,养分匮缺,难以生长植物[5]。因此,在这种渣土之上建植人工草地面临巨大挑战。近年来,经过科研工作者的不懈努力,矿区生态环境得到了部分恢复[5-7]。采用客土法能够有效的恢复退化土壤,改善土壤质量,从而建立高效的栽培草地[6]。然而,客土法在高寒矿区退化草地治理过程中依旧面临着诸多问题,例如,优质土壤供需量大,运输路线长,成本高昂等问题[6,8]。因此,采用客土法来恢复高寒矿区的土壤质量并非最佳的选择,亟需探索更加科学的途径来恢复木里矿区的土壤质量。
木里矿区周围分布着大量的牧民,长年以牧羊为主。矿区周边地表分布着大量的羊粪,在畜蹄踩踏和雨水的作用下,羊粪和土壤充分混合,形成了大量的羊板粪[6-7]。这些羊板粪通常被周边的农牧民捡拾用作生活燃料[9]。近年来,随着城镇化水平的不断提高,农牧民生活质量普遍提升,很少再捡拾羊板粪充当燃料,导致这一资源的浪费[9]。事实上,羊板粪除了充当燃料之外,还具有改善土壤质量,提高土壤养分等关键作用[10-11]。羊板粪中含有丰富的有机物质和氮、磷、钾等植物生长所需的营养元素,可以增加土壤的有机质含量,能够更好地满足作物生长的需要。研究发现,长期施用羊板粪可以改善土壤的物理性质,如通气性和保水性,有助于作物根系的生长[5,12]。此外,施用羊板粪还可以培育土壤微生物、促进微生物代谢、改善微生物群落结构和提高土壤的生物活性。化肥减量配施有机肥对土壤养分、物理特性、土壤酶活性及微生物群结构有显著改善效果[13]。如果将羊板粪和矿区的渣土结合起来,将会对矿区土壤的修复和人工草地的建植具有一定的积极作用。羊板粪代替了部分有机肥,极大地节约了经济成本,而且运输路线短,是一种经济有效的矿区植被重建途径。
在生产实践中,农牧民将羊粪施入农田以维持作物产量,或用于提升蔬菜瓜果的产量[14-15]。在此过程中,农牧民通常根据作物产量最高的标准来判断最佳的羊粪施用量。然而,羊粪对土壤的影响仍不清楚。肥料的施入极大地改善了土壤理化性质和微生物群落结构[16-18]。土壤氮含量是限制作物产量的关键土壤理化因子,一定阈值内,土壤全氮含量和作物产量呈显著正相关[19-20]。此外,土壤微生物在调节作物产量方面也发挥着重要的作用[21]。某些调节作物产量的微生物数量会在施肥状态下表现出明显的动态变化,这种变化因不同肥料类型、施肥量和区域环境而各有差异[22]。施用氮肥虽然降低了土壤微生物多样性,但极大地提高了土壤养分含量,促进了微生物功能基因的相对丰富,从而创造了适宜植物生长的土壤环境[23-25]。然而,关于羊板粪施用量对土壤理化性质和微生物群落结构影响的研究目前还未见报道,进一步探究其影响可对作物生产、高寒矿区的土壤修复与植被重建起到至关重要的作用。
本研究采用高寒矿区附近的羊板粪为施肥来源,设置了不同施肥梯度,以普遍用于青藏高原退化草地恢复的‘青海’草地早熟禾(Poa pratensis‘Qinghai’)、‘青海’冷地早熟禾(Poa crymophila‘Qinghai’)、‘青海’中华羊茅(Festuca sinensis‘Qinghai’)和‘同德’短芒披碱草(Elymus sibiricus‘Tongde’)为草种为供试材料,进行栽培草地的建植,本研究在建植后的第二年对土壤理化性质和微生物群落结构进行测定,旨在揭示施用羊板粪对高寒矿区土壤理化性质和微生物群落结构的影响,为高寒矿区的土壤修复提供科学依据和理论指导。
1 材料与方法
1.1 研究区概况
本试验区位于青藏高原东北部的青海省海西蒙古族藏族自治州天峻县的木里煤田聚乎更矿区(100°55'E,36°17'N),平均海拔约3800~4200 m,以高原冰缘地貌类型为主。矿区天然植被类型分为高寒沼泽类和高寒草甸类,具有高寒地区形态特征,植物群落结构简单,植被稀疏,对人类活动的抗干扰力较弱。聚乎更矿区为典型的高原大陆性气候,该区域气候寒冷,昼夜温差极大,6—8月份为雨季,11月至翌年5月以降雪为主,年平均气温为-4.2℃,年平均降水量为477.1 mm左右,年平均蒸发量为1049.9 mm,长冬无夏,属于祁连山高寒山地多年冻土区,区内冻土广泛发育,多年冻土厚度40~160 m,平均厚度120 m,多年冻土层深度上限0.95~5.50 m。矿区渣山植被恢复前表土基况为全氮含量1.17 g·kg-1,全磷含量0.91 g·kg-1,速效氮含量18.00 mg·kg-1,速效磷含量6.70 mg·kg-1,有机质含量93.33 g·kg-1,pH值为8.46。
1.2 试验设计
于2022年6月29日,选择储煤场的平坦区域作为试验地块进行小区布设。试验共设置4个羊板粪(N 2.1%,P2O5 0.4%,有机质15.8%)处理(不添加羊板粪,CK;0.04 m3·m-2,T1;0.05 m3·m-2,T2;0.06 m3·m-2,T3)。将羊板粪撒于事先设置好的小区内,用旋耕机进行翻地,使羊板粪和渣土混合均匀,耕层深度约30 cm。采用随机区组试验设计,每处理重复3次,共12个小区,小区面积为6 m×5 m。本研究选取青藏高原的适生乡土草种:‘青海’草地早熟禾(Poa pratensis‘Qinghai’)、‘青海’冷地早熟禾(Poa crymophila‘Qinghai’)、‘青海’中华羊茅(Festuca sinensis‘Qinghai’)和‘同德’短芒披碱草(Elymus sibiricus‘Tongde’),总播量为22.5 g·m-2,比例(重量比)为1∶1∶1,草种均来自青海大学畜牧兽医科学院。将同等量的牧草专用肥(总养分≥35%,N 18%,P2O5 12%,K2O 5%)与种子充分混合后均匀撒播,轻耙耱平,使种子埋入表层土壤中。采用人工方式进行镇压,并铺设无纺布以保温促芽。
1.3.1 样品采集方法 2023年7月30日,利用五点取样法,用直径3.5 cm的土钻采集各小区0~10 cm土壤样品,将5个钻点的土壤样品混合均匀后分为2份,其中1份过1 mm筛用于测定土壤养分含量,将另1份土样装满50 mL灭菌离心管并保存在-80℃进行微生物高通量测序。
1.3.2 土壤理化性质测定 参照鲍士旦[26]的土壤农化分析,进行土壤理化性质的测定。使用pH计(德国Sartorius PB-10)测定土壤pH(水土比为1∶2.5)[27]。采用重铬酸钾(K2Cr2O7)氧化法测定土壤有机质含量(SOM)[28]。用H2SO4和H2O2湿消解后用比色法测定总磷(TP)(UV2800A紫外分光光度计,UNIC Inc.,China)[29]。速效氮(AN)采用碱解扩散法测定[30]。全氮(TN)含量采用元素分析仪测定(FLASH SMART CHNS/0,Germany)。速效磷(AP)采用钼酸蓝法进行测定[31]。
1.3.3 土壤微生物群落的测定 委托上海美吉生物科技有限公司,通过1.0%琼脂糖凝胶电泳和NanoDrop2000分光光度计(Thermo Scientific,美国)测定DNA的质量和浓度,并在进一步使用前保持在-80℃。用引物对338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3'分别对土壤细菌和真菌进行基因序列测定。从2%琼脂糖凝胶中提取PCR产物,并根据制造商的说明使用PCR纯化试剂盒(YuHua,Shanghai,China)纯化,并使用Qubit 4.0(Thermo Fisher Scientific,USA)进行定量。FASTQ文件使用内部perl脚本进行多路复用解复用,然后由fastp版本0.19.6进行质量过滤,并由FLASH版本1.2.7合并。然后,使用UPARSE 7.1将优化的序列聚类为可操作的分类单元(OTU),序列相似度为97%。选择每个OTU最丰度的序列作为代表性序列。为了尽量减少测序深度对α和β多样性测量的影响,将每个样本的16S rRNA基因序列数量稀疏到20 000个,平均Good覆盖率分别为99.09%。纯化的扩增子以等摩尔量合并,并根据Majorbio Bio-Pharm Technology Co.,Ltd.(中国上海)的标准方案在Illumina PE300/PE250平台(Illumina,San Diego,USA)上进行双端测序。
1.3 数据分析与作图
采用SPSS 27.0对不同处理下的土壤理化性质和微生物结构及多样性进行单因素方差分析(ANOVA)。使用基于bray-curtis距离算法的PCoA分析(主坐标分析)检验样本间细菌和真菌群落结构的相似性,并结合PERMANOVA非参数检验分析样本组间微生物群落结构差异是否显著;用LEfSe分析(linear discriminant analysis Effect Size)确定不同组间门和属水平丰度显著差异的细菌和真菌类群。使用在线分析平台(https://cloud.majorbio.com/page/project/overview.html)对不同处理的土壤细菌功能(FAPROTAX)和真菌功能(FUNGuild)做了预测分析。采用冗余分析(redundancy analysis,RDA)来对探究土壤理化指标对土壤细菌和真菌群落结构的影响。使用在线网站(https://www.omicstudio.cn/tool/140)制作基于Pearson相关系数的地上生物量与土壤理化性质和微生物多样性关系的热图。文章中的其他图均在Origin 2022软件中完成。
2 结果与分析
2.1 羊板粪添加对土壤理化性质的影响
与CK处理相比较,不同羊板粪添加处理下的土壤pH差异不显著(图1A)。随着羊板粪用量的增加,土壤全磷(TP)、土壤有机质(SOM)、速效氮(AN)和速效磷(AP)含量显著提高(Plt;0.05),且均在T3水平时达到峰值(图1C-F)。此外,施羊板粪显著提高了土壤全氮(TN)含量(Plt;0.05),T1处理和T2处理差异不显著,T3处理达到峰值,T1、T2和T3处理分别较CK处理提高了117.26%,126.06%和211.07%(图1B)。
2.2 羊板粪添加对土壤微生物群落的影响
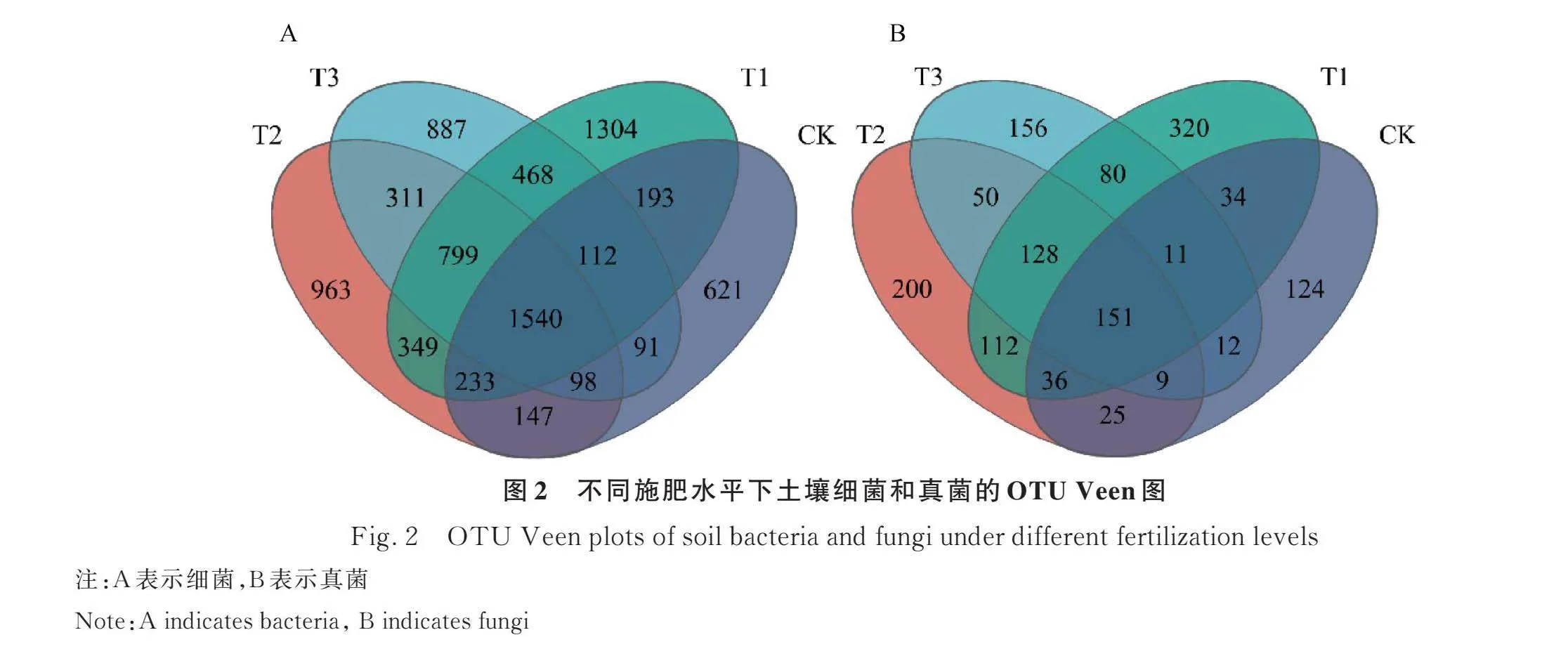
2.2.1 OTU数量分析 基于OTU水平,对不同处理下土壤细菌和真菌群落进行了韦恩分析,并绘制了韦恩图(图2)。由细菌群落的韦恩分析可以看出,CK、T1、T2和T3处理分别识别出3035,4998,4440和4306个OTU(图2A),不同处理共有的细菌OTU总数有1540个。与CK处理相比较,施用羊板粪增加了土壤中特有细菌的OTU数量,4种处理下土壤中细菌OTU分布数目由大到小为:T1gt;T2gt;T3gt;CK。由真菌群落的韦恩分析可以看出,CK、T1、T2和T3处理分别识别出402,872,711和597个OTU(图2B),不同处理共有的真菌OTU数目为151个,CK、T1、T2和T3处理所特有的OTU数目分别为124,320,200和156个,占比分别为8.56%,22.10%,13.81%和10.77%。与CK处理相比较,施用羊板粪增加了土壤中特有真菌的OTU数量,4种处理下土壤中真菌OTU分布数目由大到小为:T1gt;T2gt;T3gt;CK。
2.2.2 土壤微生物群落多样性 对不同羊板粪用量下的土壤细菌和真菌群落的Alpha多样性进行了分析(表1)。从表1可以看出,不同羊板粪施用量对土壤细菌和真菌群落的Alpha多样性具有不同程度的影响。总体来看,添加羊板粪提高了细菌和真菌群落的OTU指数、Chao 1指数和Shannon指数,降低了细菌和真菌群落的Simpson指数。相较于CK处理,T1和T2处理显著提高了细菌群落的OTU指数、Chao 1指数和Shannon指数,以及真菌群落的OTU指数和Chao 1指数(Plt;0.05)。施用羊板粪对土壤细菌和真菌群落的Simpson指数无显著影响。
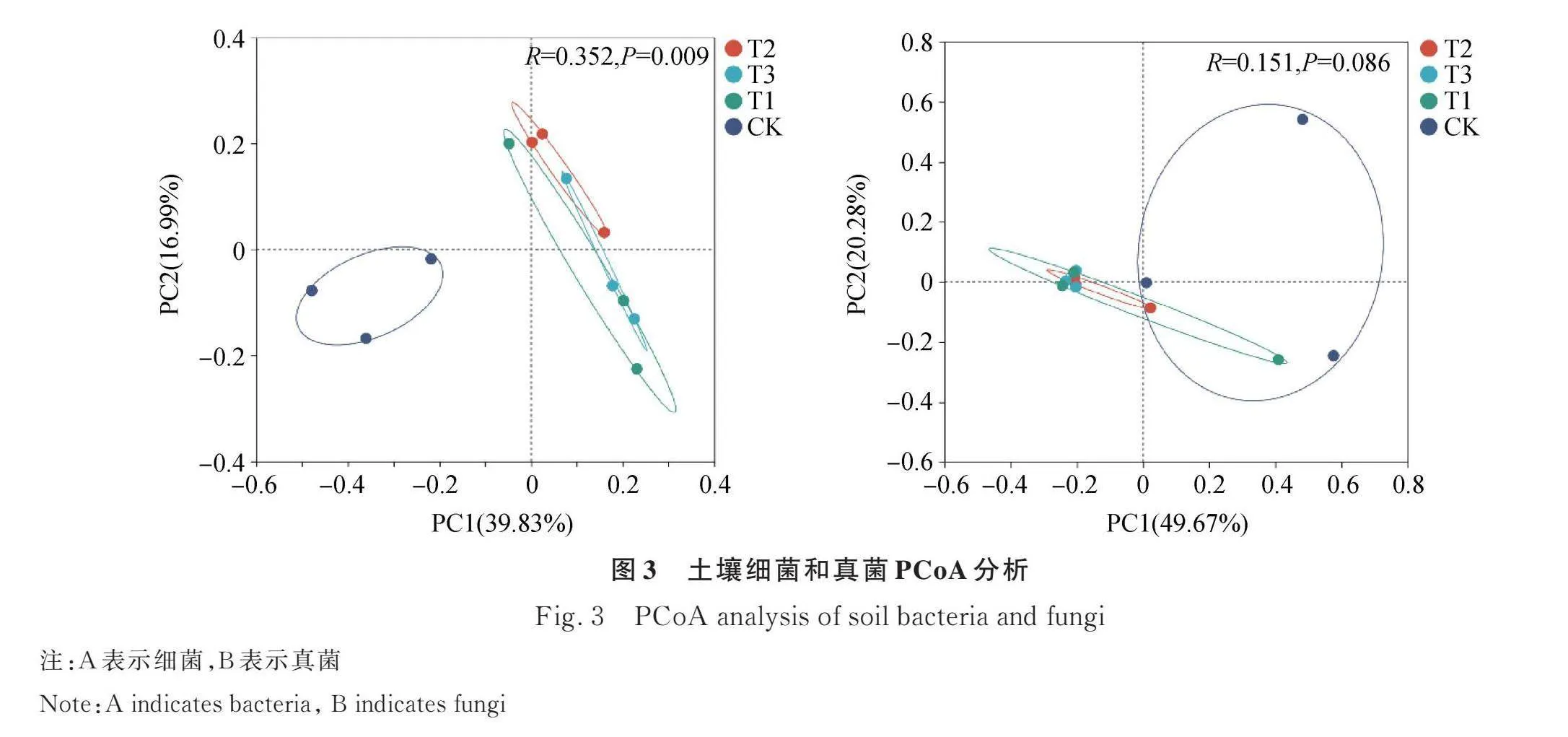
基于Bray-Curtis距离,对不同处理下的土壤细菌和真菌群落做了PCoA分析(图3)。在细菌群落的PCoA分析中,两个主成分的方差累积解释率为56.82%,组间差异显著(R=0.352,P=0.009,图3A),T2和T3处理样本聚集在一起,且与T1处理有一定的重叠,3个羊板粪处理下的细菌群落与对照组发生分离。在真菌群落的PCoA分析中,两个主成分的方差累积解释率为69.95%,组间差异不显著(R=0.151,P=0.086,图3B),4个处理下的真菌群落聚集在一起,不同群落之间重叠度较高。
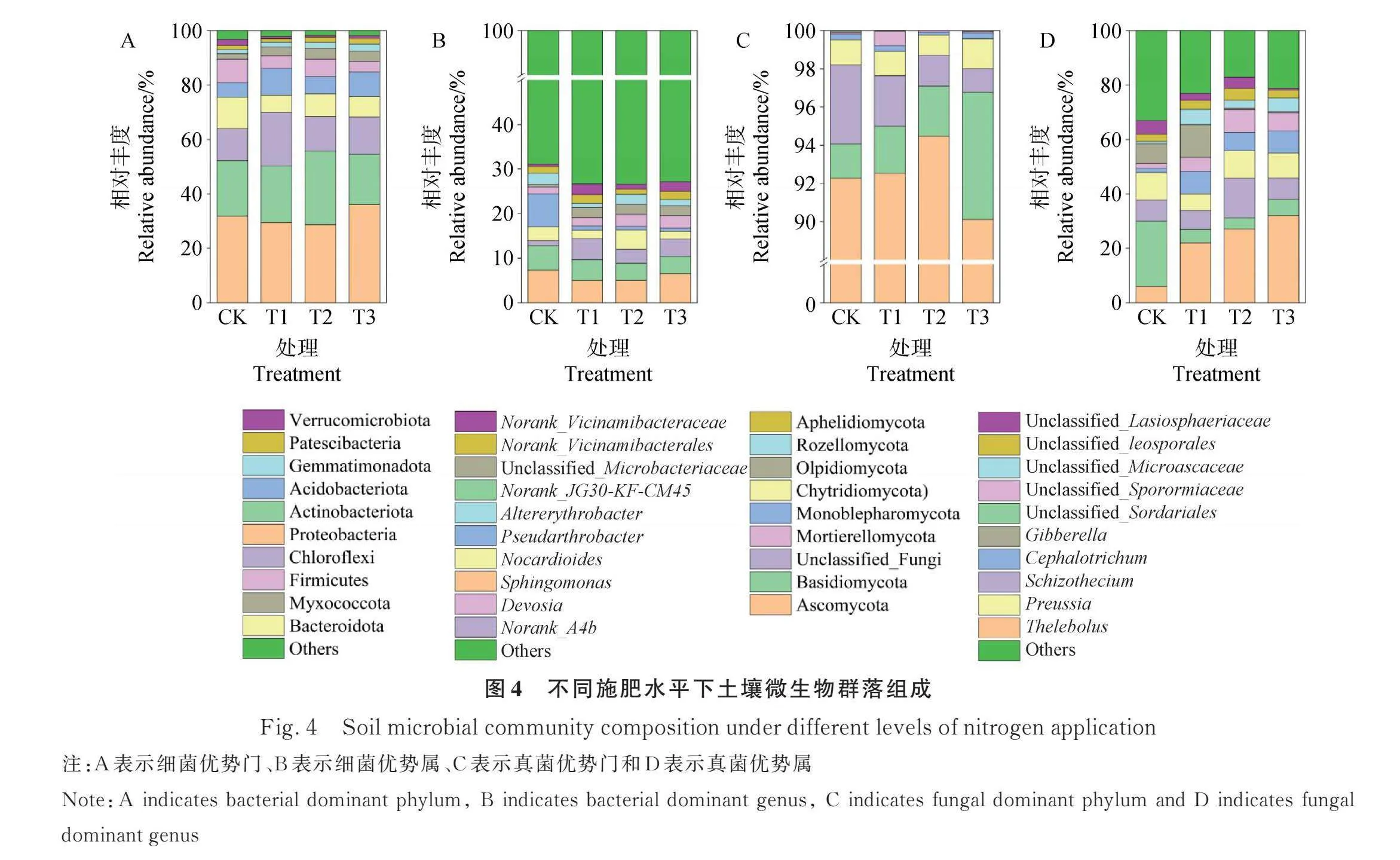
2.2.3 土壤微生物群落组成 所有的土壤样本中观察到Proteobacteria(变形菌门)、Actinobacteriota(放线菌门)、Chloroflexi(绿弯菌门)、Bacteroidota(拟杆菌门)和Acidobacteriota(酸杆菌门)等细菌门(图4A)和Ascomycota(子囊菌门)、Basidiomycota(担子菌门)、Chytridiomycota(壶菌门)、Monoblepharomycota(单毛壶菌门)和Mortierellomycota(被孢霉门)等真菌门(图4C)。此外,观察到Sphingomonas(鞘氨醇单胞菌属)、Norank_JG30-KF-CM45、Norank_A4b、Nocardioides(类诺卡氏菌属)和Pseudarthrobacter(假节杆菌属)等细菌属(图4B)和Thelebolus(寡囊盘菌属)、Schizothecium(裂壳菌属)、Preussia(光黑壳属)和Gibberella(赤霉菌属)等真菌属(图4D)。对于细菌门而言,与CK处理相比较,施用羊板粪提高了绿弯菌门、酸杆菌门、Myxococcota(粘菌门)和Gemmatimonadota(芽单胞菌属)等细菌门的相对丰度,降低了拟杆菌门、Firmicutes(厚壁菌门)和Verrucomicrobiota(疣微菌门)的相对丰度,且随着羊板粪用量的增加,芽单胞菌属的相对丰度逐渐增加(图4A)。对于真菌门而言,与CK处理相比较,施用羊板粪提高了拟杆菌门的相对丰度,降低了壶菌门的相对丰度(图4C)。对于细菌属而言,与CK处理相比较,施用羊板粪提高了Norank_A4b、Devosia(德沃斯氏菌属)和Norank_Vicinamibacteraceae等细菌属的相对丰度,降低了鞘氨醇单胞菌属、Norank_JG30-KF-CM45和假节杆菌属等的细菌属的相对丰度,且随着羊板粪用量的增加,Norank_JG30-KF-CM45和假节杆菌属的相对丰度逐渐降低,德沃斯氏菌属的相对丰度逐渐增加(图4B)。对于真菌属而言,与CK处理相比较,施用羊板粪提高了寡囊盘菌属和Cephalotrichum的相对丰度,降低了光黑壳属的相对丰度,且随着羊板粪用量的增加寡囊盘菌属的相对丰度逐渐增加(图4D)。
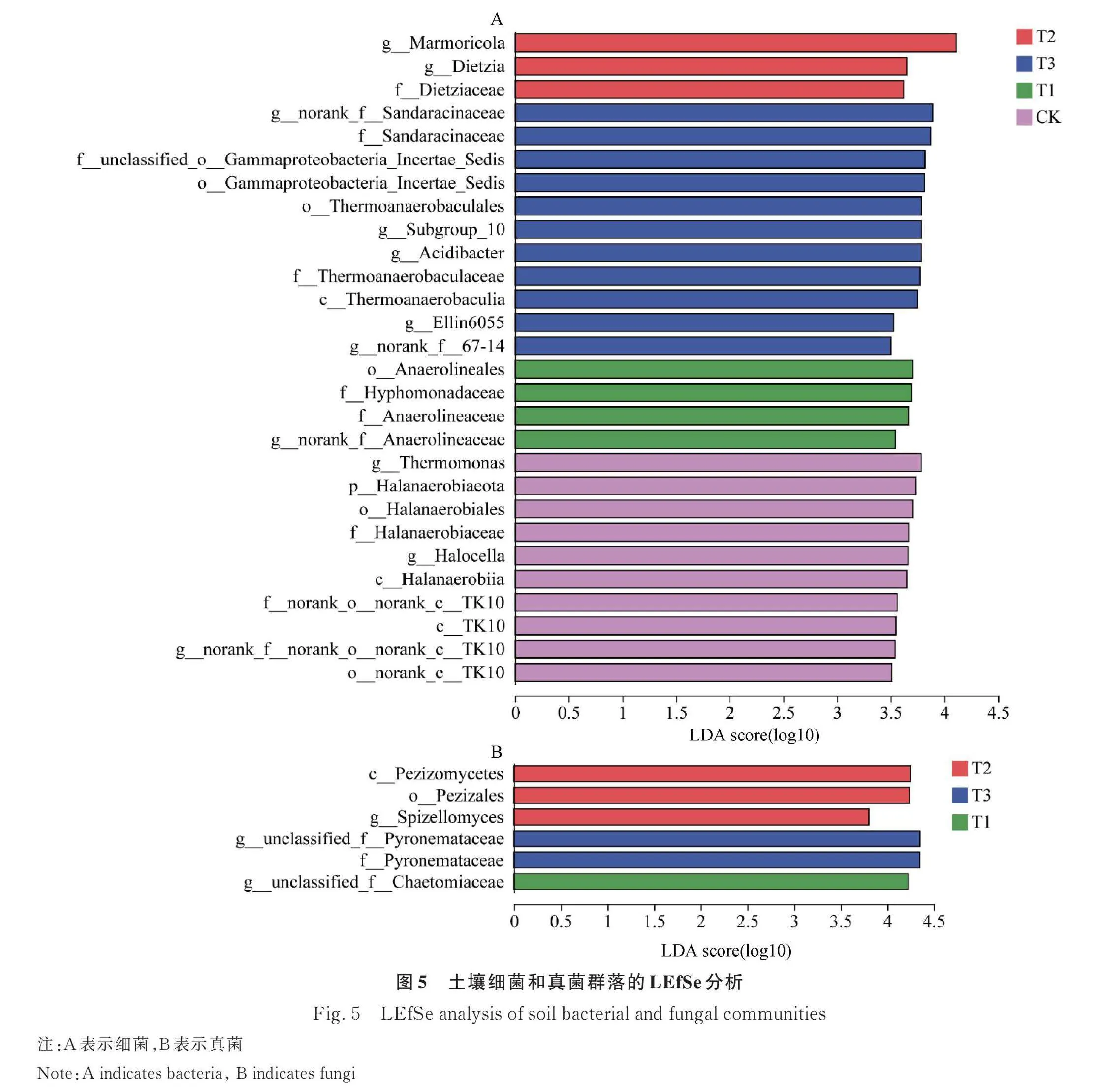
利用LEfSe分析对不同处理下的差异细菌和真菌进行了鉴定(图5)。在细菌群落中,LDA分析检测到28个(CK=10, T1=4,T2=3,T3=11)具有显著生物统计学意义的差异微生物(图5A)。在分类水平上,CK、T1、T2和T3处理得分最高的生物标记物分别为g_Thermomonas、o_Anaerolineales、g_Marmoricola和g_Norank_Sandaracinaceae(图5A)。
在真菌群落中,LDA分析检测到6个(CK=0,T1=1,T2=3,T3=2)具有显著生物统计学意义的微生物标志物(图5B)。数据过筛时将LDA值设置为2以上,均未显示其差异标记微生物。在分类水平上,T1、T2和T3处理得分最高的生物标记物分别为g_unclassified_Chaetomiaceae、c_Pezizomycetes和g_unclassified_Pyronemataceae。
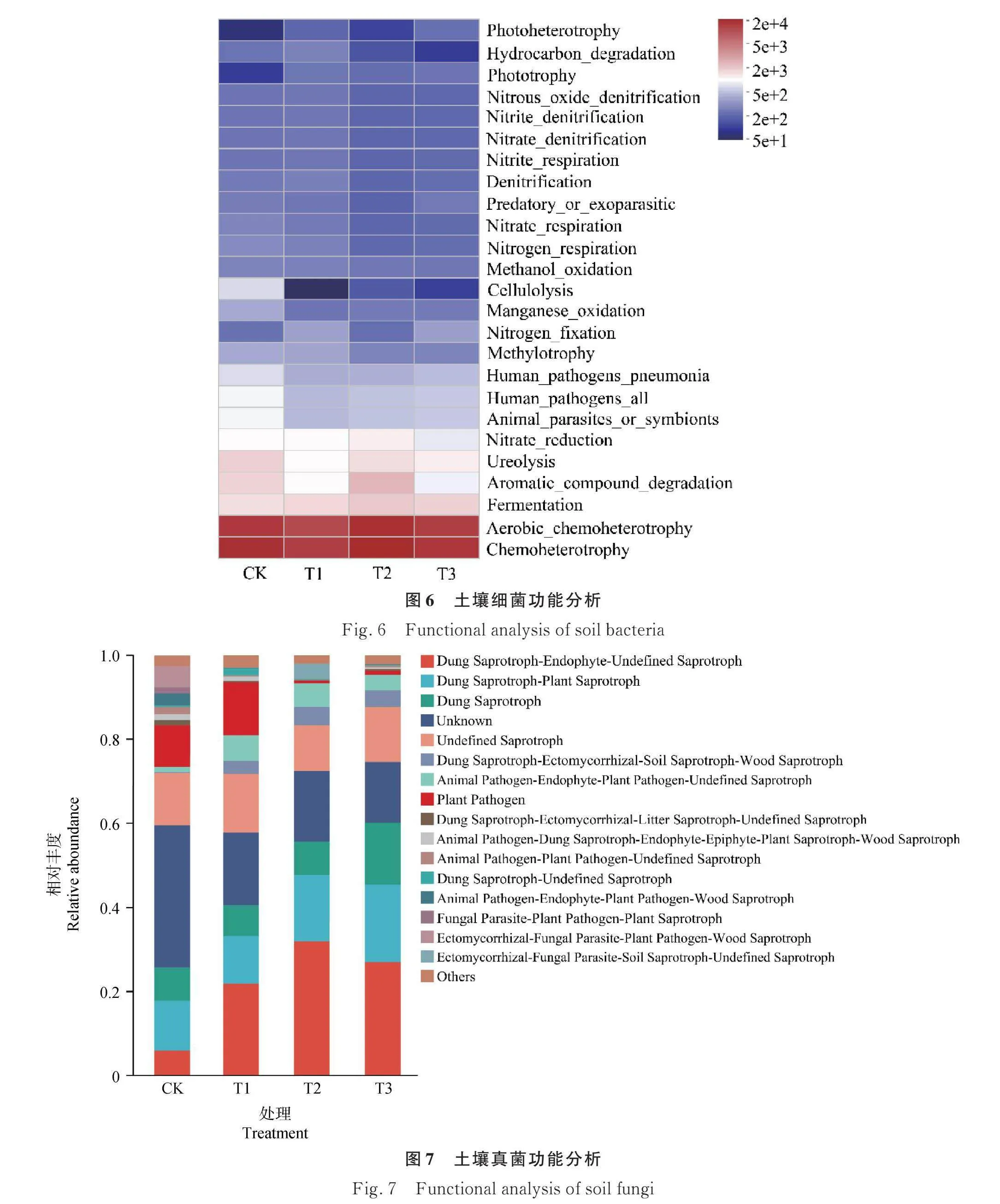
2.2.4 土壤微生物功能分析 施用羊板粪改变了土壤细菌和真菌功能微生物的相对丰度。土壤细菌的主要功能类群为Chemoheterotrophy(化能异养)、Aerobic_chemoheterotrophy(好氧化能异养)和Fermentation(发酵)(图6)。土壤真菌的主要功能类群为Dung Saprotroph-Endophyte-Undefined Saprotroph(粪腐生菌-内生菌-未定义腐生真菌)、Dung Saprotroph-Plant Saprotroph(粪腐生菌-植物腐生菌)和Dung Saprotroph(粪腐生菌)(图7)。施用羊板粪增加了Fermentation(发酵)、Cellulolysis(纤维溶解)、Manganese_oxidation(锰氧化)的相对丰度,降低了Phototrophy(光养)的相对丰度。随着羊板粪施用量的增加,粪腐生菌-植物腐生菌的相对丰度有增加趋势。不同处理相比较,T3水平显著增强了Hydrocarbon_degradation、未定义腐生真菌、粪腐生菌和粪腐生菌-植物腐生菌的相对丰度,T2水平显著增强了植物腐生菌的相对丰度。
2.4 环境因子与微生物群落的冗余分析
采用冗余分析对土壤理化因子和土壤细菌、真菌群落之间的相互关系做了进一步分析(图8),土壤理化性质与细菌优势门的冗余分析中,两个排序轴共解释了37.53%的群落变异(图8A)。TP(r2=0.7255,P=0.003)、TN(r2=0.7081,P=0.005)、SOM(r2=0.8176,P=0.007)、AN(r2=0.5505,P=0.023)和AP(r2=0.5049,P=0.041)是影响优势细菌门分布的关键土壤理化因子。土壤理化性质与细菌优势属的冗余分析中,两个排序轴共解释了33.093%的群落变异(图8B)。SOM(r2=0.6214,P=0.015)、TN(r2=0.5114,P=0.036)、TP(r2=0.4839,P=0.047)是影响优势细菌属分布的关键土壤理化因子。土壤理化性质与真菌优势门的冗余分析中,两个排序轴共解释了39.57%的群落变异(图8C)。TN(r2=0.4134,P=0.065)是影响优势真菌门分布的关键土壤理化因子。土壤理化性质与真菌优势属的冗余分析中,两个排序轴共解释了34.19%的群落变异(图8D)。SOM(r2=0.5741,P=0.022)、TN(r2=0.5404,P=0.024)、TP(r2=0.5369,P=0.026)是影响优势真菌属分布的关键土壤理化因子。综上所述,SOM、TN和TP是影响细菌和真菌群落分布的关键土壤理化因子。
3 讨论
高寒矿区的土壤属于矿渣土,土壤基质较少,养分含量极低,导致植物难以在此环境中生存繁殖[32],解决这一问题的关键在于增加土壤基质和提高土壤养分[33]。羊板粪作为一种有机肥料,不仅能够提供作物所需的养分,还能促进土壤微生物的生长和代谢,从而改善土壤环境和提高土壤质量[7]。因此,施用羊板粪便成为最有效的方法。羊板粪中含有大量的土壤基质和碳、氮、磷等营养元素[34],施入土壤后,不仅能改变土壤的物理性质,而且还能有效增强土壤的化学性质,对高寒矿区土壤的修复具有重要作用[6]。本研究通过设置不同用量的羊板粪施用量,发现施用羊板粪对土壤pH的影响较小,这可能是因为羊板粪是一种碱性肥料,原本会导致土壤pH值增加[35],但施用羊板粪后增加了土壤微生物的活性,促进了有机质的分解,促使磷酸盐溶解,将氢离子释放出来,从而降低pH值,二者相互抵消导致施用羊板粪对土壤pH的影响较小。随着羊板粪施用量的增加,土壤中的TN,TP,AN,AP和SOM含量显著提高。这主要是因为高寒矿区的土壤养分限制非常严重,对肥料的响应及其敏感[36]。在一定阈值内,土壤养分含量会随着施肥量的增加出现一个峰值而后下降,但本研究中土壤养分含量随着羊板粪施用水平的提高而逐渐增加,说明本研究的施肥量过低,在后期的研究中应加大施肥量进一步来确定最佳的羊板粪施用量。
不同羊板粪施用量均提高了土壤和真菌群落的OTU数量,这是因为羊板粪中含有较多的微生物,将其施入土壤后,微生物以土壤为载体进一步繁殖扩增,导致其数量增加[37]。本研究发现,尽管施加羊板粪可以提高土壤细菌和真菌群落的OTUs指数和Chao 1指数,但总体表现为低施肥量大于高施肥量,这可能是因为在低施肥量条件下,微生物之间的竞争压力可能较低,有利于共生关系的建立,从而促进微生物活性[38]。高施肥量可能导致土壤中某些化学物质(如硝酸盐、氨等)的积累,这些物质可能对微生物有毒害作用,从而抑制微生物活性[39]。通过PCoA分析发现,不同羊板粪施用量下的细菌群落发生了明显的分离,而真菌群落整体上聚集在一起,说明施用羊粪对土壤细菌群落的影响明显大于真菌群落,该结论和温烜琳等[40]研究结论相似,主要的原因可能是细菌群落对外界环境的响应较敏感,细菌群落更易受到养分供给的影响[41]。本研究中,施用羊板粪降低了拟杆菌门和假节杆菌属的相对丰度,提高了酸杆菌门、Norank_A4b和担子菌门的相对丰度。此外,不同施用量对微生物相对丰度具有明显的影响[42]。例如,随着施肥量的增加,担子菌门和寡囊盘菌属的相对丰度逐渐提高,Unclassified_fungi的相对丰度逐渐降低。这些现象充分不同羊板粪施用量对土壤微生物群落具有明明显的调控作用。担子菌门对于作物的栽培非常有益,其菌根不仅可以帮助植物吸收养分,还能促进土壤中有机物质的分解[43];寡囊盘菌属具有木质素降解功能,加快腐殖质的分解,为土壤提供养分[44]。养分添加会显著改变土壤微生物的生态功能[45],本研究也得到了相似的结论。施用羊板粪增加了发酵、纤维溶解、锰氧化和粪腐生菌的相对丰度,这些生态功能都有利于促进土壤养分周转,从而提高高寒矿区土壤的生态系统多功能性。因此,合理施用羊板粪有利于改善高寒矿区的土壤质量和微生物群落结构及其功能。
土壤微生物与土壤理化性质相互作用,形成一个复杂的生态系统,这种关系对于土壤肥力的维持具有意义[46]。土壤微生物可以分解土壤中的有机物质,将其转化为无机物质,从而影响土壤的有机质含量和养分状况;它还可以通过分解纤维素、木质素等复杂有机物,影响土壤结构,增加土壤孔隙度,改善土壤通气性和渗透性[47]。土壤理化性质如pH值和含水量等,都会直接影响微生物的生长和代谢活动[48]。本研究通过冗余分析探索了土壤理化性质对微生物群落结构的影响,结果表明,SOM、TN和TP是影响细菌和真菌群落分布的关键土壤理化因子,这和以往大多研究结果相似[49-50]。本研究中,T3处理显著促进了SOM,TN和TP的含量,说明T3处理通过提高土壤养分含量明显改善了土壤微生物群落结构,可作为高寒矿区土壤修复的最佳羊板粪施用量。
4 结论
随着羊板粪施用量的增加,土壤全氮、全磷、土壤有机质、速效氮和速效磷含量均逐渐提高,在T3水平时达到峰值。不同羊板粪施用量对土壤微生物群落的Beta多样性、部分优势微生物菌群和功能微生物具有显著影响。土壤有机质、全氮和全磷是影响高寒矿区土壤细菌和真菌群落分布的关键土壤理化因子。因此,施用0.06 m3·m-2的羊板粪有助于改善高寒矿区的土壤理化性质和微生物群落结构,对高寒矿区的土壤质量具有显著的促进作用。
参考文献
[1]"""""" 李聪聪,王佟,王辉,等. 木里煤田聚乎更矿区生态环境修复监测技术与方法[J]. 煤炭学报,2021,46(5):1451-1462
[2]"""""" 许泰,鄂崇毅,郑超. 高寒露天矿区渣土基质粒度组成及养分特征[J]. 生态科学,2024,43(2):132-140
[3]"""""" 金立群,李希来,孙华方,等. 高寒矿区排土场不同坡向植被和土壤特征研究[J]. 土壤,2020,52(4):831-839
[4]"""""" 王锐,李希来,马钰,等. 青海圣雄煤矿煤矸石堆积对周边高寒湿地植被和土壤的影响[J]. 土壤,2020,52(2):386-393
[5]"""""" 刘万杰,蒋福祯,马利利,等. 有机肥施用量和播种方式对高寒矿区植物群落生长和土壤养分的影响[J]. 草原与草坪,2023,43(2):116-125
[6]"""""" 张玉芳,李希来,金立群,等. 基于高寒矿区颗粒有机肥和羊板粪配施的无客土重构土壤理化性质分析[J]. 中国土壤与肥料,2023(7):129-137
[7]"""""" 乔千洛,武燕茹,李钦瑶,等. 商品有机肥和羊板粪对高寒矿区人工草地和土壤特性的影响[J]. 草地学报,2024,32(8):2659-2669
[8]"""""" 王锐,李希来,张静. 四种覆土处理对高寒煤矿区排土场渣山植被恢复的影响[J]. 草业学报,2020,29(7):40-51
[9]"""""" 安国荣,张明,周冰,等. ICP-MS检测青海省海西州地区羊板粪中的重金属元素含量[J]. 光谱实验室,2012,29(3):1912-1914
[10]"""" 赵宇,李思宇,董博文,等. 鲜羊粪添加对中度盐碱退化羊草草地植被及土壤特征的影响[J]. 中国草地学报,2023,45(9):57-65
[11]"""" 张紫轩,周颖莹,成展仪,等. 碳中和背景下牛羊粪便资源化利用关键技术及模式创新[J]. 家畜生态学报,2024,45(6):88-92
[12]"""" 熊朝阳,张青松,李佳秀,等. 冻融作用下藏北高原牦牛和藏绵羊粪便降解及养分变化特征[J]. 生态环境学报,2023,32(9):1606-1614
[13]"""" 孙子隽,钟国兴,张少搏,等. 化肥减量配施有机肥对植烟土壤理化特性和微生物群落结构的影响[J]. 华北农学报,2024,39(3):146-158
[14]"""" 陈冰,张广娜,王芸,等. 不同有机肥对蓝莓园土壤理化性质及蓝莓光合特性的影响[J]. 中国果菜,2021,41(6):149-154
[15]"""" 高赟,李尚卿,申力行,等. 腐熟羊粪对甘肃中部二阴山区饲用玉米生长特性及饲草品质的影响[J]. 饲料研究,2023,46(21):95-99
[16]"""" 高日平,段玉,张君,等. 长期施肥对农牧交错带旱地土壤微生物多样性及群落结构的影响[J]. 环境科学,2023,44(2):1063-1073
[17]"""" 徐学池,苏以荣,王桂红,等. 秸秆还田配施氮肥对喀斯特农田微生物群落及有机碳矿化的影响[J]. 环境科学,2019,40(6):2912-2919
[18]"""" 杨建强,刁华杰,胡姝娅,等. 氮磷添加对盐渍化草地土壤微生物特征的影响[J]. 环境科学,2021,42(12):6058-6066
[19]"""" 李鹏程,郑苍松,孙淼,等. 土壤全氮含量和施氮水平对棉花产量及氮肥利用效率的影响[J]. 核农学报,2017,31(8):1612-1617
[20]"""" YANG H K, LI J A, WU G, et al. Maize straw mulching with no-tillage increases fertile spike and grain yield of dryland wheat by regulating root-soil interaction and nitrogen nutrition[J]. Soil and Tillage Research,2023,228:105652
[21]"""" JANSSON J K, MCCLURE R, EGBERT R G. Soil microbiome engineering for sustainability in a changing environment[J]. Nature Biotechnology,2023,41(12):1716-1728
[22]"""" WANG M X, GE A H, MA X Z, et al. Dynamic root microbiome sustains soybean productivity under unbalanced fertilization[J]. Nature Communication,2024,15(1):1668
[23]"""" WANG C, LIU D W, BAI E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition[J]. Soil Biology and Biochemistry,2018,120:126-133
[24]"""" FREY B, MOSER B, TYTAGT B, et al. Long-term N-addition alters the community structure of functionally important N-cycling soil microorganisms across global grasslands[J]. Soil Biology and Biochemistry,2023,176:108887
[25]"""" TING J, DUNGAIT J A J, LU X K, et al. Long-term nitrogen addition modifies microbial composition and functions for slow carbon cycling and increased sequestration in tropical forest soil[J]. Global Change Biology,2019,25(10),3267-3281
[26]"""" 鲍士旦. 土壤农化分析[M]. 第3版. 北京:中国农业出版社,2005:22-187
[27]"""" YAN G Y, XING Y J, HAN S J, et al. Long-time precipitation reduction and nitrogen deposition increase alter soil nitrogen dynamic by influencing soil bacterial communities and functional groups[J]. Pedosphere,2020,30:363-377
[28]"""" KOMY Z R. Comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soils[J]. International Journal of Environmental Analytical Chemistry,1995,60:41-47
[29]"""" FU B J, LIU S L, CHEN D L, et al. Soil quality regime in relation to land cover and slope position across a highly modified slope landscape[J]. Ecological Research,2004,19:111-118
[30]"""" LI H Y, QIU Y Z, YAO T, et al. Nutrients available in the soil regulate the changes of soil microbial community alongside degradation of alpine meadows in the northeast of the Qinghai-Tibet Plateau[J]. Science of the Total Environment,2021,792:148363
[31]"""" XIONG W, ZHAO Q Y, ZHAO J, et al. Different continuous cropping spans significantly affect microbial community membership and structure in a Vanilla-Grown soil as revealed by deep pyrosequencing[J]. Microbial Ecology,2015,70:209-218
[32]"""" CHENG B J, WANG Z Y, YAN X Q, et al. Characteristics and pollution risks of Cu, Ni, Cd, Pb, Hg and As in farmland soil near coal mines[J]. Soil amp; Environmental Health,2023,1:100035
[33]"""" 高志香,李希来,张静,等. 不同施肥处理对高寒矿区渣山改良土酶活性和理化性质的影响[J]. 草地学报,2021,29(8):1748-1756
[34]"""" 孔令健,张琳,任杰,等. 高寒矿区环境材料混施下土壤改良及工程应用研究[J]. 煤炭科学技术,2024,52(S1):299-311
[35]"""" 高浩诚,牧仁,焦婷,等. 施氮对高寒垂穗披碱草草地根际、非根际土壤肥力的影响[J]. 水土保持学报,2023,37(6):276-282
[36]"""" 张玉芳,李希来,高志香,等. 不同施肥组合对木里矿区人工植被和土壤微生物特征的影响[J]. 西北农业学报,2022,31(6):741-754
[37]"""" 刘万杰. 有机肥用量和播种方式对高寒矿区渣山表层基质、植被生长和微生物的影响[D]. 西宁:青海大学,2023:22-31
[38]"""nbsp; 王光华,胡晓婧,于镇华,等. 施肥对我国黑土农田土壤微生物群落多样性影响的研究及展望[J]. 土壤与作物,2024,13(2):127-139
[39]"""" WANG J J, SUN X, DU L N, et al. Appropriate fertilization increases carbon and nitrogen sequestration and economic benefit for straw-incorporated upland farming[J]. Geoderma,2024,441:116743
[40]"""" 温烜琳,马宜林,周俊学,等. 腐熟羊粪有机肥配施无机肥对植烟土壤微生物群落结构和多样性的影响[J]. 土壤,2023,55(4):804-811
[41]"""" LI T, WANG S C, LIU C E, et al. Soil microbial communities' contributions to soil ecosystem multifunctionality in the natural restoration of abandoned metal mines[J]. Journal of Environmental Management,2024,353:120244
[42]"""" 侯彦林,王曙光,郭伟. 尿素施肥量对土壤微生物和酶活性的影响[J]. 土壤通报,2004,35(3):303-306
[43]"""" 马欣,罗珠珠,张耀全,等. 黄土高原雨养区不同种植年限紫花苜蓿土壤真菌群落的分布特征[J]. 干旱地区农业研究,2021,39(3):162-170
[44]"""" 谭勇,崔尹赡,季秀玲,等. 三七连作的根际、根内微生物变化与生态学研究进展[J]. 中草药,2017,48(2):391-399
[45]"""" 周学丽,史德军,刘汉江,等. 有机肥改变了退化人工草地微生物群落网络结构和潜在功能[J]. 草地学报,2023,31(9):2693-2702
[46]"""" 王占青,王午志,马喆,等. 不同禾草生长对甘肃马先蒿群落土壤养分和微生物群落的影响[J]. 草地学报,2023,31(12):3750-3758
[47]"""" ZHANG G L, BAI J H, JIA J, et al. Soil microbial communities regulate the threshold effect of salinity stress on SOM decomposition in coastal salt marshes[J]. Fundamental Research,2023,3(6):868-879
[48]"""" 焦嘉卉,符晓,张硕,等. 不同林龄花椒根际土壤理化性质与微生物群落结构的分析[J]. 西北林学院学报,2023,38(4):156-165
[49]"""" 闫丰,赵鑫,邵丽君,等. 冀北采石场恢复区不同植被恢复类型对土壤微生物群落结构的影响[J/OL].https://doi.org/10.13227/j.hjkx.202402063, 2024-05-14/2024-11-12
[50]"""" 童永尚,张春平,俞旸,等. 多年生高寒栽培草地土壤微生物学特性对短期氮添加的响应[J/OL].https://doi.org/10.13227/j.hjkx.202312228, 2024-04-03/2024-11-12
(责任编辑" 彭露茜)
猜你喜欢
山东农业科学(2017年3期)2017-03-29 17:09:55
绿色科技(2017年3期)2017-03-14 20:27:19
吉林农业·下半月(2017年2期)2017-03-10 12:41:59
现代农业科技(2016年20期)2016-12-20 08:54:23
江苏农业科学(2016年7期)2016-10-20 16:07:34
广西农学报(2016年1期)2016-10-11 11:05:43
天津农业科学(2016年4期)2016-04-20 15:24:10
河北渔业(2015年5期)2016-03-01 16:21:29
安徽农学通报(2015年19期)2015-10-23 01:11:53
江苏农业科学(2015年3期)2015-07-31 00:55:39
