
基于MaxEnt模型的芦苇中国潜在分布区预测
2024-12-31 00:00:00王二国游澜清易自力杜卫红
湖南农业科学 2024年8期



摘要:研究基于338条芦苇分布数据和6个环境变量,利用MaxEnt模型对芦苇在我国的潜在分布区进行模拟,结合环境变量的贡献率、置换重要性、刀切法检验结果以及单变量响应曲线综合分析影响芦苇分布的主要环境变量及其适宜范围。结果表明:MaxEnt模型在RM=1、FC=LQHP组合参数下的AUC均值为0.909,表明预测结果准确度极高;芦苇在我国的潜在分布区主要位于甘肃、山西、陕西等地区,其中高、中适生区面积分别为1.25×109、1.88×109 hm2;影响芦苇分布的主要环境变量是年平均气温、最湿月降水量、等温性、最冷季度降水量和年平均气温变化范围;当年平均气温为14~22℃、最湿月降水量为108~284 mm、等温性低于24、最冷季度降水量小于50 mm、年平均气温变化范围低于28.5℃时,芦苇的分布概率较高。
关键词:芦苇;MaxEnt模型;环境因子;潜在适生区
中图分类号:Q948 文献标识码:A 文章编号:1006-060X(2024)08-0066-07
Prediction of Potential Distribution of Phragmites australis in China Based on MaxEnt
WANG Er-guo1,YOU Lan-qing1,YI Zi-li1,2,DU Wei-hong1
(1. College of Bioscience and Biotechnology, Hunan Agricultural University, Changsha 410128, PRC; 2. Hunan Branch,"National Energy Ramp;D Center for Non-Food Biomass, Changsha 410128, PRC)
Abstract: With 338 data records and 6 environmental variables, this study adopted MaxEnt to predict the potential distribution of Phragmites australis in China. The contribution rate, permutation importance, jackknife test, and univariate response curve were employed to analyze the environmental variables affecting the distribution and the suitable distribution zones of P. australis. With the parameters of RM=1 and FC=LQHP, the MaxEnt model showed the area under the receiver operating characteristic curve of 0.909, which indicated high prediction accuracy. The potential distribution zones of P. australis in China mainly included Gansu, Shanxi,"and Shaanxi, and the highly and moderately suitable distribution areas were 1.25×109 and 1.88×109 hm2, respectively. The main environmental"variables affecting the geographical distribution of P. australis were annual mean temperature, precipitation of the wettest month, isothermality, precipitation of the coldest quarter, and annual mean temperature range. The distribution probability of P. australis was high when the annual mean temperature, precipitation of the wettest month, isothermality, precipitation of the coldest quarter, and annual mean temperature range were 14-22 °C, 108-284 mm, below 24, less than 50 mm, and within mean 28.5 °C, respectively.
Key words: Phragmites australis; MaxEnt; environmental factor; potential suitable distribution zone
芦苇[Phragmites australis(Cav.)Trin. ex Steud.]是禾本科芦苇属的多年生草本植物,也是典型的湿地挺水植物,广泛分布在世界各地[1]。芦苇具有生长速度快、生物质产量高、耐贫瘠等特点,既能通过根茎繁殖迅速扩张群体,也能以种子进行繁殖;其种子尺寸小、质量轻,可以通过风和水远距离传播[2-3]。
而且,芦苇具有广泛的环境耐受性,可以在盐沼中生长[4-5]。研究发现芦苇能吸收水中的有害污染物,净化水体,在保护湿地生态环境和维持湿地生态功能中发挥了重要作用[6-7]。此外,芦苇生物质组成成分中综纤维素含量为76.18%,木素含量为18.24%,是生物能源生产的理想原料[8-9]。当前,已成功用芦苇生产出了纤维素纳米晶体[10]、低聚木糖[11]、乳酸[12]、糠醛[13]等。
生态位模型通过收集物种分布信息和相关的环境变量,借助深度学习算法构建模型,模拟物种的生态需求,并将计算结果在不同的时间和空间进行投影,从而预测物种的潜在分布[14-15]。目前,国内外用常用的预测物种潜在适宜分布区的生态位模型有最大熵模型(MaxEnt)、规则集遗传算法模型(GARP)、生态位因子分析模型(ENFA)、气候匹配模型(CLIMEX)、生物气候分析与预测系统模型(BIOCLIM)等[16-21]。不同模型的理论基础不同,数据要求和分析方法也存在差异。其中,MaxEnt模型具有预测结果稳定、模拟精度高、操作简单、运行时间短、样本需求量小等优点[22-23],被认为是最有效的预测方法之一,且在物种分布研究中得到了广泛应用[24-28]。该研究利用MaxEnt模型预测芦苇在我国的潜在分布区,明确影响芦苇分布范围的主要环境变量,探明芦苇的适宜生境条件,以期为规模化利用芦苇提供理论依据。
1 数据与方法
1.1 数据收集与处理
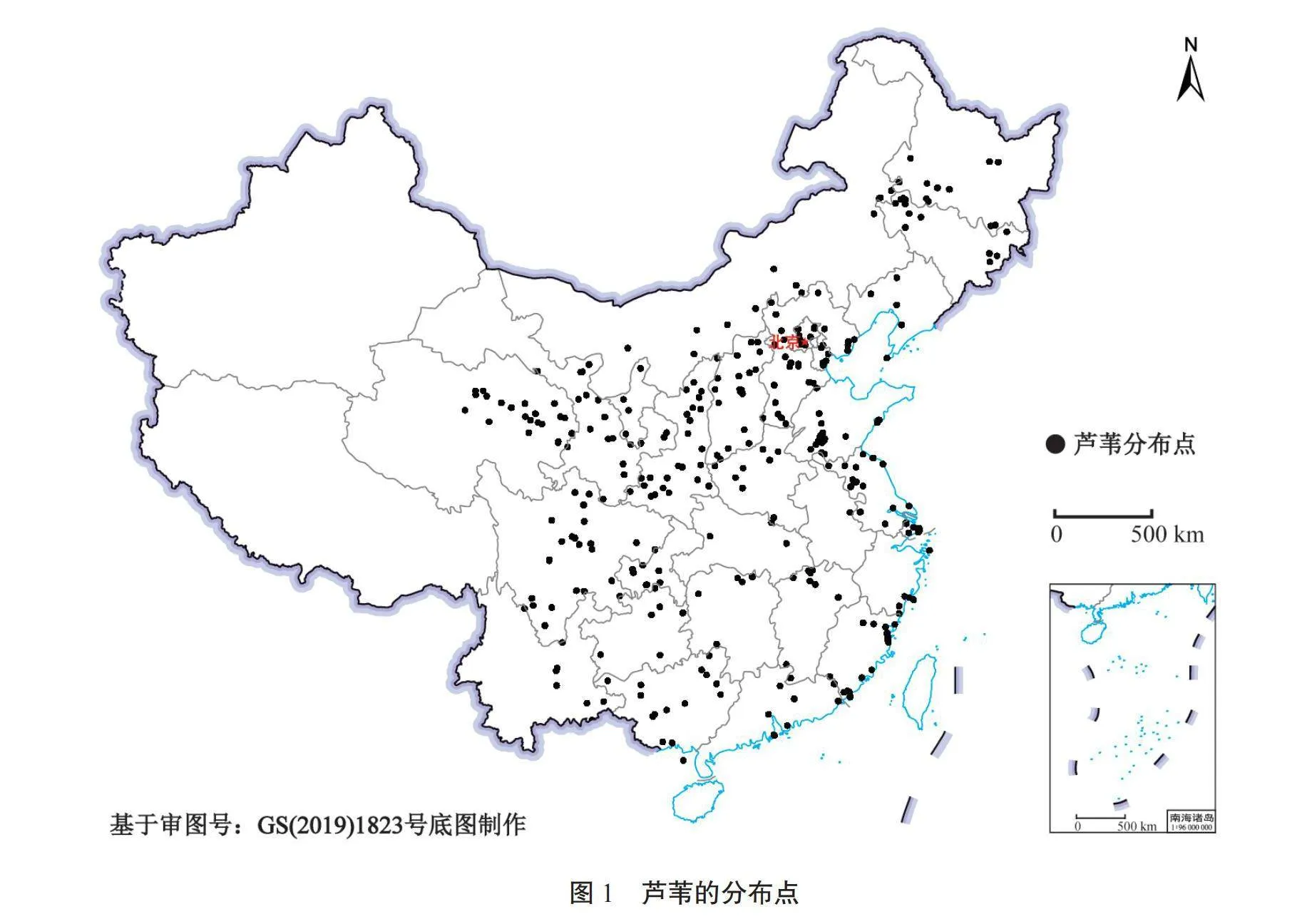
芦苇分布数据来源于全球生物多样性信息机构(GBIF,https://www.gbif.org/zh/)、中国数字植物标本馆(CVH,http://www.cvh.ac.cn/)和中国国家标本资源平台(NSII,http://www.nsii.org.cn/2017/home.php)。通过以上数据源初步获得芦苇现有分布位置,再利用百度坐标拾取系统获取对应的经纬度信息,将种名和分布经纬度以“.csv”格式记录在Excel文件中,删除重复和无效的数据,初步得到401条有效数据。由于MaxEnt模型对物种分布数据的丰度很敏感,而生境适宜性和丰度没有直接关系,为保证每个栅格中芦苇分布数据的唯一性,该研究使用R语言中的spThin包对收集到的分布数据进行稀疏化处理,最终获得了338条分布数据,芦苇的分布点如图1所示。
1.2 环境变量的获取和筛选
该研究涉及19个生物气候变量(表1),均来源于世界气候数据库(Worldclim,http://www.worldclim. org/)。为降低环境变量间的共线性,提高模型的拟合度、准确性以及多变量分析中生态位轴的可解释性,在R中[29]进行皮尔逊相关分析,选择皮尔逊相关系数绝对值|r|≤0.80的环境变量进行后续分析。
1.3 模型参数的选择
优化调控倍频(Regularization multiplier,RM)和特征组合(Feature combination,FC)这两个参数,可显著提高MaxEnt模型预测精度[30-31]。为了优化MaxEnt模型,将RM的变化范围设定为1.0~6.0,每次增加0.5,共11个梯度。MaxEnt模型有5个特征:线性特征(L)、二次型特征(Q)、片段化特征(H)、乘积型特征(P)和阈值特征(T)。FC选取了6种组合:L、LQ、H、LQH、LQHP和LQHPT。根据排列组合,一共66种参数组合。
1.4 模型的构建与评估
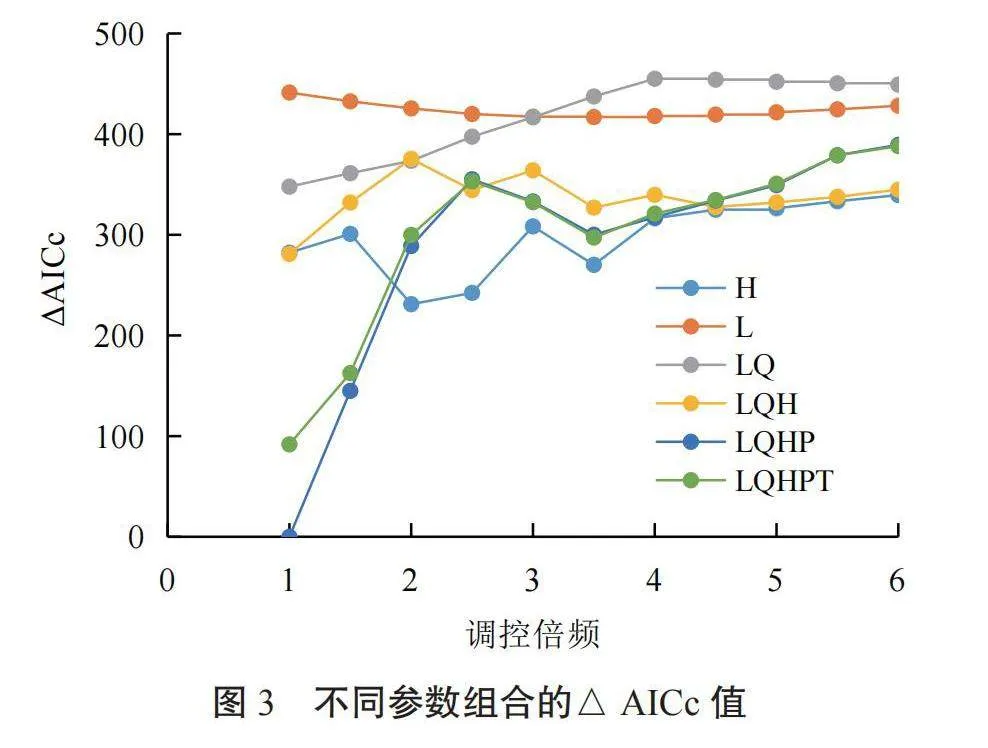
该研究采用MaxEnt软件对芦苇潜在分布区进行预测。将338条芦苇分布信息和筛选得到的环境变量导入MaxEnt软件进行建模。研究借助R的sf包[32]优化模型,首先建立以芦苇分布数据为中心,200 km为半径的缓冲区背景范围;然后利用R的dismo包在背景范围的栅格中随机抽取10 000个背景点,每个栅格最多抽取1个背景点;接着使用ENMeval包中的“checkerboard2”分区块模型对芦苇分布数据和背景点数据进行分区;最后根据Akaike信息准则(Akaike information criterion,AICc)评估不同参数组合下MaxEnt模型的复杂度和拟合度,将△AICc=0时的模型作为推荐模型。为了提高模拟结果的准确性,采用重复子采样和交叉验证方法,重复次数设置为10,取均值进行分析。
使用受试者工作特征(Receiver operating charac-teristic,ROC)曲线下方的面积值AUC来评价MaxEnt模型预测的准确性。AUC越接近1,预测模型的准确性越高[33]。AUC小于0.6表示预测结果不合格,AUC在0.6~0.7表示预测性能差,在0.7~0.8表示预测性能一般,在0.8~0.9表示预测性能良好,在0.9~1.0表示预测性能优异[34-35]。采用刀切法对各环境变量的相对影响进行评价,同时结合各环境变量的贡献率、置换重要性和单变量响应曲线来确定影响芦苇分布的主要环境变量及其适宜范围。
2 结果与分析
2.1 环境变量相关性分析结果
如图2所示,从19个环境变量中选出了6个环境变量,变量间的皮尔逊相关系数绝对值均小于0.8,这6个环境变量分别为Bio1、Bio3、Bio7、Bio13、Bio15、Bio19,即年平均气温、等温性、年平均气温变化范围、最湿月降水量、降水量变异系数和最冷季度降水量。
2.2 模型的优化与准确性评价
在运行MaxEnt模型时,其默认参数设置为RM=1、FC=LQHPT,该参数组合的△AICc=91.79,该值远大于2,表明基于默认参数条件下使用MaxEnt模型进行芦苇适生区预测是不可靠的。由图3可知:在66种组合中,当RM=1、FC=LQHP时,△AICc=0,模型是最优的,因此,选取该参数组合作为模型的最终参数。同时,在该参数组合下利用受试者工作特征曲线进行模型准确性检验,得到AUC均值为0.909,表明预测的芦苇潜在分布区具有较高的准确性。
2.3 影响芦苇分布的主要环境变量
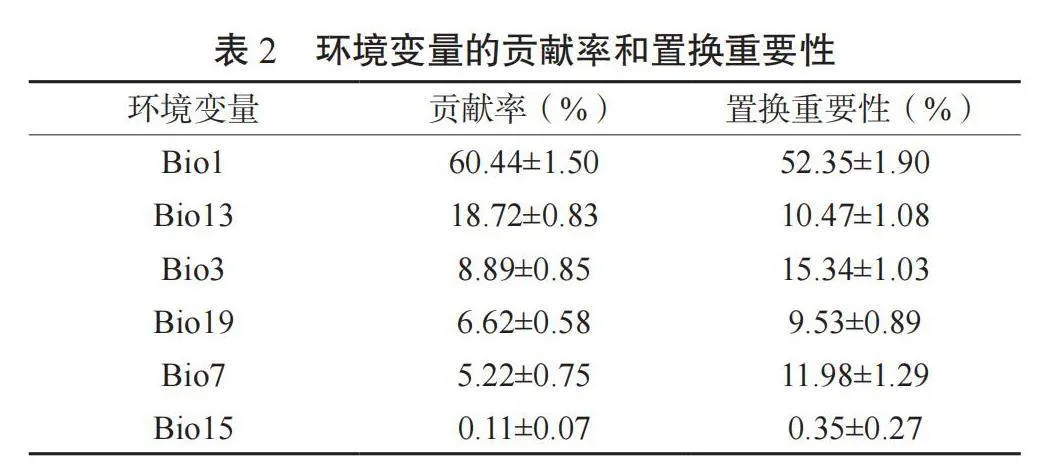
如表2所示,对芦苇分布占主导地位的环境变量有Bio1、Bio3、Bio7、Bio13、Bio15和Bio19,其中有5个环境变量对模型的贡献率超过了5%,分别为Bio1、Bio3、Bio7、Bio13和Bio19,其总贡献率达到了99.89%,总平均置换重要性达到了99.67%。
其中,Bio1置换重要性最高,为52.35%;其次是Bio3和Bio7,分别为15.34%和11.98%。由刀切法分析结果可知(图4),当考虑单一环境变量时,对正规化训练增益影响最大环境因子变量为Bio1(年平均气温),其正规化训练增益和AUC值均最高。综合考虑环境变量的贡献率、置换重要性、刀切法检验结果,可以确认影响芦苇分布的主要环境变量是Bio1(年平均气温)、Bio13(最湿月降水量)、Bio3(等温性)、Bio19(最冷季度降水量)和Bio7(年平均气温变化范围)。
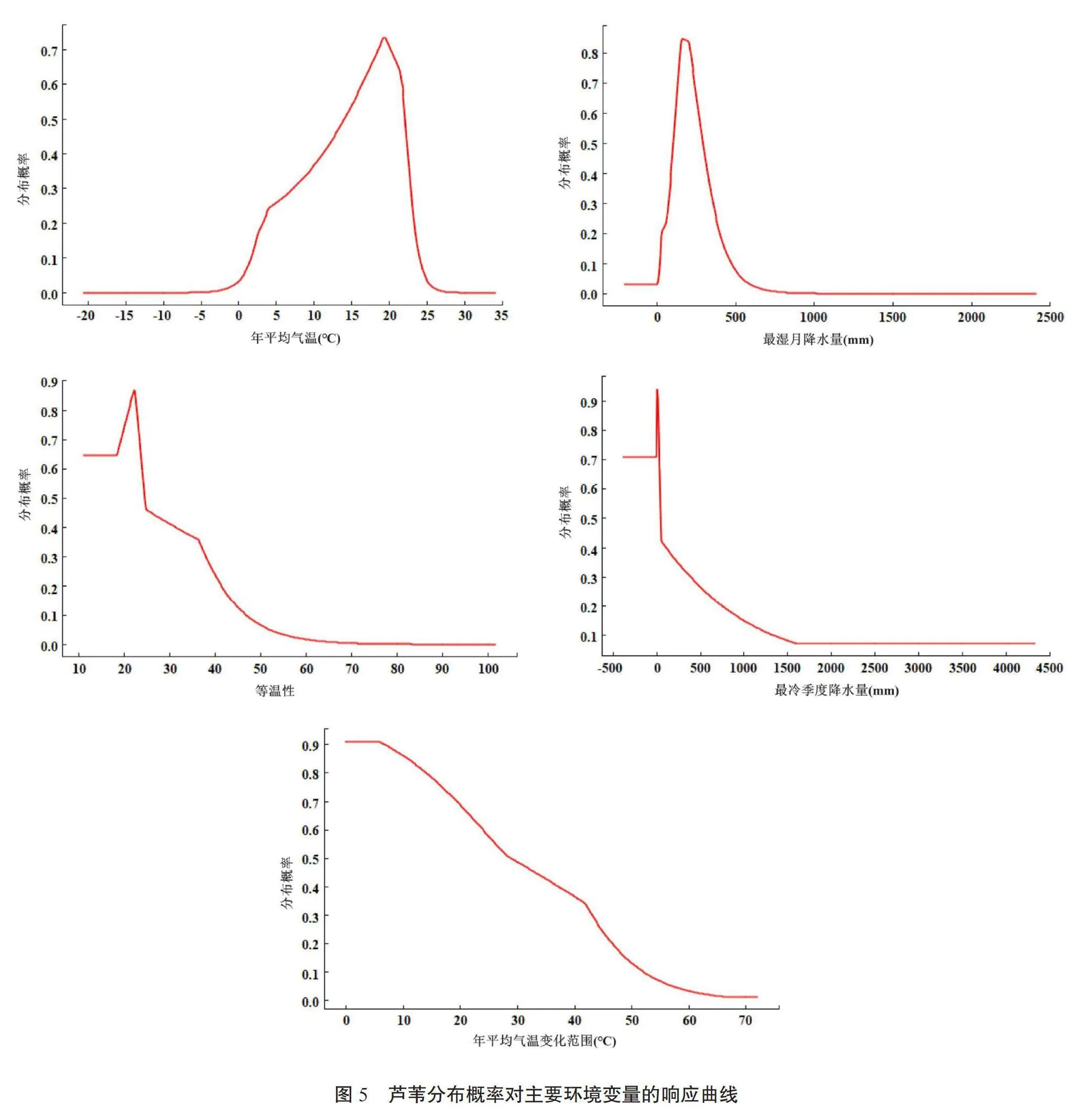
2.4 主要环境变量的适宜区间
将5个主要环境因子导入MaxEnt模型中,建立单因子模型,可分别得到芦苇分布概率对环境变量的单变量响应曲线。当分布概率P>0.5时,认为其所对应的环境变量区间比较适宜芦苇的生存和繁殖。由图5可知主要环境变量的适宜区间分别为:年平均气温14~22℃、最湿月降水量108~284 mm、等温性低于24、最冷季度降水量小于50 mm、年平均气温变化范围低于28.5℃。
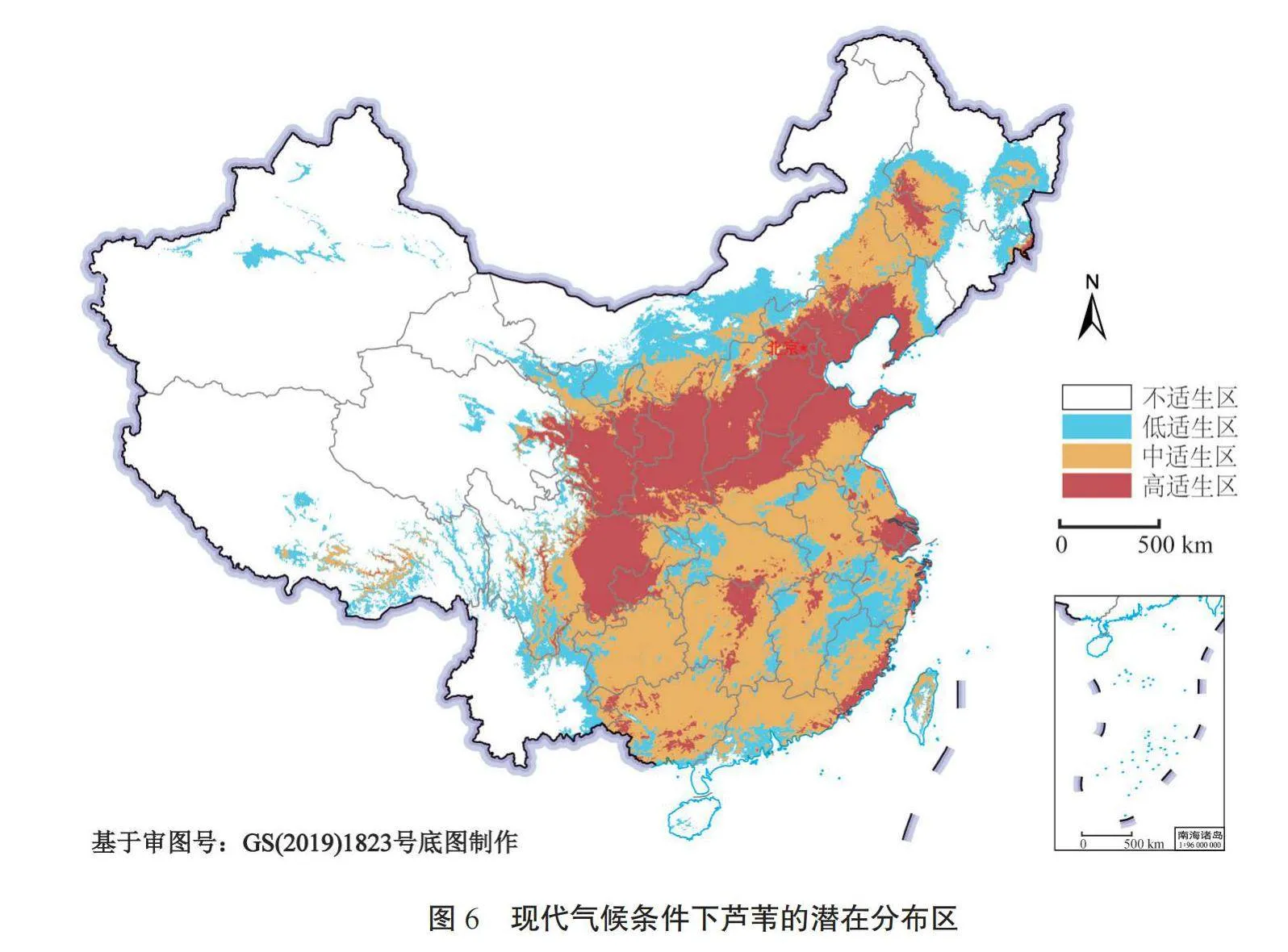
2.5 现代气候条件下的芦苇潜在分布区
由图6可知,现代气候条件下,我国甘肃、山西、陕西、河南北部、河北、北京、天津、山东除南部地区、辽宁西部、宁夏、内蒙古西南部、湖南北部、四川东部、重庆西部、上海、江苏南部、浙江和福建东南部沿海区域为芦苇的高度适生区,其面积约为1.25×109 hm2。广西、广东、贵州、湖南除北部地区、湖北、河南除北部地区、安徽、江苏除南部地区、福建西南部、浙江东北部、江西、内蒙古南部和东北部、黑龙江西部、吉林北部、西藏东南部、青海东部、甘肃北部、台湾除西南部等地区为芦苇的中度适生区,其面积约为1.88×109 hm2。黑龙江东南部、内蒙古中部、福建北部、浙江南部、安徽南部、广东南部、湖北西部、四川西部、西藏东部、新疆北部、青海中部、云南东北部、海南南部、吉林东北部、辽宁中部等地区为芦苇的低适生区,其面积约为1.08×109 hm2。
3 讨论与结论
该研究基于338条芦苇分布数据和6个环境变量,利用MaxEnt物种分布模型对芦苇在我国的潜在适生区进行预测。首先通过调参对MaxEnt模型进行优化,最终选择RM=1、FC=LQHP作为组合参数进行芦苇适生区预测,该参数组合下MaxEnt模型的AUC均值为0.909,表明MaxEnt模型预测芦苇适生区结果的可信度高。预测结果表明,芦苇的潜在分布区主要位于甘肃、山西、陕西、宁夏、河南、河北、北京、天津、山东、四川、重庆、上海、吉林、辽宁、黑龙江、湖北、湖南、江西、广西、广东、浙江、福建、江苏、安徽、贵州、内蒙古等地区。这与郭春秀等[36]研究表明的芦苇广泛分布在我国东北、华北、西北各省区以及长江流域的结果一致。
气候因子是制约物种分布的主要因素[37],MaxEnt模型预测的芦苇潜在分布区实质上反映了芦苇对该地区自然环境的选择。分析筛选得到的环境变量的贡献率、置换重要性和刀切法检验结果,得知年平均气温、最湿月降水量、等温性、最冷季度降水量、年平均气温变化范围是影响芦苇分布的主要环境变量。其中,年平均气温的贡献率和置换重要性远超其他环境变量之和,说明温度对芦苇分布的影响大于降水。沈红[38]发现温度是影响芦苇早发快长的重要因素之一,适宜的年平均气温促进芦苇的生长发育。雷玉红等[39]研究格尔木地区年平均气温、降水量对芦苇萌芽期的影响,发现暖冬促进芦苇萌发,而降水量对芦苇萌发的影响不显著。这些结论进一步验证了该研究的分析结果。
通过MaxEnt模型分别分析5个主要环境变量对芦苇分布的影响,由单变量响应曲线可知:当年平均气温为14~22℃、最湿月降水量为108~284 mm、等温性低于24、最冷季度降水量小于50 mm、年平均变化范围低于28.5℃时,芦苇的分布概率较高。
当前芦苇高度适生区集中在我国甘肃、山西、陕西、河南北部、河北、北京、天津、山东除南部、辽宁西部、宁夏、内蒙古西南部、湖南北部、四川东部、重庆西部、上海、江苏南部等地区。可综合考虑收割、运输以及加工成本,在高度适生区大规模种植芦苇,实现生态环境保护、资源利用和经济价值最大化,促进生物质资源的高值化利用。
参考文献:
[1] SRIVASTAVA J,KALRA S J S,NARAIAN R. Environmental perspectives of Phragmites australis(Cav.)Trin. Ex. Steudel[J]. Applied Water Science,2014,4(3):193-202.
[2] KETTENRING K M,WHIGHAM D F. Seed viability and seed dormancy of non-native Phragmites australis in suburbanized and forested watersheds of the Chesapeake Bay,USA[J]. Aquatic Botany,2009,91(3):199-204.
[3] BALDWIN A H,KETTENRING K M,Whigham D F. Seed banks of Phragmites australis-dominated brackish wetlands:relationships to seed viability,inundation,and land cover[J]. Aquatic Botany,2010,93(3):163-169.
[4] HOCKING P J,FINLAYSON C M,CHICK A J. The biology of Australian weeds. 12. Phragmites Australis(Cav.)Trin. ex Steud.[J]. Journal of the Australian Institute of Agricultural Science,1983,49:123-132.
[5] MARKS M,LAPIN B,RANDALL J. Phragmites australis(P. communis):threats,management and monitoring[J]. Natural Areas Journal,1994,14(4):285-294.
[6] SOTO-RÍOS P C,LEÓN-ROMERO M A,SUKHBAATAR O,et al. Biosorption of mercury by reed (Phragmites australis) as a potential clean water technology[J]. Water,Air,amp; Soil Pollution,2018,229(10):328.
[7] CHEN X X,ZHAI R,SHI K Q,et al. Mixing alkali pretreated and acid pretreated biomass for cellulosic ethanol production featuring reduced chemical use and decreased inhibitory effect[J]. Industrial Crops and Products,2018,124:719-725.
[8] 邢效功,孙衍宁,薛璟,等. 预水解对芦苇主要化学组分及化学制浆性能的影响[J]. 造纸科学与技术,2019,38(2):44-50.
[9] CAVALAGLIO G,GELOSIA M,INGLES D,et al. Response surface methodology for the optimization of cellulosic ethanol production from Phragmites australis through pre-saccharification and simultaneous saccharification and fermentation[J]. Industrial Crops and Products,2016,83:431-437.
[10] 杨倩. 采用低共熔溶剂制备芦苇纤维素纳米晶体[D]. 大连:大连工业大学,2021.
[11] CHEN M X,LI Q,ZHANG Y,et al. Xylo-oligosaccharides enriched yeast protein feed production from reed sawdust[J]. Bioresource Technology,2018,270:738-741.
[12] WANG Y Q,LU J,ZHOU S Y,et al. Bioconversion of cellulose and hemicellulose in reed sawdust to xylo-oligosaccharides and L-lactic acid[J]. Industrial Crops and Products,2022,187:115390.
[13] YANG D,ZHAO N N,TANG S X,et al. A hybrid strategy for efficient valorization of bulrush into furoic acid in water-ChCl-based deep eutectic solvent[J]. Industrial Crops and Products,2022,177:114434.
[14] ELITH J,LEATHWICK J R. Species distribution models:ecological explanation and prediction across space and time[J]. Annual Review of Ecology,Evolution,and Systematics,2009,40:677-697.
[15] SOBERÓN J,NAKAMURA M. Niches and distributional areas:concepts,methods,and assumptions[J]. Proceedings of the National Academy of Sciences of the United States of America,2009,106(supplement" 2):19644-19650.
[16] WANG Y J,XIE L Y,ZHOU X Y,et al. Prediction of the potentially suitable areas of Leonurus japonicus in China based on future climate change using the optimized MaxEnt model[J]. Ecology and Evolution,2023,13(10):e10597.
[17] HAASE C G,YANG A N,MCNYSET K M,et al. GARPTools:R software for data preparation and model evaluation of GARP models[J]. Ecography,2021,44(12):1790-1796.
[18] ANDERSEN D,BORZÉE A,JANG Y. Predicting global climatic suitability for the four most invasive anuran species using ecological niche factor analysis[J]. Global Ecology and Conservation,2021,25:e01433.
[19] SOUZA P G C,AIDOO O F,FARNEZI P K B,et al. Tamarixia radiata global distribution to current and future climate using the climate change experiment(CLIMEX)model[J]. Scientific Reports,2023,13:1823.
[20] BOOTH T H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important[J]. Austral Ecology,2018,43(8):852-860.
[21] PHILLIPS S J,ANDERSON R P,SCHAPIRE R E. Maximum entropy modeling of species geographic distributions[J]. Ecological Modelling,2006,190(3/4):231-259.
[22] PHILLIPS S J,DUDÍK M. Modeling of species distributions with Maxent:new extensions and a comprehensive evaluation[J]. Ecography,2008,31(2):161-175.
[23] SORBE F,GRÄNZIG T,FÖRSTER M. Evaluating sampling bias correction methods for invasive species distribution modeling in Maxent[J]. Ecological Informatics,2023,76:102124.
[24] WANG Y J,XIE L Y,ZHOU X Y,et al. Prediction of the potentially suitable areas of Leonurus japonicus in China based on future climate change using the optimized MaxEnt model[J]. Ecology and Evolution,2023,13(10):e10597.
[25] GAO R H,LIU L,ZHAO L J,et al. Potentially suitable geographical area for Monochamus alternatus under current and future climatic scenarios based on optimized MaxEnt model[J]. Insects,2023,14(2):182.
[26] KARUPPAIAH V,MARUTHADURAI R,DAS B,et al. Predicting the potential geographical distribution of onion Thrips,Thrips tabaci in India based on climate change projections using MaxEnt[J]. Scientific Reports,2023,13:7934.
[27] ZHANG Y,TANG J S,REN G,et al. Global potential distribution prediction of Xanthium italicum based on Maxent model[J]. Scientific Reports,2021,11:16545.
[28] 李世成,易自力,廖剑锋,等. 基于Maxent模型对尼泊尔芒适生区时空分布的预测[J]. 湖南农业大学学报(自然科学版),2020,46(2):176-183.
[29] MUSCARELLA R,GALANTE P J,SOLEY-GUARDIA M,et al. ENMeval:an R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models[J]. Methods in Ecology and Evolution,2014,5(11):1198-1205.
[30] RADOSAVLJEVIC A,ANDERSON R P. Making better Maxent models of species distributions: complexity,overfitting and evaluation[J]. Journal of Biogeography,2014,41(4):629-643.
[31] COBOS M E,PETERSON A T,BARVE N,et al. Kuenm:an R package for detailed development of ecological niche models using Maxent[J]. PeerJ,2019,7:e6281.
[32] PEBESMA E. Simple features for R:standardized support for spatial vector data[J]. The R Journal,2018,10(1):439-446.
[33] 廖剑锋,易自力,李世成,等. 基于Maxent模型的双药芒不同时期潜在分布研究[J]. 生态学报,2020,40(22):8297-8305.
[34] FIELDING A H,BELL J F. A review of methods for the assessment of prediction errors in conservation presence/absence models[J]. Environmental Conservation,1997,24(1):38-49.
[35] WILEY E O,MCNYSET K,PETERSON T,et al. Niche modeling perspective on geographic range predictions in the marine environment using a machine-learning algorithm[J]. Oceanography,2003,16(3):120-127.
[36] 郭春秀,李发明,张莹花,等. 河西走廊芦苇草地资源特征及其保护利用[J]. 草原与草坪,2012,32(4):93-96.
[37] HAMANN A,WANG T L. Potential effects of climate change on ecosystem and tree species distribution in British Columbia[J]. Ecology,2006,87(11):2773-2786.
[38] 沈红. 温度对芦苇生长发育的影响[J]. 现代农业,2013(12):41.
[39] 雷玉红,钟存,张翠花. 格尔木地区芦苇物候期与气候条件变化的响应[J]. 青海农林科技,2018(3):13-17.
(责任编辑:王婷)
猜你喜欢
海外文摘·文学版(2022年4期)2022-04-14 21:55:16
黄河之声(2021年19期)2021-02-24 03:25:24
公民与法治(2020年5期)2020-05-30 12:33:58
岁月(2018年2期)2018-02-28 20:40:58
中国诗歌(2017年12期)2017-11-15 04:04:05
农业与技术(2016年20期)2016-12-08 21:22:42
中国市场(2016年36期)2016-10-19 05:25:57
幼儿100(2016年28期)2016-02-28 21:26:18
天津农业科学(2015年12期)2015-12-03 00:07:53
河北渔业(2015年10期)2015-10-15 11:10:10
