NLRP6过表达通过AMPK/CPT1A/PGC1A通路促进肝细胞脂肪氧化分解改善非酒精性脂肪肝
2025-02-06 00:00:00石情冉苏叶宋铃榆杨红王文娟刘晗琳刘琦
南方医科大学学报 2025年1期
关键词:非酒精性脂肪肝


摘要:目的 研究核苷酸结合寡聚化结构域样受体含pyrin 结构域蛋白6(NLRP6)在肝脏脂代谢及非酒精性脂肪性肝病(NAFLD)发展过程中的作用及机制。方法 构建高脂饲料喂养(HFD)模型小鼠,分为对照组(正常喂养)、模型组[高脂(脂质含量60%饮食)喂养],6 只/组,持续喂养16 周。构建蛋氨酸胆碱缺乏饮食(MCD)小鼠模型,分为对照组[喂养蛋氨酸-胆碱充足(MCS)饮食]和模型组(喂养蛋氨酸-MCD饮食),6只/组,持续喂养8周。使用HE染色及油红O染色鉴定模型是否造模成功,免疫组化染色(IHC)检测两种小鼠模型肝脏中NLRP6 的表达。在正常人肝细胞系LO2 细胞中使用腺病毒过表达 NLRP6,或siRNA 敲低 NLRP6,使用棕榈酸(PA)构建NAFLD细胞模型,NLRP6 过表达处理分组Ad-Vec 组、Ad-NLRP6 组、Ad-Vec+PA组、Ad-NLRP6+PA组;NLRP6 敲低处理分组阴性对照(NC)组、sh-NLRP6 组、NC+PA组、sh-NLRP6+PA组。采用甘油三酯(TG) 试剂盒、油红染色、qPCR、Western blotting、ATP 试剂盒和 β-羟基丁酸试剂盒检测NLRP6对脂质代谢的影响及NLRP6对AMPK通路及使用AMPK抑制剂Compound C后AMPK通路的变化。结果 在HFD、MCD饮食诱导NAFLD小鼠模型小鼠肝脏中NLRP6 表达降低(Plt;0.001)。在细胞实验中LO2 细胞过表达NLRP6 后,与Ad-Vec+PA组相比,Ad-NLRP6+PA组细胞内TG含量降低(Plt;0.0001),且脂质沉积减少;敲低NLRP6,与NC+PA组相比,sh-NLRP6+PA组细胞内TG含量增加(Plt;0.05),并且脂质沉积加重;与Ad-Vec+PA组相比,Ad-NLRP6+PA组PGC1A、CPT1A mRNA及蛋白表达水平增加(Plt;0.01)。脂质氧化分解代谢产物ATP、β-羟丁酸含量升高(Plt;0.05)。与Ad-Vec+PA组相比,Ad-NLRP6+PA组肝细胞中AMPK通路磷酸化水平升高(Plt;0.05);使用AMPK抑制剂后,Ad-NLRP6引起的脂质氧化分解作用被抑制关。结论 NLRP6通过AMPK/CPT1A/PGC1A促进肝细胞脂肪氧化分解,改善脂质沉积。
关键词:非酒精性脂肪肝;NLRP6;AMPK;脂肪酸氧化
非酒精性脂肪性肝病(NAFLD)是与肥胖、胰岛素抵抗、高血压、高血脂等代谢功能紊乱相关的疾病,全世界患病率约为25%,是最常见的慢性肝病[1],同时也是慢性肝病进展至肝硬化和肝癌的重要过程[2]。NAFLD的主要病理特征包括肝细胞水肿、肝细胞脂肪变性、炎症、同时可伴有不同程度的肝纤维化[3]。NAFLD的病理进程在一定程度上是可逆的,因此早期干预NAFLD的进展,对于延缓或者阻止慢性肝脏疾病发展成为肝硬化、肝癌等终末期肝病具有重要意义。但NAFLD发病机制复杂,目前并没有特效治疗药物或生物制剂[4],因此,深入探索NAFLD的发生发展机制,对于该病的临床治疗策略的研发具有重要意义。
核苷酸结合寡聚化结构域样受体含pyrin结构域蛋白6(NLRP6)是形成炎性小体的新型nod样受体(NLR)家族成员,在肝脏、肠道中高表达[5],并且可以在肠道炎症、肺炎等疾病中发挥抗炎作用[6-8]。但NLRP6在肝脏中的作用及参与NAFLD 进程的机制尚不清楚。NLRP6 与非酒精性脂肪肝相关仅有1 项研究发现NLRP6-/-小鼠能通过抑制NF-κB通路炎症改善NASH[9],但NLRP6对肝细胞直接的作用及对肝细胞脂质代谢过程的影响尚不清楚。为了阐明NLRP6在肝细胞脂质代谢中的作用,本研究构建高脂饲料喂养(HFD)模型小鼠和蛋氨酸胆碱缺乏饮食(MCD)小鼠模型,体内外实验检测肝脏中NLRP6的表达,并通过体外细胞实验,发现NLRP6可能通过激活AMPK通路上调PGC1A、CPT1A水平,促进肝细胞脂质氧化分解,从而改善NAFLD,为NAFLD提供新的治疗思路及实验基础。
1 材料和方法
1.1 材料与试剂
棕榈酸(PA,Sigma),DMEM(Gibico)免疫组化(IHC) 染色试剂盒、β-羟丁酸检测试剂盒(CaymanChemical);ATP检测试剂盒(上海碧云天生物技术有限公司);组织细胞三酰甘油(TG)酶法测定试剂盒(北京普利莱基因技术有限公司);油红染色测试盒(南京建成生物工程研究所有限公司);GAPDH抗体、AMPK抗体、PGC1A抗体、HRP偶联的山羊抗小鼠IgG(H+L)(武汉三鹰生物技术有限公司)。p-AMPK 抗体(CST)。NLRP6 抗体(Sigma),NLRP6 抗体(ImmunoWay),CPT1A抗体(Abcam),辣根酶标记山羊抗兔IgG(H+L)(中杉金桥)。
1.2 细胞培养
人LO2细胞(BNCC),用含10%胎牛血清和1%双抗的DMEM培养于37 ℃,5% CO2培养箱内。使用腺病毒(2.765×106 PFU/mL)或siRNA(5 nmol/mL)分别构建过表达与敲低NLRP6,用PA 0.5 mmol/L处理LO2细胞48 h构建NAFLD细胞模型[10]。细胞实验分组:Ad-Vec组、Ad-NLRP6 组、Ad-Vec+PA组、Ad-NLRP6+PA组、阴性对照(NC)组、sh-NLRP6 组、NC+PA 组、sh-NLRP6+PA组,各组均用对应腺病毒/siRNA转染24 h后换液,Ad-NLRP6+PA组、Ad-NLRP6+PA组、NC+PA组、sh-NLRP6+PA组加PA处理48 h 后收样。Ad-Vec组、Ad-NLRP6 组、NC 组、sh-NLRP6 组加同等体积5%BSA处理48 h后收样。
1.3 动物实验
本研究动物实验经贵州医科大学动物伦理委员会审查批准(伦理批号:2000639)。使用6 周龄C57BL/6雄性小鼠,分笼饲养于相对湿度40%、温度约 23±2 ℃清洁级动物房,自由进食、进水。将小鼠适应性喂养1周,按随机分组。HFD模型小鼠构建:分为ND组、HFD组,ND组正常喂养,HFD组高脂(脂质含量60%)饮食喂养,6只/组,持续喂养16周,每周记录小鼠体质量变化。MCD小鼠模型构建:分为胆碱充实(MCS)组、MCD组。MCS 组喂养蛋氨酸-MCS 饮食,MCD 组喂养蛋氨酸-MCD饮食,6 只/组,持续喂养8 周,每周记录小鼠体质量变化。戊巴比妥钠麻醉小鼠(50 mg/kg剂量腹腔注射),取血后处死。记录肝质量,收集组织进行后续实验。
1.4 检测指标与检测方法
1.4.1 免疫组化 小鼠肝脏组织制成厚度为3 μm石蜡切片,经烤片(70 ℃,30 min)、二甲苯脱蜡、水化、阻断内源性过氧化物酶、柠檬酸盐溶液抗原修复,室温封闭,滴加鼠兔抗小鼠NLRP6 抗体(1∶50),4℃孵育过夜,二抗增强剂37 ℃ 20 min,二抗(1∶200)孵37 ℃ 30 min,3,3'-二氨基联苯胺(DAB)显色,苏木素染色液复染,脱水,二甲苯透明,封片,光学显微镜观察。随机选取5个400倍视野进行图片采集。
1.4.2 油红O染色 小鼠肝组织使用OCT包埋,制成厚度为10 μm的冰冻切片进行后续油红O染色。使用油红O染色试剂盒染色,光学显微镜观察。随机选取5个400倍视野进行图片采集。
1.4.3 Western blotting 取小鼠肝组织于组织匀浆管中加入匀浆磁珠并混合配制好的蛋白裂解液,5000 r/min低温匀浆、裂解30 min、超声破碎,12 000×g离心30 min,取上清,使用BCA(Beyotime)法测定蛋白浓度,加入5×上样缓冲液,100 ℃水浴5 min;使用裂解液将细胞冰上裂解1 h,12 000×g离心15 min,取上清,使用BCA法测定蛋白浓度,5×上样缓冲液,100 ℃水浴5 min。加入将备好的蛋白样本进行聚丙烯酰胺凝胶电泳(80~120 V),转移至PVDF 膜上,经过脱脂牛奶封闭、一抗孵育4 ℃过夜[(GAPDH 1∶10 000)、(AMPK 1∶8000)、(PGC1A 1∶1000)、(p-AMPK 1∶1000)、(NLRP6 1∶1000)、(CPT1A 1∶1000)]、二抗(1∶5000)孵育1h后采用化学发光成像分析系统采集图片。
1.4.4 RT-PCR 采用TRIzol 法提取总RNA,使用分光光度计(Nanodrop,Thermo)进行RNA浓度测定及质量评估;取500 ng 总RNA使用逆转录试剂盒(Takara)逆转录为cDNA,使用SYBR Green Master Mix(11184ES08)进行qPCR检测。引物由生工生物工程有限公司合成(表1)。
1.4.5 生化指标测定 细胞TG、ATP、β-羟丁酸测定使用相应试剂盒进行检测。
1.5 统计学分析
使用 SPSS 26.0 统计软件及Grapgpad Prism 9 进行数据分析及处理。计量资料符合正态分布以均数±标准差表示,两组间比较采用独立样本t 检验;多组间比较采用单因素方差分析,其分组间两两比较采用LSD法;计量资料若不符合正态分布,采用M(P25,P75)进行描述,组间比较采用秩和检验。计数资料用百分比(%)表示,组间比较采用χ2检验;Plt;0.05 为差异具有统计学意义。
2 结果
2.1 NLRP6在NAFLD小鼠模型肝脏中表达降低
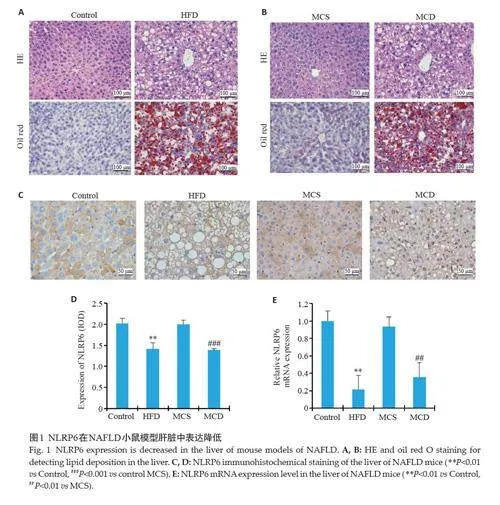
HE 染色见HFD组及MCD组小鼠肝细胞明显脂肪变,油红O染色见大量脂滴沉积(图1A、B),验证造模成功。NLRP6 mRNA在HFD、MCD组中表达降低(Plt;0.01,图1E),NLRP6主要表达于细胞质中,且与对应对照组相比,HFD、MCD组肝脏中NLRP6的蛋白表达降低(Plt;0.01,图1C、D)。
2.2 过表达NLRP6可改善PA构建的NAFLD细胞模型的脂质沉积
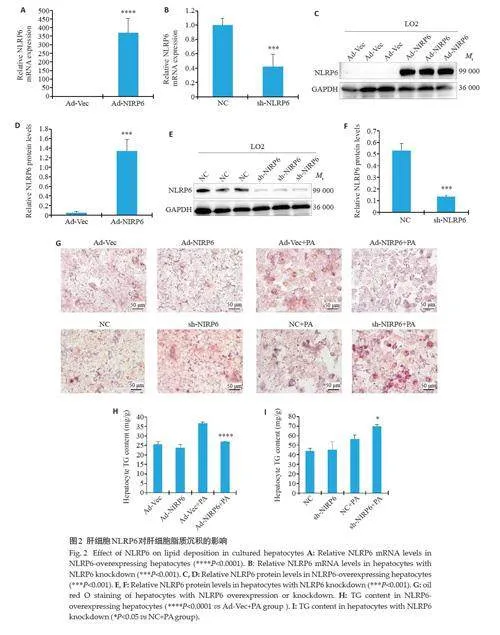
肝细胞中NLRP6 过表达NLRP6 mRNA 表达水平(Plt;0.0001,图2A)及蛋白表达水平升高(Plt;0.001,图2C、D),肝细胞NLRP6 敲低NLRP6 mRNA表达水平(Plt;0.001,图2B)及蛋白表达水平降低(Plt;0.001,图2E、F)。油红O染色结果显示过表达NLRP6 改善PA诱导的细胞脂质沉积,而敲低NLRP6 结果则相反(图2G)。采用TG检测试剂盒检测肝细胞TG结果显示在过表达NLRP6 时,与Ad-Vec+PA 处理组相比,Ad-NLRP6+PA组TG水平降低(Plt;0.0001,图2H)。在敲低NLRP6时,与NC+PA组相比,sh-NLRP6+PA组细胞TG含量升高(Plt;0.05,图2I)。
2.3 NLRP6通过CPT1A/PGC1A促进肝细胞脂肪酸氧化分解
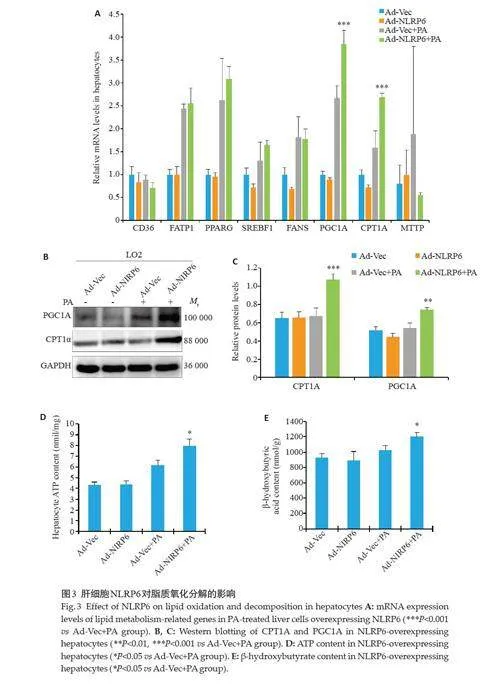
Ad-NLRP6+PA 组中脂肪酸氧化分解相关基因CPT1A、PGC1A mRNA 表达水平升高(Plt;0.001,图3A),同时WB检测也发现CPT1A、PGC1A蛋白水平升高(Plt;0.001,Plt;0.01,图3B、C)。Ad-NLRP6+PA 组细胞ATP(Plt;0.05,图3D)、β-羟丁酸含量高于Ad-Vec+PA组(Plt;0.05,图3E)。
2.4 NLRP6激活AMPK通路调控脂质氧化
过表达NLRP6 后,与Ad-Vec+PA组相比,AMPK磷酸化水平升高(Plt;0.05,图4A、B)。使用AMPK抑制剂Compound C后,PA处理条件下,与Ad-NLRP6+PA组相比Ad-NLRP6+PA+Com组p-AMPK、CPT1A、PGC1A蛋白表达水平降低(Plt;0.001,图4C~F),肝细胞ATP水平及β-羟丁酸水平也降低(Plt;0.001,图4G、H)。
3 讨论
NLRP6可参与组成炎症小体,在癌症、免疫等发挥保护作用,并在肝癌中发挥重要的作用,当NLRP6缺失时,可促进肝癌的进展[11]。我们在PA构建的NAFLD细胞模型中发现,过表达NLRP6后,肝细胞TG含量明显降低,油红染色结果也表明细胞脂质沉积明显改善。在PA处理的LO2细胞中敲低NLRP6时,则显示的是相反的结果,表明NLRP6可能有改善肝细胞脂质沉积的作用,为NAFLD的治疗提供了新的干预靶点。
进一步探索NLRP6 改善NAFLD 的具体作用机制,由于肝细胞脂质沉积的发生主要与以下4个环节密切相关:肝细胞摄取的游离脂肪酸(FFA)增加;肝细胞纸质重头合成产生的FFA增加;FFA氧化分解过程受阻;肝细胞FFA 合成TG 以后,TG 转运出肝细胞减少[12-15]。为了进一步探索NLRP6如何影响脂质代谢,我们对参与上述4 个过程的相关基因进行检测。我们将过表达NLRP6的LO2细胞使用PA处理,使用qPCR检测脂质代谢相关基因mRNA的表达水平。我们发现,NLRP6 在脂质分解代谢过程中发挥重要作用,过表达NLRP6 会增加PGC1A、CPT1A 的表达。PGC1A、CPT1A 在脂质代谢过程中参与脂肪酸氧化分解阶段[16-18],CPT1A是脂肪酸氧化的限速酶,它可使长链的乙酰辅酶A进入线粒体基质内,参与三羧酸循环,促进脂肪酸β氧化[19]。通过上调CPT1A的表达,在转录水平刺激β氧化,降低脂肪酸的水平,抑制TG合成[20]。有研究发现当PGC1A在小鼠原代肝细胞中过表达时,TG合成减少,脂肪酸氧化分解增加,并且发现PGC1A可刺激过氧化物酶体活性,并提升人原代肌管中的长链和超长链脂肪酸氧化分解[21]。在本研究中过表达NLRP6组肝细胞中脂质代谢产物ATP、β-羟丁酸水平显著升高,进一步证明了NLRP6能通过调控PGC1A、CPT1A表达改善肝脂肪变性。NLRP6主要参与调控脂质代谢过程中脂质氧化分解环节,其通过促进肝细胞脂质氧化分解,从而减少细胞中的脂质沉积。
AMPK是能量传递和代谢的重要枢纽[22],参与调控脂质氧化分解,在非酒精脂肪肝中发挥重要的作用[23]。我们推测NLRP6 是否可以通过AMPK通路影响脂质代谢。我们发现NLRP6 可上调AMPK磷酸化水平。AMPK是一种异源三聚体丝氨酸/苏氨酸激酶,有细胞能量状态的传感器的作用,AMPK被激活后促进ATP生成,促进脂质氧化分解,在脂质氧化分解中起到关键性作用[24-27]。在本研究中,在PA构建的NAFLD细胞模型中,过表达NLRP6 后,肝细胞中ATP 水平升高;且AMPK可以通过减轻丙二酰辅酶A对CPT1A的抑制作用,从而促进脂肪酸氧化分解[27]。活化的AMPK增加了PGC1A的转录,最终改善线粒体功能、能量代谢和胰岛素抵抗[28, 29],这与本研究结果一致,在本研究中NLRP6增加AMPK磷酸化水平,并且CPT1A、PGC1A的变化水平升高。当使用AMPK抑制剂后,过表达NLRP6促进AMPK磷酸化水平作用消失,且CPT1A、PGC1A蛋白水平降低,表明NLRP6能通过AMPK通路影响脂质氧化分解。
NLRP6如何调控AMPK的激活,可能通过直接或者间接影响LKB1、STRAD和MO25 组成的异源三聚体、CAMKK2或细胞内钙离子的变化调控AMPK的激活,但是具体如何调控AMPK变化需进一步阐明。在既往研究中,AMPK激活可以调节炎症小体如NLRP3、NLRC4,那么AMPK是否也可以反向作用影响NLRP6有待进一步研究[30, 31]。我们在筛选NLRP6 如何影响TG沉积的时候对脂质代谢不同的过程进行了筛选,但本研究中只筛选了mRNA水平,并没有对这些基因的蛋白水平进行检测,这些没有明显变化的基因,是否有存在转录后修饰或蛋白水平调控的调控方式,最终影响它们的表达尚需进一步研究。
综上所述,本研究NLRP6可能通过激活AMPK通路上调PGC1A、CPT1A水平,促进肝细胞脂质氧化分解,从而改善NAFLD,为NAFLD提供新的治疗思路及实验基础。
参考文献:
[1] Powell EE, Wong VW, Rinella M. Non-alcoholic fatty liver disease
[J]. Lancet, 2021, 397(10290): 2212-24.
[2] Younossi ZM. Non-alcoholic fatty liver disease-A global public
health perspective[J]. J Hepatol, 2019, 70(3): 531-44.
[3] Sanyal AJ, van Natta ML, Clark J, et al. Prospective study of
outcomes in adults with nonalcoholic fatty liver disease[J]. N Engl J
Med, 2021, 385(17): 1559-69.
[4] Rong L, Zou JY, Ran W, et al. Advancements in the treatment of nonalcoholic
fatty liver disease (NAFLD) [J]. Front Endocrinol, 2023,
13: 1087260.
[5] Li RZ, Zhu S. NLRP6 inflammasome[J]. Mol Aspects Med, 2020,
76: 100859.
[6] Zheng DP, Kern L, Elinav E. The NLRP6 inflammasome[J].
Immunology, 2021, 162(3): 281-9.
[7] Tao Q, Xu DY, Jia KX, et al. NLRP6 serves as a negative regulator
of neutrophil recruitment and function during Streptococcus
pneumoniae infection[J]. Front Microbiol, 2022, 13: 898559.
[8] Mukherjee S, Kumar R, Tsakem Lenou E, et al. Deubiquitination of
NLRP6 inflammasome by Cyld critically regulates intestinal
inflammation[J]. Nat Immunol, 2020, 21(6): 626-35.
[9] Huang CY, Liu QH, Tang Q, et al. Hepatocyte-specific deletion of
Nlrp6 in mice exacerbates the development of non-alcoholic
steatohepatitis[J]. Free Radic Biol Med, 2021, 169: 110-21.
[10]Frietze KK, Brown AM, Das D, et al. Lipotoxicity reduces DDX58/
Rig-1 expression and activity leading to impaired autophagy and cell
death[J]. Autophagy, 2022, 18(1): 142-60.
[11] Schneider KM, Mohs A, Gui WF, et al. Imbalanced gut microbiota
fuels hepatocellular carcinoma development by shaping the hepatic
inflammatory microenvironment[J]. Nat Commun, 2022, 13(1):
3964.
[12]Badmus OO, Hillhouse SA, Anderson CD, et al. Molecular
mechanisms of metabolic associated fatty liver disease (MAFLD):
functional analysis of lipid metabolism pathways[J]. Clin Sci, 2022,
136(18): 1347-66.
[13]Yki-Järvinen H, Luukkonen PK, Hodson L, et al. Dietary
carbohydrates and fats in nonalcoholic fatty liver disease[J]. Nat
Rev Gastroenterol Hepatol, 2021, 18(11): 770-86.
[14]Machado MV, Diehl AM. Pathogenesis of nonalcoholic
steatohepatitis[J]. Gastroenterology, 2016, 150(8): 1769-77.
[15] Ipsen DH, Lykkesfeldt J, Tveden-Nyborg P. Molecular mechanisms
of hepatic lipid accumulation in non-alcoholic fatty liver disease[J].
Cell Mol Life Sci, 2018, 75(18): 3313-27.
[16]Yamamoto T, Takabatake Y, Minami S, et al. Eicosapentaenoic acid
attenuates renal lipotoxicity by restoring autophagic flux[J].
Autophagy, 2021, 17(7): 1700-13.
[17]Li K, Gao L, Zhou S, et al. Erythropoietin promotes energy
metabolism to improve LPS-induced injury in HK-2 cells via SIRT1/
PGC1-α pathway[J]. Mol Cell Biochem, 2023, 478(3): 651-63.
[18]Helsley RN, Park SH, Vekaria HJ, et al. Ketohexokinase-C regulates
global protein acetylation to decrease carnitine palmitoyltransferase
1a-mediated fatty acid oxidation[J]. J Hepatol, 2023, 79(1): 25-42.
[19]Sun W, Nie T, Li K, et al. Hepatic CPT1A facilitates liver-adipose
cross-talk via induction of FGF21 in mice[J]. Diabetes, 2021:
db210363.
[20]Schlaepfer IR, Joshi M. CPT1A-mediated fat oxidation,
mechanisms, and therapeutic potential[J]. Endocrinology, 2020, 161
(2): bqz046.
[21]Qian L, Zhu Y, Deng C, et al. Peroxisome proliferator-activated
receptor gamma coactivator-1 (PGC-1) family in physiological and
pathophysiological process and diseases[J]. Signal Transduct Target
Ther, 2024,9(1):50.
[22]Trefts E, Shaw RJ. AMPK: restoring metabolic homeostasis over
space and time[J]. Mol Cell, 2021, 81(18): 3677-90.
[23]Diniz TA, de Lima Junior EA, Teixeira AA, et al. Aerobic training
improves NAFLD markers and insulin resistance through AMPKPPAR-
α signaling in obese mice[J]. Life Sci, 2021, 266: 118868.
[24]Herzig S, Shaw RJ. AMPK: guardian of metabolism and
mitochondrial homeostasis[J]. Nat Rev Mol Cell Biol, 2018, 19(2):
121-35.
[25]Lin SC, Hardie DG. AMPK: sensing glucose as well as cellular
energy status[J]. Cell Metab, 2018, 27(2): 299-313.
[26]Steinberg GR, Hardie DG. New insights into activation and function
of the AMPK[J]. Nat Rev Mol Cell Biol, 2023, 24(4): 255-72.
[27]Nakamura MT, Yudell BE, Loor JJ. Regulation of energy
metabolism by long-chain fatty acids[J]. Prog Lipid Res, 2014, 53:
124-44.
[28]Fang CQ, Pan JH, Qu N, et al. The AMPK pathway in fatty liver
disease[J]. Front Physiol, 2022, 13: 970292.
[29]Turkieh A, Caubère C, Barutaut M, et al. Apolipoprotein O is
mitochondrial and promotes lipotoxicity in heart[J]. J Clin Investig,
2014, 124(5): 2277-86.
[30]Bai DS, Du JY, Bu XM, et al. ALDOA maintains NLRP3
inflammasome activation by controlling AMPK activation[J].
Autophagy, 2022, 18(7): 1673-93.
[31]He YT, Xu K, Wang Y, et al. AMPK as a potential pharmacological
target for alleviating LPS-induced acute lung injury partly via
NLRC4 inflammasome pathway inhibition[J]. Exp Gerontol, 2019,
125: 110661.
(编辑:吴锦雅)
猜你喜欢
中国医药导报(2016年33期)2017-03-06 17:15:56
中外医疗(2016年33期)2017-03-02 20:05:29
中国现代医生(2016年30期)2017-03-02 19:09:48
中国当代医药(2016年33期)2017-02-27 14:31:59
医学信息(2016年36期)2017-02-23 14:01:37
中国实用医药(2016年8期)2016-03-30 00:14:15
中国实用医药(2016年4期)2016-02-23 10:54:10
中国当代医药(2015年15期)2015-08-07 16:34:22
中国民族民间医药·下半月(2014年12期)2015-01-15 06:30:46
中国现代医生(2014年33期)2014-12-25 00:55:47