
大花紫薇和‘丹红紫叶’紫薇杂交F1代不育细胞学分析
2024-12-31 00:00:00杨琦王湘莹唐丽王晓明乔中全陈艺曾慧杰李永欣
中南林业科技大学学报 2024年7期
关键词:细胞学
摘 要:【目的】大花紫薇和‘丹红紫叶’紫薇子代高度不育,为了解析其子代不育的机理。【方法】本试验以大花紫薇作母本和‘丹红紫叶’紫薇作父本的杂交F1代‘潇湘华秀’为材料,通过石蜡切片显微技术观察其雌雄蕊的生长发育过程及成熟花粉粒中的可溶性糖、淀粉、脂类物质的积累情况;并对3种紫薇花蕾不同生长发育阶段的可溶性糖含量、淀粉含量、游离脯氨酸含量进行分析测定。【结果】1)‘潇湘华秀’不育是雄性不育和雌性不育共同作用形成的,其花粉存在干瘪、畸形、大小不均等异常现象,绝大部分花粉不能萌发。2)与大花紫薇和‘丹红紫叶’紫薇相比,F1代在减数分裂时期发生异常,具体表现为减数分裂时,有部分染色体无法正常联会并排布到赤道板中央,染色体不均匀分离,发育成异常的二分体、带微核二分体、带微核四分体、五分体、六分体,且大部分花粉细胞核在二核期消解,成熟花粉粒大部分畸形,无内含物且不能萌发,可萌发的花粉粒为2n花粉粒。3)大孢子在四分体时期萎缩消解,后期形成有少量胚囊痕迹的空洞。4)F1代花蕾的可溶性糖、淀粉、游离脯氨酸含量都低于同等时期的亲本花蕾,且大部分花粉中未观测到脂类、可溶性糖和淀粉积淀。【结论】染色体行为异常导致异常四分体的出现和二核花粉粒时期细胞核的解体是花粉败育的主要原因;四分体时期大孢子萎缩消解是雌性不育的主要原因。
关键词:大花紫薇;丹红紫叶紫薇;杂交后代不育;花粉内含物;细胞学
中图分类号:S792.99 文献标志码:A 文章编号:1673-923X(2024)07-0140-13
基金项目:国家重点研发计划项目(2019YFD1001000);湖南省花卉苗木产业发展项目(HH202201)。
Cytological studies of F1 hybrid sterility between Lagerstroemia speciosa and Lagerstroemia indica ‘Ebony Embers’
YANG Qi1,2, WANG Xiangying2, TANG Li1, WANG Xiaoming2, QIAO Zhongquan2, CHEN Yi2, ZENG Huijie2, LI Yongxin2
(1. College of Forestry, Central South University of Forestry Technology, Changsha 410004, Hunan, China; 2. Hunan Academy of Forestry, Changsha 410004, Hunan, China)
Abstract:【Objective】The progeny of Lagerstroemia speciosa and Lagerstroemia indica ‘Ebony Embers’ are highly sterile, In order to analyze the reasons for the sterility of their offspring.【Method】In this experiment, L. indica ‘Xiaoxiang Huaxiu’ was used as the material which was the F1 hybrid of L. speciosa as the female parent and L. indica ‘Ebony Embers’ as the male parent, by paraffin section microscopy to observe the growth and development process of the stamens and pistils of male, and the mature pollen grains in the accumulation of soluble sugar, starch, lipid; and the contents of soluble sugar, starch, free proline in different growth stages were determined.【Result】1) The sterility of L. indica ‘Xiaoxiang Huaxiu’ was the result of combined male and female sterility. Pollen grains exhibit aberrant characteristics including shriveled appearance, deformities, and size variations, leading to the majority of pollen grains being unable to germinate. 2) Compared with L. speciosa and L. indica ‘Ebony Embers’, abnormalities occurred in the F1 generation during meiosis, specifically, some chromosomes were not arranged normally to the center of the equatorial plate during meiosis, and the chromosomes segregation was not uniform, developing into abnormal dyads, micronucleated dyads, micronucleated tetrads, pentads and sexads, and most pollen nuclei dissolve during the binucleate pollen grains stage. Mature pollen grains were mostly deformed without contents and couldnot sprout, the germinable pollen grains were 2n pollen grains. 3) The megaspore atrophies and dissolved in the tetrad stage, and formed a cavity with a few embryo sac traces in the later stage. 4) The contents of soluble sugar,starch and free proline in F1 generation’s flower bud were lower than those in the parent’s flower bud of the same period. Lipid, soluble sugar and starch accumulation was not observed in most pollen.【Conclusion】The abnormal behavior of chromosomes leads to the appearance of abnormal tetrads and the disintegration of nuclei in the period of binucleate pollen grains are the main reasons for pollen abortion. Atrophy and disintegration of megaspore during tetrad is the main cause of female sterility.
Keywords: Lagerstroemia speciosa, Lagerstroemia indica ‘Ebony Embers’, hybrid sterility, nutrients in pollen, cytology
紫薇Lagerstroemia indica与大花紫薇Lagerstroemia speciosa分类学上同属不同亚组,亲缘关系较远。研究发现紫薇与大花紫薇Lagerstroemia speciosa杂交后代存在高度不育现象[1-2],已有研究初步分析了大花紫薇和紫薇远缘杂交后代的不育性[2],但并没有从细胞学层面深入系统地观察杂交后代雌蕊和雄蕊的生长发育状况,没有阐明杂交后代的不育机理。‘丹红紫叶’紫薇Lagerstroemia indica ‘Ebony Embers’叶片紫黑色,花深红色,其与大花紫薇远缘杂交后代的花色比大花紫薇深,为深紫色,花径和叶片比‘丹红紫叶’紫薇大,抗寒性比大花紫薇强,能在湖南长沙安全越冬,且抗蚜虫和白粉病的能力较强,是性状优良的紫薇新种质。本研究以大花紫薇与‘丹红紫叶’紫薇的杂交后代为试验材料,通过显微观察杂交后代花蕾生长发育过程,从细胞学层面对杂交后代不育的原因进行探析,并结合生理指标测定,旨在探明大花紫薇和‘丹红紫叶’紫薇杂交后代不育的细胞学机理,为大花紫薇和紫薇远缘杂交育种提供理论依据。
1 材料与方法
1.1 试验材料
以湖南省林业科学院试验林场紫薇基地的大花紫薇(可育)、‘丹红紫叶’紫薇(可育)、‘丹红紫叶’紫薇作父本和大花紫薇作母本的杂交F1代‘潇湘华秀’Lagerstroemia indica ‘Xiaoxiang Huaxiu’(不育)为试材。
1.2 试验方法
1.2.1 花器官观察及杂交试验
2023年7月25日早晨7:00前采集3种紫薇未散粉的花朵进行花器官结构观察,使用英国皇家园林协会RHS植物比色卡对花色和叶色进行比色。用镊子剥离母本的花药并套袋标记,于9:00左右柱头可授性最强时用新鲜花粉授粉,套袋。每隔3日观察一次,7日后解开硫酸纸袋。设计5个杂交组合,分别是‘潇湘华秀’♀ב丹红紫叶’♂、‘潇湘华秀’♀×大花紫薇♂、大花紫薇♀ב潇湘华秀’♂、‘丹红紫叶’♀ב潇湘华秀’♂、‘潇湘华秀’自交。每组重复30次。
1.2.2 细胞学观察
2023年6月,采集‘潇湘华秀’花蕾,参照罗雪梦等[3-4]的切片方法切去花蕾顶端,剥离花瓣,置入FAA固定液:(V70%乙醇∶V甲醛∶V冰乙酸=18∶1∶1)抽气30 min浸泡24 h后转入70%乙醇4 ℃保存。常规石蜡切片,切片厚度10~18 μm。染色方法:铁苏木素染色、爱式苏木素-伊红对染,中性树胶封片,Olympus BX51光学显微镜观察并拍照。对照‘丹红紫叶’紫薇样本为本研究团队已制作完成的铁苏木素染色石蜡切片。
1.2.3 花粉扫描电镜观察
参照Zhou等[5]的试验方法取当日盛开的花药散出的花粉,2.5%戊二醛固定1.5 h,磷酸缓冲液冲洗2次,每次15 min,不同浓度乙醇逐级脱水,叔丁醇冷冻干燥,样品粘贴至扫描电镜铜板上镀金20 min,扫描电子显微镜拍照观察。对照‘丹红紫叶’为本研究团队已有的电镜扫描照片。
1.2.4 花粉萌发试验
2023年6月,参照张良英等[6-7]的试验方法,于7:00前采集未散粉的长短雄蕊置于通风阴凉干燥处,等待其脱水散粉后用毛笔轻刷入花粉培养基(蒸馏水1 000 mL、琼脂0.5 g、氯化钙20 mg、硼酸150 mg、蔗糖200 g)中,培养12 h后置于体式显微镜下取5个不同视野拍照观察并计算萌发率。使用亚历山大染色法、碘-碘化钾染色法对花粉染色并观察花粉活性,置于Olympus BX51光学显微镜下取5个不同视野拍照观察并计算花粉活性。
1.2.5 花粉内可溶性糖、脂质染色观察
2023年6月采集成熟期花蕾,使用常规石蜡切片切取成熟期花蕾进行苏丹黑B溶液染色,观察花粉脂肪积累情况;希夫试剂染色,观察可溶性糖积累情况。
1.2.6 花蕾营养物质含量测定
参照朱栗琼等[8]的方法确定紫薇花蕾的发育时期,随机采集‘潇湘华秀’各直径的花蕾的花药各15个,游标卡尺测量花蕾直径,结合花药压片、醋酸洋红染色确定不同直径花蕾发育时期:减数分裂时期(3.0~5.2 mm)、四分体时期(5.3~6.0 mm)、单核小孢子时期(6.1~6.5 mm)、单核靠边时期(6.5~6.8 mm)、二核花粉粒期(6.9~7.5 mm)、成熟期(>7.5 mm)。按各自直径采集3种紫薇5个时期的花蕾各20个,液氮速冻后-80 ℃冰箱保存,使用北京索莱宝公司可溶性糖含量、淀粉含量、游离脯氨酸含量检测试剂盒,严格按照说明书操作,进行3个指标含量的测定,各指标测定重复3次。
1.2.7 数据统计分析
图片处理使用Photoshop CS6软件;数据统计使用Excel 2016软件,数据分析制图使用GraphPad Prism软件。
2 结果与分析
2.1 花叶性状观察
‘丹红紫叶’紫薇花色深红(RHS 46A),花径平均3.20 cm,花瓣边缘褶皱明显,叶片中等大小,平均长6.32 cm、平均宽4.24 cm,叶片褐色(RHS 203B);大花紫薇花色紫(RHS N78C),花径平均:7.96 cm,花瓣边缘褶皱较平缓,叶片甚大,平均长19.82 cm,平均宽10.73 cm,叶片深绿(RHS NN137A);‘潇湘华秀’花色深紫(RHS N80A),花径4.21 cm,大小介于亲本之间,花瓣褶皱较为明显,叶片较大,平均长14.29 cm,平均宽7.18 cm,叶片紫绿色(RHS 147A)。由图1可知,大花紫薇、‘丹红紫叶’紫薇、‘潇湘华秀’均为两性花,三角形花萼6片、花瓣六枚、‘丹红紫叶’紫薇和‘潇湘华秀’具长雄蕊6枚、短雄蕊若干,3种紫薇花药均为丁字着药,大花紫薇花药的花粉囊扭曲至花丝花药连接处,呈现环状姿态;‘丹红紫叶’紫薇花药的花粉囊并未向花药花丝连接处延伸;‘潇湘华秀’花药的花粉囊介于亲本之间,有少部分向花药花丝连接处延伸,但并不成环状姿态。
2.2 杂交结实性
由表1可知,杂交授粉3日后,‘潇湘华秀’♀ב丹红紫叶’紫薇♂有30%的花留存,‘丹红紫叶’紫薇♀ב潇湘华秀’♂有3.33%的花留存,‘潇湘华秀’♀×大花紫薇♂组合有26.67%的花留存,而大花紫薇♀ב潇湘华秀’♂及‘潇湘华秀’自交的花全部脱落。杂交授粉7日后,五组杂交组合的花全部脱落,均未见子房发育为果实,且绝大多数杂交组合的子房未见膨大迹象,全部掉落。这表明‘潇湘华秀’与可育的‘丹红紫叶’和大花紫薇正交及反交均不结实,说明大花紫薇与‘丹红紫叶’紫薇的杂交后代‘潇湘华秀’同时存在雄性和雌性不育。


2.3 花粉扫描电镜观察
由图2可知,‘潇湘华秀’花粉形状极不规则,表面粗糙、干瘪,大小差异显著,部分花粉无明显萌发沟。大花紫薇和‘丹红紫叶’花粉为饱满圆润的花粉粒,3条萌发沟明显。
2.4 ‘潇湘华秀’及其亲本花粉活性观察

由图3可知,‘潇湘华秀’的花粉在体外萌发实验中基本不萌发,花粉萌发率仅为3%(图3d),花粉几乎没有活力;‘丹红紫叶’紫薇大多数花粉都可以萌发,花粉管生长状态良好,萌发率为68%(图3a);大花紫薇花粉萌发率也较高,萌发率为53%(图3c)。花粉碘-碘化钾试剂染色结果显示‘潇湘华秀’大部分花粉未着色,只有少部分直径较大的花粉被染成红褐色,说明大部分花粉内部无淀粉积累(图3e);亲本 ‘丹红紫叶’紫薇与大花紫薇的大部分花粉都被染成紫褐色,表明花粉内部有淀粉积累(图3b,h);亚历山大染色剂染色结果中‘潇湘华秀’的花粉大部分呈现蓝绿色,只有少数花粉呈紫红色,大部分花粉无活性(图3f),亲本只有少数花粉被染成蓝绿色,大部分花粉都呈现出紫红色,说明花粉活性较高(图3c,i)。这说明‘潇湘华秀’的花粉萌发率和花粉活性很低,显著低于亲本。
2.5 ‘潇湘华秀’小孢子的异常发育阶段
由图3可知‘潇湘华秀’雄配子体败育原因有绒毡层程序性死亡异常(图4a,b,d,e)、花药萎缩(图4c)和四分体异常(图4f)3种情况,其中二核期(图4b)和单核靠边期(图4e)可观察到明显无内含物积累和细胞核消解的透明状态花粉粒,主要的败育类型为四分体时期小孢子异常导致的败育。至花粉母细胞时期,不育系‘潇湘华秀’的小孢子体发育未出现异常,与亲本的发育相似性较高。减数分裂期后的四分体时期,出现异常五分体、六分体(图5a)和二分体(图5b),与可育系‘丹红紫叶’紫薇的正常四面体型四分体存在显著差异(图6a)。

紫薇形成四分体时胞质分裂方式为同时型,多分体的出现代表‘潇湘华秀’在减数分裂时期染色体的行为出现了异常,染色体的联会配对及减数分裂后染色体的分配有异。四分体时期后期,细胞板和细胞壁形成,随后绒毡层分泌出胼胝质酶将包围在四分体周围的胼胝质溶解,小孢子释放于花药的花粉囊中进一步生长发育,可见到不育系‘潇湘华秀’的单核中央期小孢子(图5c)为大小极不均匀且几近透明的泡状小孢子,细胞内部未见内含物与细胞核存在。而‘丹红紫叶’紫薇单核小孢子大小均匀,形状规整,内含物充足(图6b)。‘潇湘华秀’单核靠边期小孢子大小差异显著,逐渐出现四棱形的畸形小孢子(图5e);单核靠边期至二核花粉粒时期,液泡化的细胞质将细胞核挤至紧贴细胞壁处,细胞核将进行一次有丝分裂产生一个营养核和一个生殖核。‘丹红紫叶’紫薇此阶段可明显观察到位于细胞壁边缘的生殖细胞细胞核(图6c),特征明显(图6d),但‘潇湘华秀’大部分花粉粒细胞核都在此阶段发生消解,细胞内无明显的细胞核(图4e),唯一观察到的细胞核未消解的二核花粉粒的细胞核仍处于细胞中央,未被挤压到紧贴细胞壁处(图5f)。
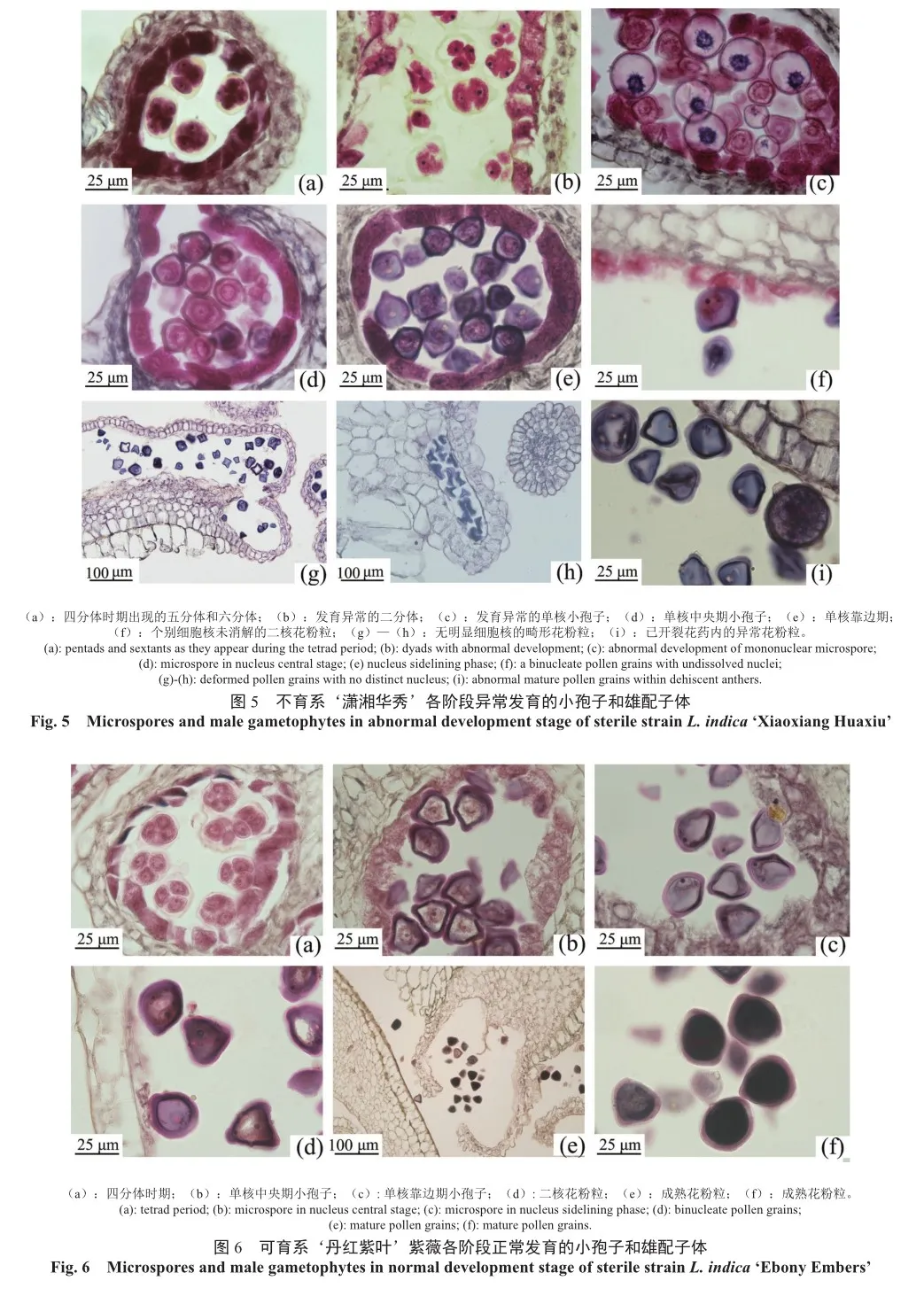
‘潇湘华秀’成熟期的花粉粒呈半透明状,无明显细胞质积累,花粉畸形,可观察到六面体花粉粒、半月形花粉粒、三棱花粉粒和四棱花粉粒(图5g),部分花粉囊内出现严重皱缩、排列紧密的干瘪花粉粒(图5h),但其花药可正常开裂,花粉可正常散出,且仅直径较大的花粉粒内含物较充足,其他花粉粒呈透明状(图5i)。同时期‘丹红紫叶’紫薇的花粉粒细胞质浓厚,花粉形状规则,大小均匀(图6e—f)。由此得出‘潇湘华秀’单核期小孢子的细胞核在有丝分裂生成营养核和生殖核过程中出现了异常。
2.6 ‘潇湘华秀’小孢子发育减数分裂时期、四分体时期异常情况观察
综合石蜡切片观察结果和花粉萌发实验结果,对‘潇湘华秀’小孢子母细胞的减数分裂时期和四分体时期进一步的研究,观测到7个重要的减数分裂阶段。
‘潇湘华秀’减数第一次分裂前期,细胞核消失,核仁解体,此时染色体进行同源染色体联会(图7a);减数第一次分裂中期,同源染色体着丝点对称排列于赤道板两侧(图7b,c),此时部分染色体并未被纺锤丝牵引至赤道板中央进行联会配对活动,明显落后于其他染色体(图7b);减数第一次分裂后期,同源染色体分离,非同源染色体自由组合后被纺锤丝牵引至细胞两侧(图7d)。
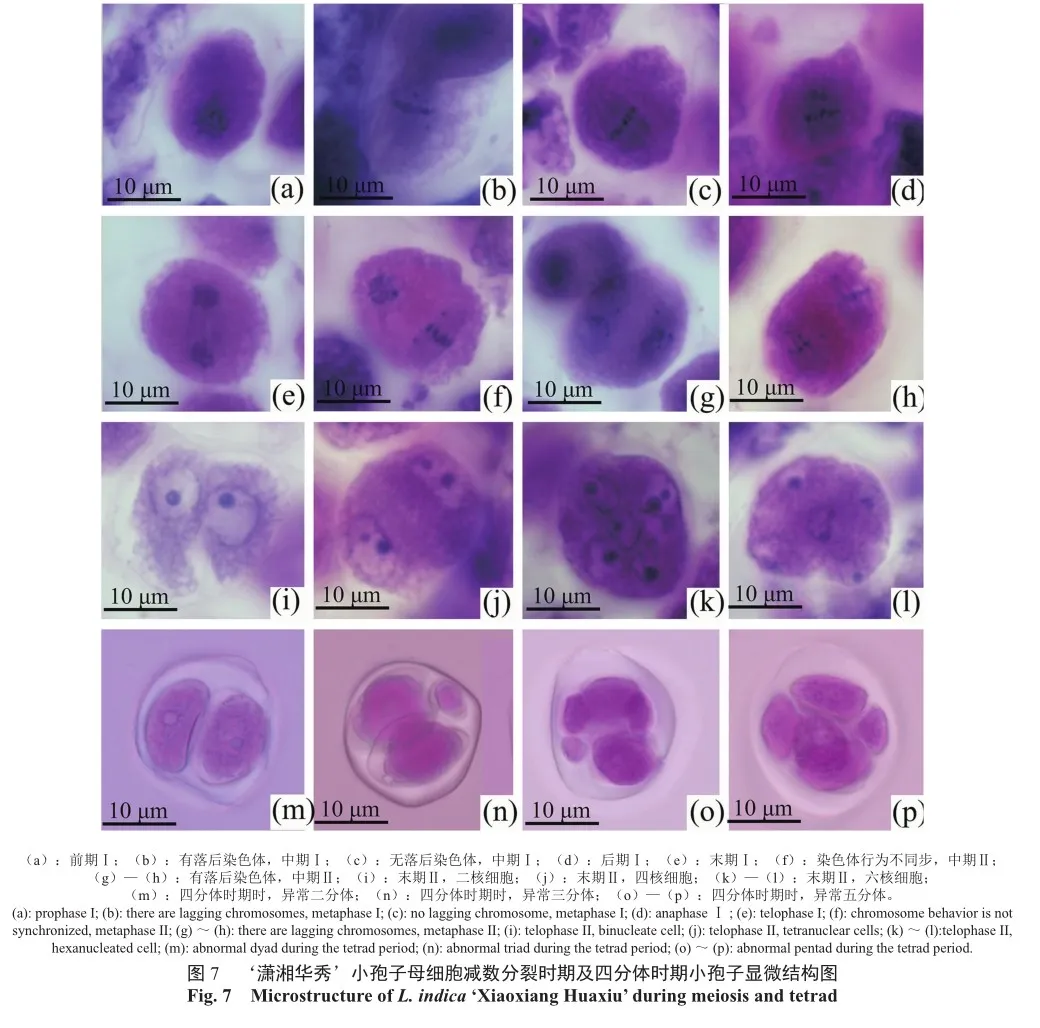
减数第二次分裂末期,核仁再次出现,形成两个细胞核(图7e)。此时部分细胞将停止减数进行减数分裂,并逐渐形成细胞壁和细胞板,最终演变为二分体(图7m);有落后的染色体的细胞将形成一个微核,并逐渐成为一个带微核的二分体(图7n);继续进行减数分裂的细胞在减数第二次分裂中期同样可以观测到大量行为异常的染色体(图7f,g),这些异常染色体的出现,是形成多分体的根本原因。减数第二次分裂末期,可以见到大量异常的细胞,即拥有两个细胞核(多数大小不均且有部分带有1个微核)、四个细胞核(多数带有1~2个微核)、五个细胞核(多数大小不均)、六个细胞核(多数大小不均)的细胞(图7i—l),并最终形成异常的多分体。
对‘潇湘华秀’的四分体阶段进行切片观察,发现只进行了减数第一次分裂的二分体数量虽然较多,但多数二分体的染色体在减数第一次分裂中期发生异常从而形成了带微核的二分体(图 8a,e,f),二分体和带微核的二分体在单核期形成了直径最大的单核小孢子(图8c,g),直径较小的小孢子来源于四分体、五分体、六分体(图8b),直径最小的小孢子是减数分裂时落后的染色体形成的微核携带少量细胞质形成的。成熟期的花粉粒大部分内含物空洞,未见细胞核,呈透明状,少数直径较大的花粉粒被染成蓝紫色(图8d,h),说明二分体形成的花粉粒有内含物积累。在‘潇湘华秀’的花粉体外萌发实验中,直径最大的花粉能萌发出花粉管(图8j),直径较小的花粉粒则不具有萌发能力。部分由带微核二分体转变的花粉粒在减数第一次分裂期丢失了部分染色体,虽然直径大但也不具萌发能力(图8i)。说明雄配子体败育的根本原因是染色体丢失。

2.7 ‘潇湘华秀’大孢子发育的显微结构
紫薇为单孢子蓼型胚囊,对各时期的子房进行了石蜡切片,发现‘潇湘华秀’大孢子二分体前的各阶段发育正常(图9a—b),胚囊母细胞在经过减数第一次分裂后形成二分体(图9c),大部分二分体进一步分裂为四分体时出现异常,四分体的细胞核出现消解现象,且大孢子细胞开始萎缩解体(图9d),少部分四分体可以正常发育为单核胚囊(图9e),但单核胚囊后,大孢子无法继续生长发育,萎缩为干瘪棒状细胞(图9f),随着花蕾逐渐生长,大孢子进一步消解(图9h),至花蕾开放时,只在珠心处观测到一个仍残留有部分絮状物的空洞(图9i)。说明减数分裂时期大孢子二分体不能进一步分裂为大孢子四分体,导致后期大孢子消解是‘潇湘华秀’雌性败育的主要原因。

2.8 ‘潇湘华秀’及其亲本花粉内脂类、可溶性糖积累情况检测
淀粉、可溶性糖和脂质是花粉内的重要内含物,其含量会影响花粉萌发和花粉管生长。苏丹黑B溶液可以将脂质着色,花粉壁富含脂类物质。由图10可知,3种紫薇的花粉粒都有不同程度的着色。‘潇湘华秀’亲本染色较深(图10a,c),‘潇湘华秀’的着色程度明显浅于亲本,呈透明状(图10b),说明‘潇湘华秀’花粉粒内部空洞,脂类物质积累很少,大花紫薇的和花粉粒也呈现出大小不均匀的状态(图10c),且较小的花粉既无内含物积累也无萌发能力,具体原因有待进一步研究。
高碘酸可将花粉内含的可溶性糖氧化为二醛基,进而与希夫试剂中的无色品红结合形成紫红色。‘丹红紫叶’紫薇、大花紫薇成熟花粉粒可溶性糖含量丰富,大部分花粉被希夫试剂染成深红色(图10d,f),而‘潇湘华秀’大部分花粉都呈现出淡红色,只有直径较大的一些花粉粒被希夫试剂染成深红色(图10e),说明‘潇湘华秀’大部分花粉粒内部可溶性糖的沉积很少。
2.9 ‘潇湘华秀’及亲本不同时期花蕾可溶性糖、淀粉和游离脯氨酸含量检测
由图11可知,3种紫薇花蕾发育过程中整体为上升趋势,在总体含量水平上,‘丹红紫叶’紫薇花蕾中可溶性糖含量在五个时期都高于大花紫薇,但在四分体时期和成熟期,三者花蕾中可溶性糖的含量并无显著差别。‘潇湘华秀’的可溶性糖在各阶段的含量都低于亲本。‘潇湘华秀’可溶性糖含量均值在成熟期与亲本差异显著,‘潇湘华秀’为16.54 mg·g-1,大花紫薇为21.20 mg·g-1,比‘潇湘华秀’高28.2%,‘丹红紫叶’为21.63 mg·g-1,比‘潇湘华秀’高30.8%。3种紫薇花蕾的可溶性糖含量在四分体时期到二核花粉粒时期上升较快,大花紫薇可溶性糖含量一直在增长、‘潇湘华秀’和‘丹红紫叶’紫薇可溶性糖含量在二核花粉粒时期后逐渐趋于平稳。
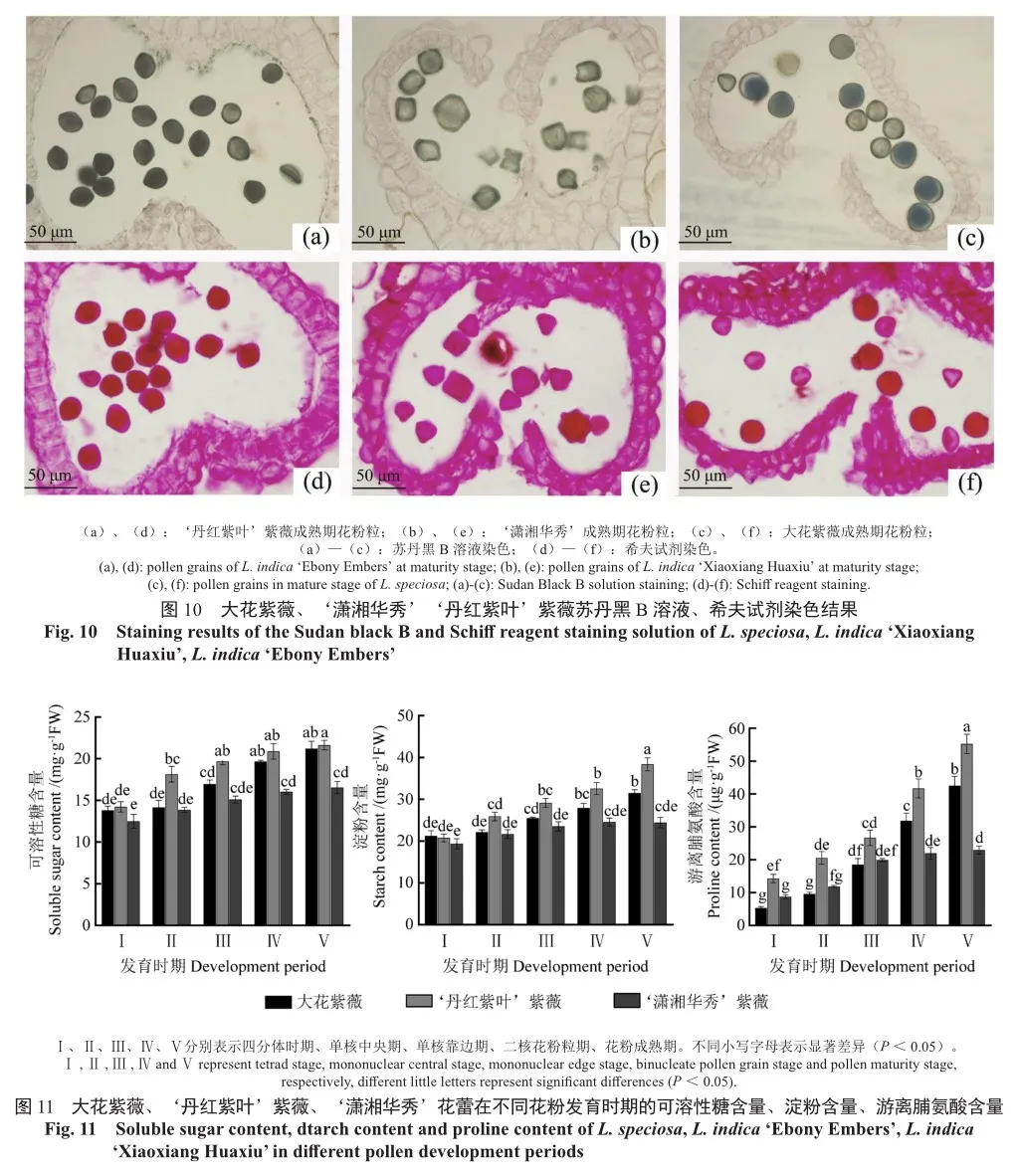
由图11可知,3种紫薇淀粉含量在花蕾发育过程中变化趋势与可溶性糖含量类似,呈上升趋势。但在成熟期‘丹红紫叶’紫薇的淀粉含量(38.38 mg·g-1)明显高于大花紫薇(31.50 mg·g-1)和不育的‘潇湘华秀’(24.45 mg·g-1),四分体时期至成熟期‘丹红紫叶’紫薇和大花紫薇的淀粉含量一直在递增。而‘潇湘华秀’的淀粉含量只在四分体时期(19.37 mg·g-1)至单核花粉粒时期(23.54 mg·g-1)有少量增长现象,单核靠边期后,淀粉含量无显著变化,且5个时期的淀粉含量差异并不显著。
由图11可知,3种紫薇脯氨酸含量在花蕾发育过程中变化趋势与可溶性糖和淀粉含量也类似,呈上升趋势,但大花紫薇、‘丹红紫叶’紫薇各时期游离脯氨酸含量差异显著,而‘潇湘华秀’花蕾中游离脯氨酸只在单核中央期(11.86 μg·g-1)至单核靠边期(19.91 μg·g-1)增长显著,其他时期增长无显著性差异。大花紫薇、‘丹红紫叶’紫薇、‘潇湘华秀’花蕾内游离脯氨酸含量差异显著,‘丹红紫叶’紫薇的游离脯氨酸含量在五个时期均显著高于其他两个紫薇品种。‘丹红紫叶’紫薇的游离脯氨酸含量在成熟期(55.25 μg·g-1)明显高于大花紫薇(42.53 μg·g-1)和‘潇湘华秀’(22.99 μg·g-1)。大花紫薇和‘潇湘华秀’在四分体时期至单核中央期的游离脯氨酸含量没有显著差异。大花紫薇二核期(31.85 μg·g-1)和成熟期(42.53 μg·g-1)花蕾内游离脯氨酸含量显著高于‘潇湘华秀’的二核期(21.98 μg·g-1)和成熟期(22.97 μg·g-1)。这进一步证明了‘潇湘华秀’花蕾内营养物质的积累低于亲本。
3 结论与讨论
3.1 结 论
研究结果表明,‘潇湘华秀’与可育的‘丹红紫叶’和大花紫薇正交及反交均不结实,大花紫薇与‘丹红紫叶’紫薇的杂交后代‘潇湘华秀’同时存在雄性和雌性不育。
研究结果表明,‘潇湘华秀’不育是雄性不育和雌性不育共同作用形成的。其花粉存在干瘪、畸形、大小不均等异常现象,绝大部分花粉不能萌发。雄性不育的根本原因是减数分裂时期染色体行为异常,部分染色体无法正常排列到赤道板上后逐渐形成微核,造成小孢子染色体丢失或染色体不均匀分配从而无法进行正常的生长发育。导致‘潇湘华秀’四分体时期形成异常二分体、带微核二分体、带微核四分体、五分体、六分体;使单核小孢子时期形成大小不均匀的畸形单核小孢子;二核花粉粒时期大部分花粉粒内含物空洞,花粉粒畸形,细胞核消解;成熟花粉粒中大部分花粉粒空瘪无萌发能力,只有直径较大的由二分体发育而来的2n花粉粒内含物丰富且能够萌发。
‘潇湘华秀’的雌性败育从二分体时期开始,二分体无法正常发育成四分体并萎缩凋亡,推测雌性不育与雄性不育的原因类似,都是减数分裂异常所导致的不育,由于染色体分裂不均匀等异常原因,致使二分体阶段后的大孢子细胞无法正常进行生长发育等生命活动,从而导致雄性不育。
‘潇湘华秀’花粉中无脂质、可溶性糖、淀粉的积累,花蕾中可溶性糖、淀粉、游离脯氨酸的含量显著低于亲本‘丹红紫叶’和大花紫薇,说明‘潇湘华秀’花蕾内营养物质积累较少,这可能与其不育性有关。
3.2 讨 论
植物不育类型分为雄性不育、雌性不育、雌雄性皆不育。目前国内外对植物雄性不育系研究较多,在水稻、柑橘[7]、油茶[9]、烟草[10]、杨树[11]、白菜[12]等重要经济植物种中都有报道。雄性不育的发生一般是减数分裂时期的染色体行为[13]、纺锤体行为[14]、绒毡层程序性死亡[15]、胼胝质沉积、花药开裂等生命活动发生异常引起。‘潇湘华秀’的雄性不育是以染色体行为异常为主,绒毡层程序性死亡异常和花药萎缩协同导致的雄性不育。植物雌性不育一般是减数分裂异常[16]、雌蕊柱头或花柱退化或发育异常[17]、胚珠发育异常[18]造成的,但是大部分的雌性不育现象,都是减数分裂异常无法分化出正常的功能性大孢子导致,这与本研究结果一致。
在植物界中,远缘杂交通常会出现生殖隔离,表现为父母本杂交不亲和的合子前生殖障碍[19]和杂交受精卵不能正常发育为单独个体的合子后生殖障碍[20],有学者对紫薇属的种间杂交的生殖隔离与不育现象进行了相应的报道,发现川黔紫薇和紫薇的正反交组合会由于花柱中胼胝质的大量积累导致花粉管无法正常伸长到子房,合子前生殖障碍让二者无法产生受精卵[21];在有关不育紫薇‘湘韵’的细胞学研究之中,指出了不育紫薇的花粉存在内含物缺失,花粉粒空瘪等现象,并且发现不育花蕾的花丝细胞存在严重变形现象[5,22]。
‘潇湘华秀’高度不育,但其自然状态下产生的可萌发的2n花粉粒可作为紫薇育种的重要材料。2n雄配子是自然界中三倍体植物的重要来源,一般认为高温等外界条件的影响可以促使小孢子减数分裂时期发生异常,从而诱导植物产生2n雄配子。‘潇湘华秀’中存在只进行一次减数分裂的二分体,和只有一半染色体排列在赤道板上在进行着丝点的分裂的减数第二次分裂中期的细胞,这与高温诱导油茶产生2n花粉粒的相关研究之中诱导2n雄配子时出现的染色体异常分裂行为相似[23],至于二者2n雄配子出现的内在机理是否一致,还需进一步研究。
植物花粉内含物中的可溶性糖和淀粉是与花粉管萌发相关的重要营养物质,游离脯氨酸广泛存在于植物体内,在植物各项生命活动中承担重要功能,尤其是在植物生长旺盛的组织内和生殖器官中含量较高,游离脯氨酸在花粉发育及花粉萌发等生命进程中作用较大[24],它是合成相关蛋白和酶的重要原料[25],同时也是花粉管萌发和生长的重要氨源,因此游离脯氨酸含量也在一定程度上与植物可育性存在关联。
大花紫薇[8]和‘丹红紫叶’紫薇[23]雌雄配子体发育已有相关文献发表,但大花紫薇和‘丹红紫叶’紫薇杂交后代不育机理尚是空白,至今未见到从细胞学层面解析大花紫薇和紫薇杂交后代不育机理的相关研究报道。本研究对大花紫薇和‘丹红紫叶’紫薇杂交后代‘潇湘华秀’的雌雄配子体发育过程进行的细胞学观察与相关生理实验,可为紫薇不育新品种创制提供重要的理论依据,但本研究只观测到了减数分裂终变期及以后的染色体行为,未对细线期、偶线期、粗线期及双线期的染色体行为进行观察,无法对上述时期的染色体行为进行判断,下一步应该对减数分裂各相关时期的染色体进行详细观察,找到染色体落后等异常情况发生的原因,并从分子层面对‘潇湘华秀’紫薇的不育机理进行研究。
参考文献:
[1] 焦垚.大花紫薇与紫薇杂交后代重要观赏性状遗传分析[D].北京:北京林业大学,2017. JIAO Y. Phenotypic and genetic analysis of ornamental traits in hybrids of Lagerstroemia indica and L. speciosa[D]. Beijing: Beijing Forestry University,2017.
[2] 胡杏,鞠易倩,叶远俊,等.紫薇×大花紫薇不同育性株系花粉生活力及柱头可授性对比分析[J].河南农业大学学报, 2014,48(2):145-149. HU X, JU Y Q, YE Y J, et al. Comparative investigation on the pollen viability and stigma receptivity of different hybrids between Lagerstroemia indica and L. speciosa[J]. Journal of Henan Agricultural University,2014,48(2):145-149.
[3] 罗雪梦,王晓明,曾慧杰,等.紫叶紫薇花芽分化及雌雄配子体发育的形态学研究[J].中南林业科技大学学报,2022,42(2): 129-138. LUO X M, WANG X M, ZENG H J, et al. Morphological study on floral bud differentiation and the development of female and male gametophytes in purple-leaf Lagerstroemia indica[J]. Journal of Central South University of Forestry Technology,2022,42(2): 129-138.
[4] 韦开萍,杨茜,和润喜,等.腾冲红花油茶花芽分化及雌配子体发育的形态学观察[J].经济林研究,2023,41(3):244-251. WEI K P, YANG X, HE R X, et al. Morphological study on floral bud differentiation and the development of female gametophytes in Camellia reticulate f. simplex Sealy[J]. Non-wood Forest Research,2023,41(3):244-251.
[5] ZHOU W, WANG X M, CHEN J H, et al. Abortion categories and characteristics of acarpous crape myrtle floral organs[J]. Journal of the American Society for Horticultural Science,2019,144(6): 387-393.
[6] 张良英,刘林,翟秋喜,等.不同软枣猕猴桃雄株花粉离体培养条件及花粉性状的比较[J].经济林研究,2023,41(1): 255-264. ZHANG L Y, LIU L, ZHAI Q X, et al. Comparison of pollen culture in vitro conditions and pollen traits of different male plants of Actinidia arguta[J]. Non-wood Forest Research,2023,41(1): 255-264.
[7] 叶丽霞.多组学解析‘华农’无籽椪柑花粉败育机制[D].武汉:华中农业大学,2021. YE L X. Multiomics analysis of the mechanism of pollen abortion in ‘Huanong’ seedless ponkan[D]. Wuhan: Huazhong Agricultural University,2021.
[8] 朱栗琼,袁娟,韦立迪,等.大花紫薇大小孢子的发生及雌雄配子体的发育[J].植物研究,2018,38(2):182-188. ZHU L Q, YUAN J, WEI L D, et al. Sporogenesis and gametogenesis of Lagerstroemia speciosa[J]. Bulletin of Botanical Research,2018,38(2):182-188.
[9] 蒋会兵,杨盛美,刘玉飞,等.厚轴茶雄性不育株花药败育的生物学特性和细胞学研究[J].作物学报,2020,46(7): 1076-1086. JIANG H B, YANG S M, LIU Y F, et al. Biological characteristics and cytological studies on anther abortion of male sterile Camellia crassocolumna[J]. Acta Agronomica Sinica,2020,46(7): 1076-1086.
[10] 郑云,崔芳芳,郑九洲,等.烟草雄性不育突变转录组相关差异表达基因分析[J].西南农业学报,2022,35(8):1733-1741. ZHENG Y, CUI F F, ZHENG J Z, et al. Analysis of differentially expressed genes related to tobacco male sterility mutant transcriptome[J]. Southwest China Journal of Agricultural Sciences, 2022,35(8):1733-1741.
[11] 张媛.‘抱头毛白杨’花粉败育的细胞学机理研究[D].北京:北京林业大学,2019. ZHANG Y. Cytological mechanism of pollen abortion in Populus tomentosa clone ‘baotou’[D]. Beijing: Beijing Forestry University, 2019.
[12] 杨琳.受体激酶SRK与FERONIA调控大白菜远缘杂交生殖隔离的分子机制[D].泰安:山东农业大学,2023. YANG L. SRK and FERONIA receptor kinases control distant hybridization reproductive barriers in the brassicaceae[D]. Tai’an: Shandong Agricultural University,2023.
[13] KUMARI K, SAGGOO M I. Meiotic abnormalities in some members of the poaceae collected in district kinnaur, himachal pradesh, India[J]. Chromosome Botany,2016,11(1):14-20.
[14] ZHANG Z H, KANG X Y. Cytological characteristics of numerically unreduced pollen production in Populus tomentosa[J]. Euphytica,2010,173(2):151-159.
[15] KIRAN A, KUMAR S, NAYYAR H, et al. Low temperatureinduced aberrations in male and female reproductive organ development cause flower abortion in Chickpea[J]. Plant, Cell Environment,2019,42(7):2075-2089.
[16] SZURMAN-ZUBRZYCKA M, BARAN B, STOLAREKJANUSZKIEWICZ M, et al. The dmc1 mutant allows an insight into the DNA double-strand break repair during meiosis in barley(Hordeum vulgare)[J]. Frontiers in Plant Science,2019,10:761.
[17] 王成.掌叶木雄花雌蕊败育的形态学和生理生化特性研究[D].南宁:广西大学,2015. WANG C. Morphological and physicochemical characteristics of female sterility of male flowers of Handeliodendron bodinievri[D]. Nanning: Guangxi University,2015.
[18] LILLECRAPP A M, WALLWORK M A, SEDGLEY M. Female and male sterility cause low fruit set in a clone of the ‘Trevatt’variety of apricot (Prunus armeniaca)[J]. Scientia Horticulturae, 1999,82:255-263.
[19] OUYANG Y D, ZHANG Q F. Understanding reproductive isolation based on the rice model[J]. Annual Review of Plant Biology,2013,64:111-135.
[20] MCCLURE B A, FRANKLIN-TONG V. Gametophytic self- incompatibility: understanding the cellular mechanisms involved in “self” pollen tube inhibition[J]. Planta,2006,224:233-245.
[21] 王甜.紫薇和川黔紫薇远缘杂交亲和性及种子萌发特性研究[D].长沙:中南林业科技大学,2022. WANG T. Study on compatibility and seed germination characteristics of the distant hybridization between Lagerstroemia indica and Lagerstroemia excelsa[D]. Changsha: Central South University of Forestry Technology,2022.
[22] 许欢.结实与不结实紫薇生物学特性比较研究[D].长沙:中南林业科技大学,2015. XU H. Biological characteristic comparison research between fructiferous and acarpous Lagerstroemia indica[D]. Changsha: Central South University of Forestry Technology,2015.
[23] 张晓瑜.高温诱导油茶花粉母细胞减数分裂异常行为研究[D].长沙:中南林业科技大学,2022. ZHANG X Y. Abnormal behavior of pollen mother cells induced by high temperature in Camellia oleifera[D]. Changsha: Central South University of Forestry Technology,2022.
[24] 陈伟,付薇,吴佳海,等.高羊茅雄性不育小孢子败育的细胞学及生理生化特性[J]. 植物生理学报,2017,53(9):1761-1771. CHEN W, FU W, WU J H, et al. Cytology, physiological and biochemical characterization in microspore abortion of male sterile plant in Festuca arundinacea[J]. Plant Physiology Journal, 2017,53(9):1761-1771.
[25] 郭艳玲.梨雄性不育细胞学和生理机理研究[D].南京:南京农业大学,2006. GUO Y L. Study on the mechanism in cytology and physiology of the male-sterility in pear[D]. Nanjing: Nanjing Agricultural University,2006.
[本文编校:吴 毅]
猜你喜欢
世界最新医学信息文摘(2021年12期)2021-06-09 08:36:28
国际呼吸杂志(2019年2期)2019-02-14 06:11:26
中外医疗(2016年15期)2016-12-01 04:25:40
中国卫生标准管理(2015年25期)2016-01-14 09:29:31
中国麻业科学(2015年5期)2015-12-28 06:22:07
转化医学电子杂志(2015年4期)2015-12-27 12:17:02
肿瘤预防与治疗(2015年1期)2015-09-26 07:26:22
中国卫生标准管理(2015年25期)2015-01-27 01:05:38
当代医学(2014年35期)2014-08-07 02:54:32
湖北科技学院学报(医学版)(2014年2期)2014-02-28 19:42:33
