
桂西南岩溶区植物叶性状的系统发育信号及其关联分析
2024-12-31 00:00:00庞世龙欧芷阳申文辉叶斯进黄宝珍凌福诚陆国导
中南林业科技大学学报 2024年7期
关键词:岩溶
摘 要:【目的】探究岩溶植物叶性状在不同生活型水平上的差异以及系统发育保守性,有助于深入理解植物对异质性生境的适应机制,进而为岩溶植被的保护与恢复提供科学依据。【方法】以桂西南岩溶区20种常见阔叶木本植物为对象,采用K值法检验了叶性状的系统发育信号,并运用系统发育独立比较(PIC)和标准化主轴估计(SMA)的方法分析了叶性状间的关联适应性。【结果】1)研究区植物群落10个叶性状均未表现出较强的系统发育保守性,系统发育结构与性状结构并不完全一致。2)叶面积(LA)与叶长(LL)、叶宽(LW)以及叶组织密度(LTD)与LL、叶体积(LV)间均呈异速生长关系,而LL与LW、LA与叶干质量(LDW)间均呈等速生长关系;其中,LTD与LL的生长关系在常绿植物组发生了改变,由异速生长关系转变为等速生长关系;其他性状组合间的生长关系并未因生活型的不同而引起差异,表明常绿植物倾向于限制LL以适应环境的变化;3)LA、LL和LW以及LTD与LL、LV在常绿和落叶植物组间的斜率均沿共同主轴方向显著漂移(P<0.05),表明常绿和落叶植物对环境的适应策略发生了位移,常绿植物倾向于选择小叶、组织密度大的资源保守型生长策略,而落叶植物则倾向于选择大叶、组织密度小的资源获取型生长策略。【结论】生态位分化在很大程度上是促进亚热带阔叶林群落物种共存和生物多样性维持的重要机制。
关键词:叶性状;标准化主轴估计;系统发育独立比较;异速生长;系统发育信号;岩溶
中图分类号:S792.99 文献标志码:A 文章编号:1673-923X(2024)07-0001-09
基金项目:广西林业科技推广示范项目(桂林科研〔2022〕24号);中央财政林业科技推广示范项目(〔2022〕TG23号);广西科技基地和人才专项(桂科22-035-130-02)。
Phylogenetic signals of plant leaf traits and their correlation analysis in the karst region of southwestern Guangxi
PANG Shilong1, OU Zhiyang1, SHEN Wenhui1, YE Sijin2, HUANG Baozhen3, LING Fucheng3, LU Guodao1
(1.a. Central South Key Laboratory of Fast-growing Tree Cultivation of National Forestry and Grassland Adiministration; b. Guangxi Key Laboratory of Superior Timber Trees Cultivation, Guangxi Forestry Research Institute, Nanning 530002, Guangxi, China; 2. Nanning Qingxiushan Tourism Development Co. Ltd., Nanning 530022, Guangxi, China; 3. Pingguo Forestry Bureau, Pingguo 531499, Guangxi, China)
Abstract:【Objective】Exploring the differences in leaf traits and phylogenetic conservatism of karst plants of different life types can help to deeply understand the adaptive mechanisms of plants to highly heterogeneous habitat, and provide scientific basis for the conservation and restoration of karst vegetation.【Method】Twenty common broad-leaved woody plants in a karst region of southwestern Guangxi were studied. The Phylogenetic signals and correlations among leaf traits were tested by using Blomber’s K value, phylogenetic independent contrast (PIC) and standardized major axis estimation (SMA).【Result】1) None of the studied ten leaf traits showed strong phylogenetic conservatism, and the phylogenetic structure was not fully consistent with the trait structure. 2) Overall, there were allometric growth relationship between leaf area (LA) and leaf length (LL) and width (LW), and between leaf tissue density (LTD) and leaf length (LL) and volume (LV). In contrast, the isokinetic growth relationship were found between leaf length and leaf width, and between leaf area and leaf mass. Except that the growth relationship between LTD and LL shifted from allometric to isokineticin the deciduous plant, the growth relationship between other trait combinations didn’t differ between life forms, which indicated that ever-green plant tends to limit LL growth for adapting the environment changes. 3) The slopes between LA and LL and LW, as well as between LTD and LL and LV for both of ever-green and deciduous plant significantly along the direction of the common principal axis (P<0.05), suggesting a change of adaptive strategies of evergreen and deciduous plants to their environment. The ever-green plant tended to choose the resource-conserving growth strategy with smaller leaf area and higher leaf tissue density, whereas the deciduous plant tends to choose the resource-acquiring growth strategy with larger leaf area and lower leaf tissue density.【Conclusion】Ecological niche differentiation is largely an important mechanism to promote species coexistence and biodiversity maintenance in broadleaf forest communities in subtropical regions.
Keywords: leaf trait; standardized major axis estimation; phylogenetic independent contrast; allometric growth; phylogenetic signal; karst
植物功能性状是指植物对外界环境长期适应和进化后所表现出的可量度特征,是联系植物、环境和生态系统功能的桥梁,能够客观地表达植物对环境的适应性[1],同时也反映了物种所在生态系统的功能特征[2]。近20年来,植物功能性状的研究得到了广泛关注和迅速发展,已成为探索各类生态学前沿领域及热点问题的重要手段[3]。在诸多植物功能性状中,叶片性状对气候变化非常敏感且可塑性较大,直观反映了植物对气候变化的响应与适应,直接影响植物的基本行为和功能,体现了植物为获得最大碳收入和最小水分消耗所采取的生存适应策略[4],具有重要的生态学和生物进化意义。
中国西南碳酸盐岩出露面积53.26万km2,是全球三大岩溶集中分布区中连片面积最大、岩溶发育最强烈的地区。其特殊的地质背景和二元水文结构导致土层浅薄,土被不连续,岩体裂隙发育密集且连通性好、透水性强,地表水大量渗漏、地下水深埋,水文过程异常迅速,岩溶干旱频发。水分亏缺是制约该区植被生长、分布及恢复重建的关键限制因子[5]。岩溶植被在长期的适应性进化过程中具有强烈的选择性(喜钙性、石生性和旱生性),且顺向演替难、逆向演替易,其适应机理与其他类型区显著不同。与非岩溶植物相比,岩溶植物具有较高的叶磷含量、光合能力、水分利用效率和较小的气孔安全边界,且叶性状之间的关联性更高[5-6]。植物功能性状的变异受进化历史和环境的共同驱动[7]。植物系统发育背景是植物功能性状变异的主要来源之一,甚至超过环境梯度的影响[8-9]。但过去在进行性状种间差异或相关分析时,较少考虑物种间不同进化历史的影响(即假设物种间相互独立),这可能错误估计了性状间的相互关系,从而导致接受了不存在的进化权衡。本研究以桂西南岩溶山地20种常见阔叶木本植物(常绿和落叶植物各10种)为对象,采用K值法检验了叶性状的系统发育信号,并运用系统发育独立比较和标准化主轴估计的方法分析了叶性状间的关联适应性,试图探讨以下两个问题:1)植物叶性状是否具有显著的系统发育信号?即叶性状是否受到系统发育历史的显著影响?2)植物叶性状在不同生活型物种间是否具有一致的生长关系?研究结果将有助于更好地理解亚热带常绿落叶阔叶混交林群落的构建与维持机制,为西南岩溶区植被的保护与恢复提供科学依据。
1 材料与方法
1.1 研究区概况
研究地位于广西平果市太平镇旺里村旺上屯(23°34′32″N,107°29′52″E),属南亚热带季风气候区,光照充足,热量丰富,雨热同期。年平均气温19~21.5 ℃,降水多集中于5—9月,年降水量1 400~1 550 mm,年平均蒸发量1 572 mm,相对湿度81%,无霜期345 d以上。研究区域为典型的峰丛洼地分布区,土壤以深色或棕色石灰土为主,土层浅薄,岩石裸露率高。由于人类活动频繁,原生植被已退化为藤刺灌丛和稀疏灌草丛。2002年开始退耕还林,后经多次补植补播和除草施肥等干预措施的恢复,形成了独特的常绿落叶阔叶混交林群落。
1.2 样品采集
于植被生长盛期(8月),在研究地内选取20种常见阔叶木本植物(表1)。每个物种选择5株长势良好的个体,大小参考群落平均胸径。利用高枝剪在每棵样株冠层的外缘,随机采集12片不同方位、完全展开且无病虫害的成熟叶片。将采集的叶片用湿润的滤纸包裹,迅速放入自封袋储藏于便携式冷藏箱中,带回实验室分析。
1.3 性状测定
洗净擦干叶片并编号标记后,用电子天平(精度0.001 g)称其鲜质量(LFW,g);用叶面积仪(CI-203,CID,Inc,USA)测量叶长(LL,cm)、叶宽(LW,cm)和叶面积(LA,cm2);用数显测厚规(精度0.001 mm)沿叶片主脉方向均匀选取3个点,测量主脉一侧约0.25 cm处的厚度,取其均值即叶片厚度(LT,cm);然后将叶片放入 105 ℃烘箱内杀青30 min,随后80 ℃烘干至恒质量,冷却后称量叶干质量(LDW,g)。叶含水率(LWC)、叶体积(LV)、比叶面积(SLA)、叶干物质含量(LDMC)和叶组织密度(LTD)按照参考文献[10]的方法进行计算。

1.4 数据分析
1.4.1 系统发育信号检验
采用Blomberg?s K值检验性状的系统发育信号强度。利用R语言的picante程序包对性状系统发育信号K值及其显著性P值进行估计。若K>1,表明性状具有比按布朗运动模型(Brownian motion)进化更强的系统发育信号,性状表现出系统发育保守性,系统发育与性状结构具有一致性;若K<1,表明性状的系统发育信号较弱,物种趋同进化,系统发育结构与性状结构并不完全一致;若K接近于0,则表明性状的系统发育信号趋向于随机,性状结构与系统发育结构无关。
1.4.2 系统发育独立比较
为确定不同性状间的关系是否随进化分歧而改变,采用系统发育独立比较(PIC)的方法进行分析。首先利用物种的科属信息,应用R语言的V.PhyloMaker程序包生成系统发育树,用ape包计算性状的系统发育独立比较值;然后采用Pearson相关分析检验各性状间的进化关联。
1.4.3 叶性状间的关系
对于植物叶性状以物种算术平均值进行分析。叶性状间的关系采用经典异速方程Y=βXα来描述,各变量经对数(以10为底)转换成线性关系:logY=logβ+αlogX;式中,X和Y分别代表所考察的不同性状值,logβ为性状关系的截距,α为斜率。当|α|=1时,表示拟合性状间为等速生长关系,当|α|显著偏离1时,表示拟合性状间为异速生长关系。参数估计及检验采用标准化主轴估计(SMA)的方法,由软件(S)MATR Version 2.0计算完成。
2 结果与分析
2.1 叶性状的系统发育信号
桂西南岩溶区植物群落10个叶性状的Blomberg’s K值均小于1(表2);其中,LWC、LDMC和LTD表现出显著的系统发育信号(P<0.05),而其余7个叶性状均未检测到显著的系统发育信号。故该区植物叶性状进化存在一定的非保守性,且系统发育对不同性状的影响程度不同,除LWC、LDMC和LTD相对影响程度较大外,总体影响较弱。

2.2 叶性状间的关联进化
基于性状物种均值及其系统发育独立比较值的相关分析表明,部分性状间表现为趋同进化或趋异进化,且程度不一(图1)。无论是否考虑系统发育的影响,LW、LL、LA、LDM和LV以及SLA与LW、LL、LA间均存在显著正相关关系(P<0.05),物种性状间表现为趋同进化;而LTD与LW、LL、LA、LDM、LV、SLA以及LWC与LDMC间均存在显著负相关关系(P<0.05),表现为趋异进化。在剔除系统发育的影响之后,LT与LW、LL、LA、LDM、LV、LTD和SLA与LDMC、LWC以及LTD与LDMC、LWC之间的相关关系均不显著。

2.3 叶性状间的关系
2.3.1 叶面积与叶长及叶宽的关系
LA与LL、LW在常绿和落叶植物组间的斜率均无显著差异(表3,图2A—B),其共同斜率分别为1.660和1.890,均显著大于1(P<0.001),表明LA与LL、LW之间存在显著的异速生长关系,且LA的扩展速率显著大于LL和LW的增长速率。两组关系的截距在常绿和落叶植物组间亦无显著差异,但其斜率均沿共同主轴显著向上漂移(P<0.05)。
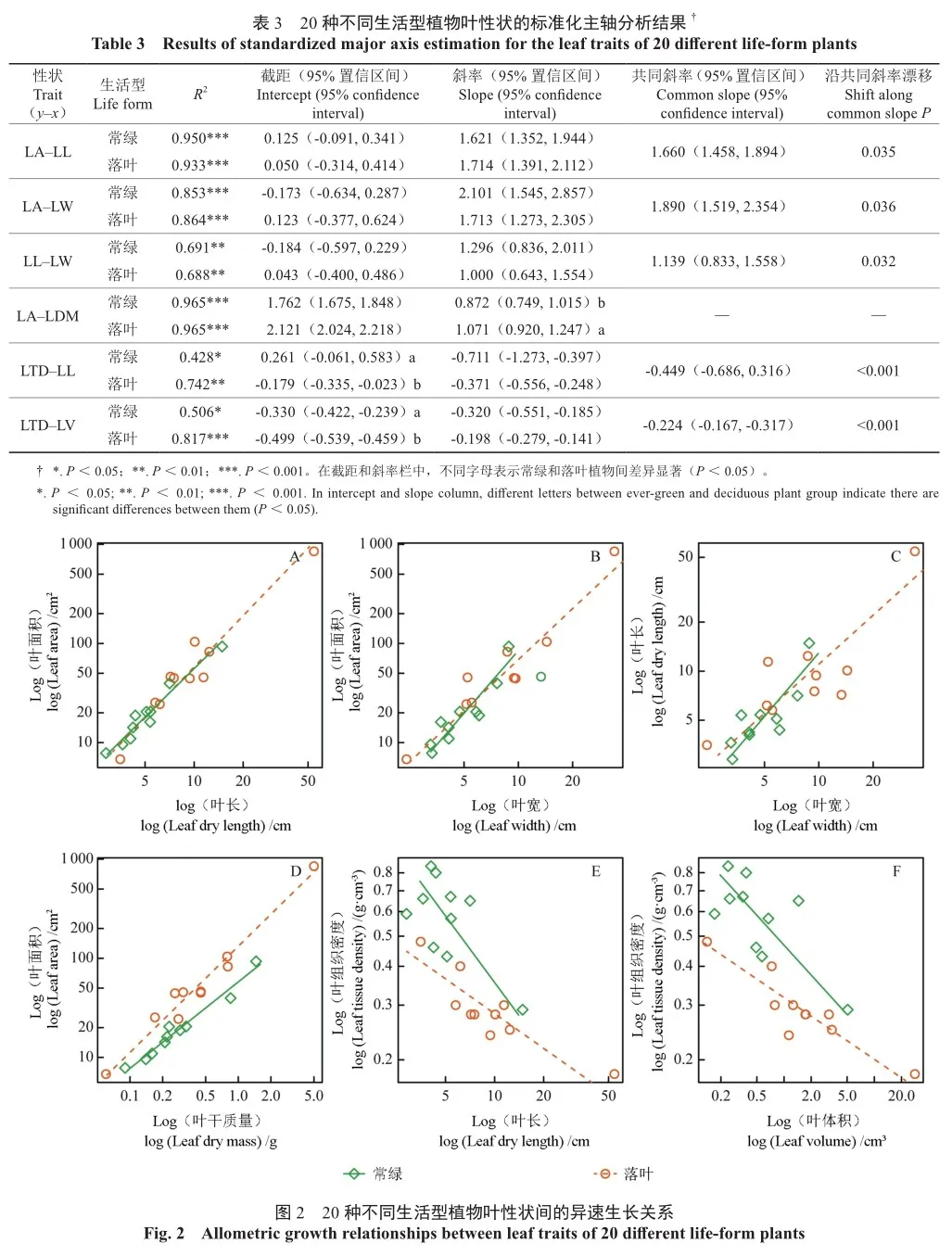
2.3.2 叶长与叶宽的关系
LL与LW的斜率在常绿和落叶植物组间差异不显著(表3,图2C),其共同斜率(1.139)与1无显著差异的等速生长关系,即LL随LW等速生长。他们的截距在常绿和落叶植物组间无显著差异,但其斜率沿共同主轴显著向上漂移(P<0.05)。
2.3.3 叶面积与叶干质量的关系
落叶植物组LA与LDW的斜率(1.071)显著高于常绿植物组(0.872)(P<0.05,表3,图2D),表明在LDW增加相同的情况下,落叶植物的LA比常绿植物增长更快。他们的斜率与1均无显著差异,表明常绿和落叶植物的LA扩展速率与LDW积累速率大致相等,二者呈等速生长关系,即在叶片生长过程中SLA趋于稳定。
2.3.4 叶组织密度与叶长及叶体积的关系
LTD与LL、LV均为共同斜率(-0.449、-0.224)显著小于1的异速生长关系(P<0.001,表3,图2E—F),但常绿植物组LTD与LL的斜率(-0.711)却表现为与1无显著差异的等速生长关系,与总体异速生长趋势不一致,表明常绿植物倾向于限制LL以调整叶大小来适应环境的变化。两组关系的截距常绿植物组均显著高于落叶植物组(P<0.001),表明在某一给定的LL或LV下,常绿植物比落叶植物具有更大的LTD。此外,两组关系的斜率均沿共同主轴显著向下漂移(P<0.001)。
3 讨 论
3.1 叶性状的系统发育信号
桂西南岩溶区植物群落10个叶性状的Blomberg’s K值均小于1,未表现出较强的系统发育保守性,系统发育结构与性状结构并不完全一致。这主要是因为叶性状对气候变化非常敏感,其变异更多地受持续的环境过滤而非进化历史的影响[11-12]。这与车应弟等[13]、程毅康等[14]、王诗韵等[15]在甘南藏族自治州亚高寒草甸、海南岛热带云雾林和新疆艾比湖流域的研究结果一致。这种结果可能是很多原因造成的。首先,在资源限制环境中,生境过滤往往主导群落的构建过程。其次,系统发育结构与性状结构的匹配关系通常出现在较大的分类学尺度上,在物种水平上无法形成一一对应的关系。最后,某些非生物胁迫也可能促使远缘物种的耐受性表现出一定的相似性,从而产生趋同适应,这些性状有时并不表现出明显的系统发育信号。本研究中,除LWC、LDMC和LTD表现出显著的系统发育信号(P<0.05),其余7个叶性状均未检测到显著的系统发育信号,表明LWC、LDMC和LTD受到植物系统发育历史的显著影响,具有进化的保守性,这可能对维持物种的生存和繁衍具有重要意义;其余7个叶性状受环境因子的影响更大,这些性状的变异可促使植物有效地适应环境的变化,引起适应辐射。曹科等[16]、侯嫚嫚等[17]的研究结果表明多数功能性状具有显著的系统发育信号。造成这种差异的原因可能与物种种类、空间尺度、土壤水分、样本数量以及取样时间等不同有关。岩溶生境高度异质性,季节性和地质性干旱频发,群落处于植被恢复演替早期,环境过滤作用主导了群落构建过程,物种性状趋同;而古田山和长白山植被处于群落演替中后期,竞争排斥作用占相对优势,性状保守。
3.2 叶性状间的关联进化
性状间关联反映了植物对环境的特异性适应。无论是否考虑系统发育的影响,LW、LL、LA、LDM和LV以及SLA与LW、LL、LA间均存在显著正相关关系(P<0.05),而LTD与LW、LL、LA、LDM、LV、SLA以及LWC与LDMC间均存在显著负相关关系(P<0.05),表明这些性状间存在关联进化关系,他们之间的关系在植物类群的进化历史中普遍存在并持续至今,这些关系的维持对植物适应性进化至关重要,可能是植物生存、繁衍和扩散的重要基础[5]。在剔除系统发育的影响之后,LT与LW、LL、LA、LDM、LV、LTD之间的相关关系不再显著,表明LT与LW、LL、LA、LDM、LV、LTD间不存在关联进化或各自独立进化。张亚等[18]研究也显示,LT与气孔、叶脉之间不存在关联进化,这可能是自然选择压力所致,对物种的适应辐射具有重要促进意义。而具有显著系统发育信号的LWC、LDMC和LTD在进行PIC处理后,SLA与LDMC、LWC和LTD与LDMC、LWC间的相关性不再显著,表明系统发育关系对SLA与LDMC、LWC和LTD与LDMC、LWC间的耦合具有一定的影响。在系统发育的作用下,不同性状受基因和环境的调控程度不同,从而导致耦合关系发生变化。这也在一定程度上解释了系统发育关系在性状演化过程中所起的作用。
3.3 叶性状间的关系
异速生长关系被认为是生物界普遍存在的一种规律[19-20]。然而,本研究发现植物叶性状之间的生长关系并非都是异速生长关系。如LL与LW、LA与LDW间均表现为等速生长。相关研究表明,植物在干旱年份LA变小的过程中,LW具有优先降低的趋势;而在湿润年份LA变大过程中,LL显现优先增长的特征。在炎热、干旱环境下,随着降水的减少、水分有效性降低或气温升高,叶片最大宽度逐渐降低,LA快速减少[21],叶片降温能力持续加强[22],同时快速降低植物水分损失量[21],从而提高植物对高温和干旱胁迫的耐受性。植物通过降低LW和LA的适应策略来降低叶片生长代谢,呈现出LL与LW、LA与LDW等速生长的适应模式。这与已有的研究结果一致[1, 23]。LTD与LL总体呈异速生长关系,但在不同生活型水平间,常绿植物却表现为等速生长关系。这可能是因为常绿植物的冠层叶片数量较多且较为密集,邻体竞争对LL的限制作用更强;同时也表明常绿植物倾向于限制LL以适应邻体光资源竞争,维持种群的稳定和扩张。林华等[24]对苦竹叶片性状的研究也得出了相同的结果。
研究表明,异速生长关系的斜率能较好地反映叶性状对环境变化的敏感性[23]。本研究中,LA与LDW生长关系的斜率在常绿和落叶植物组间存在显著差异(P<0.05),表明常绿和落叶植物在最大光合生产能力和适应逆境之间的权衡存在较大差异。叶大小(LA与LDW)对环境变化较为敏感,可显著影响植物的光获取效率和水分利用效率[7]。在相同LDW下,落叶植物比常绿植物具有更大的LA,意味着落叶植物具有更高的光获取效率,能够利用夏季良好的水热条件迅速获得更多的碳水化合物完成生长。同时,采取落叶的策略来适应季节性低温和干旱胁迫。与落叶植物相反,常绿植物的LA较小,具有较高的主脉密度和较低的气孔导度,可实现水分利用效率的最大化[25]。这也是常绿植物对炎热、干旱环境在结构上表现出的一种适应性特征。其他几组性状之间的斜率在常绿和落叶植物组间均无显著差异,表明常绿和落叶植物在这几组性状上具有一致或相似的生长策略。
截距表征异速生长的起始点,反映叶性状对环境变化的可塑性[23]。LTD与LL、LV间的截距常绿植物组均显著高于落叶植物组(P<0.001),表明常绿植物LTD与LL、LV间的异速生长起始点的可塑性较大,其对异质生境的适应性更强[10]。水分胁迫下,植物会将更多光合产物用于防御组织构建,以增强其抗旱性[26],LTD增加,导致其LL、LA和LV显著减少。植物通过LTD与LL及LV之间的权衡最大程度地适应环境水分的变化。常绿植物对水分胁迫的容忍性较高;而落叶植物容忍水分胁迫的能力较差[27],多采用落叶的方式减少旱季蒸腾和水分消耗。研究表明,常绿是物种对低资源可用性的适应性反应,落叶是减少干旱胁迫期间水分损失的适应[28]。常绿和落叶植物通过截距上的差异来响应叶习性的变化[28-29]。
此外,LA、LL和LW以及LTD与LL、LV在常绿和落叶植物组间的斜率均沿共同主轴方向显著漂移(P<0.05),表明常绿植物较落叶植物具有更小的LA、LL、LW、LV和更大的LTD。物种性状的分异反映了植物对岩溶异质生境的适应,同时也表明常绿和落叶植物对环境的适应策略发生了偏移。在有限的资源环境下,常绿植物的叶片更倾向于资源保守型生长策略,其LA较小、LTD较大,有利于维持植物体内水分平衡和降低食叶天敌以及风、霜、雪等的危害。而落叶植物的叶片倾向于资源获取型生长策略,较大的LA具有较高的光获取效率和相对生长速率,能够捕获更多的光能、合成更多的碳水化合物,增强竞争力。常绿和落叶植物生态适应策略的差异,提示这两类植物存在明显的生态位分化[30],直接减少了二者对群落空间和资源的竞争强度,最终形成了亚热带常绿落叶阔叶混交林群落。这也与全球叶经济谱假说的预测一致,常绿植物靠近叶经济谱中“缓慢投资-收益”型物种的一端,而落叶植物则与之相反,位于叶经济谱“快速投资-收益”型物种的一端。常绿和落叶植物的分异特征对理解生态位分化、物种共存和群落构建机制等具有重要的理论和现实意义。在岩溶区生态修复和重建过程中,应充分利用常绿和落叶植物的生态位互补效应,营建近自然复层异龄混交林。多物种共存更能充分有效地利用有限的时空资源,进而在群落水平上促进物种丰富度和优势度。
4 结 论
综上所述,桂西南岩溶区植物叶性状未表现出较强的系统发育保守性,系统发育结构与性状结构并不完全一致。叶性状在不同生活型物种间的生长关系不尽相同。除常绿植物LTD与LL的生长关系与总体异速生长趋势不一致外,其余性状组合间的生长关系并未因生活型的不同而引起差异。LA、LL和LW以及LTD与LL、LV在常绿和落叶植物组间的斜率均沿共同主轴方向显著漂移(P<0.05),表明常绿和落叶植物对环境的适应策略发生了位移,常绿植物倾向于选择小叶、组织密度大的资源保守型生长策略,而落叶植物则倾向于选择大叶、组织密度小的资源获取型生长策略。生态位分化在很大程度上是促进亚热带常绿落叶阔叶混交林群落物种共存和生物多样性维持的重要机制。由于研究中并未对相关的环境因子进行测定,无法定量分析环境因子对叶性状及其异速生长关系的影响,在今后的研究中应予以关注。
参考文献:
[1] 唐玉瑞,赵成章,赵辉,等.不同光环境下洮河护岸林沙棘叶干重与叶面积、叶厚度间的关系[J].生态学杂志,2021,40(9): 2745-2753. TANG Y R, ZHAO C Z, ZHAO H, et al. The relationship between leaf dry mass and leaf area, leaf thickness of Hippophae rhamnoides under different light conditions in Taohe River riparian forest[J]. Chinese Journal of Ecology,2021,40(9): 2745-2753.
[2] 孟婷婷,倪健,王国宏.植物功能性状与环境和生态系统功能[J].植物生态学报,2007,31(1):150-165. MENG T T, NI J, WANG G H. Plant functional traits, environments and ecosystem functioning[J]. Chinese Journal of Plant Ecology,2007,31(1):150-165.
[3] 贺鹏程,叶清.基于植物功能性状的生态学研究进展:从个体水平到全球尺度[J].热带亚热带植物学报,2019,27(5): 523-533. HE P C, YE Q. Plant functional traits: from individual plant to global scale[J]. Journal of Tropical and Subtropical Botany, 2019,27(5):523-533.
[4] 王常顺,汪诗平.植物叶片性状对气候变化的响应研究进展[J].植物生态学报,2015,39(2):206-216. WANG C S, WANG S P. A review of research on responses of leaf traits to climate change[J]. Chinese Journal of Plant Ecology, 2015,39(2):206-216.
[5] FU P L, ZHU S D, ZHANG J L, et al. The contrasting leaf functional traits between a karst forest and a nearby nonkarst forest in southwest China[J]. Functional Plant Biology, 2019,46(10):907-915.
[6] 袁丛军,程娟,丁访军,等.喀斯特森林优势树种蒸腾特征及其对气孔和气象因子的响应[J].中南林业科技大学学报, 2022,42(7):96-105. YUAN C J, CHENG J, DING F J, et al. Transpiration characteristics of dominant tree species and their responses to stomatal and meteorological factors in karst regions[J]. Journal of Central South University of Forestry Technology, 2022,42(7):96-105.
[7] 李耀琪,王志恒.植物叶片形态的生态功能、地理分布与成因[J].植物生态学报,2021,45(10):1154-1172. LI Y Q, WANG Z H. Leaf morphological traits: ecological function, geographic distribution and drivers[J]. Chinese Journal of Plant Ecology,2021,45(10):1154-1172.
[8] CAVENDER-BARES J, KOZAK K H, FINE P V A, et al. The merging of community ecology and phylogenetic biology[J]. Ecology Letters,2009,12(7):693-715.
[9] ZHANG S B, FERRY S J W, ZHANG J L, et al. Spatial patterns of wood traits in China are controlled by phylogeny and the environment[J]. Global Ecology and Biogeography,2011,20(2): 241-250.
[10] 庞世龙,欧芷阳,凌福诚,等.桂西南岩溶区18种适生植物叶性状变异及经济谱[J].生态学杂志,2021,40(10):3041-3049. PANG S L, OU Z Y, LING F C, et al. Leaf trait variations and leaf economic spectrum of 18 plant species suitable in a karst area of southwest Guangxi[J]. Chinese Journal of Ecology,2021,40(10): 3041-3049.
[11] WRIGHT I J, DONG N, MAIRE V, et al. Global climatic drivers of leaf size[J]. Science,2017,357(6354):917-921.
[12] LI Y Q, REICH P B, SCHMID B, et al. Leaf size of woody dicots predicts ecosystem primary productivity[J]. Ecology Letters, 2020,23(6):1003-1013.
[13] 车应弟,刘旻霞,李俐蓉,等.基于功能性状及系统发育的亚高寒草甸群落构建[J].植物生态学报,2017,41(11):1157-1167. CHE Y D, LIU M X, LI L R, et al. Exploring the community assembly of subalpine meadow communities based on functional traits and community phylogeny[J]. Chinese Journal of Plant Ecology,2017,41(11):1157-1167.
[14] 程毅康,张辉,王旭,等.功能多样性和谱系多样性对热带云雾林群落构建的影响[J].植物生态学报,2019,43(3):217-226. CHENG Y K, ZHANG H, WANG X, et al. Effects of functional diversity and phylogenetic diversity on the tropical cloud forest community assembly[J]. Chinese Journal of Plant Ecology, 2019,43(3):217-226.
[15] 王诗韵,吕光辉,蒋腊梅,等.不同尺度下艾比湖典型植物群落功能多样性和系统发育多样性研究[J].生态环境学报, 2020,29(5):889-900. WANG S Y, LYU G H, JIANG L M, et al. Multi-scale analysis on functional diversity and phylogenetic diversity of typical plant community in Ebinur Lake[J]. Ecology and Environmental Sciences,2020,29(5):889-900.
[16] 曹科,饶米德,余建平,等.古田山木本植物功能性状的系统发育信号及其对群落结构的影响[J].生物多样性,2013,21(5): 564-571. CAO K, RAO M D, YU J P, et al. The phylogenetic signal of functional traits and their effects on community structure in an evergreen broad-leaved forest[J]. Biodiversity Science, 2013,21(5):564-571.
[17] 侯嫚嫚,李晓宇,王均伟,等.长白山针阔混交林不同演替阶段群落系统发育和功能性状结构[J].生态学报,2017,37(22): 7503-7513. HOU M M, LI X Y, WANG J W, et al. Phylogenetic development and functional structures during successional stages of conifer and broad-leaved mixed forest communities in Changbai Mountains[J]. Acta Ecologica Sinica,2017,37(22):7503-7513.
[18] 张亚,杨石建,孙梅,等.基部被子植物气孔性状与叶脉密度的关联进化[J].植物科学学报,2014,32(4):320-328. ZHANG Y, YANG S J, SUN M, et al. Stomatal traits are evolutionarily associated with vein density in basal angiosperms[J]. Plant Science Journal,2014,32(4):320-328.
[19] SUN S C, JIN D M, SHI P L. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship[J]. Annals of Botany, 2006,97(1):97-107.
[20] 李晓红,徐健程,肖宜安,等.武功山亚高山草甸群落优势植物野古草和芒异速生长对气候变暖的响应[J].植物生态学报, 2016,40(9):871-882. LI X H, XU J C, XIAO Y A, et al. Responses in allometric growth of two dominant species of subalpine meadow: Arundinella anomala and Miscanthus sinensis to climate warming in Wugongshan mountains, China [J]. Chinese Journal of Plant Ecology,2016,40(9):871-882.
[21] 李永华,卢琦,吴波,等.干旱区叶片形态特征与植物响应和适应的关系[J].植物生态学报,2012,36(1):88-98. LI Y H, LU Q, WU B, et al. A review of leaf morphology plasticity linked to plant response and adaptation characteristics in arid ecosystems[J]. Chinese Journal of Plant Ecology,2012,36(1): 88-98.
[22] YATES M J, VERBOOM G A, REBELO A G, et al. Ecophysiological significance of leaf size variation in Proteaceae from the Cape Floristic Region[J]. Functional Ecology,2010, 24(3):485-492.
[23] 李晓玲,温浩然,王雪松,等.三峡库区不同生境下中华蚊母树叶片表型可塑性及其与土壤环境因子的关系[J].生态学报, 2018,38(10):3581-3591. LI X L, WEN H R, WANG X S, et al. Phenotypic plasticity of Distylium chinense leaves in relation to soil environmental factors in heterogeneous habitats in the Three Gorges Reservoir Region[J]. Acta Ecologica Sinica,2018,38(10):3581-3591.
[24] 林华,陈双林,郭子武,等.苦竹叶片性状及其异速生长关系的密度效应[J].林业科学研究,2017,30(4):617-623. LIN H, CHEN S L, GUO Z W, et al. Allometric relationship among leaf traits in different stand density of Pleioblastus amarus[J]. Forest Research,2017,30(4):617-623.
[25] NARDINI A, ?UNAPUU-PIKAS E, SAVI T. When smaller is better: leaf hydraulic conductance and drought vulnerability correlate to leaf size and venation density across four Coffea arabica genotypes[J]. Functional Plant Biology, 2014,41(9):972-982.
[26] 时应贵,熊德成,叶旺敏,等.土壤和大气增温对杉木幼树细根生物量和叶片性状的影响[J].生态学杂志,2021,40(7):1916-1923. SHI Y G, XIONG D C, YE W M, et al. Effects of soil and air warming on fine root biomass and leaf traits of Chinese fir saplings[J]. Chinese Journal of Ecology,2021,40(7):1916-1923.
[27] 金鹰,王传宽.植物叶片水力与经济性状权衡关系的研究进展[J].植物生态学报,2015,39(10):1021-1032. JIN Y, WANG C K. Trade-offs between plant leaf hydraulic and economic traits[J]. Chinese Journal of Plant Ecology,2015,39(10): 1021-1032.
[28] 王钊颖,陈晓萍,程英,等.武夷山49种木本植物叶片与细根经济谱[J].植物生态学报,2021,45(3):242-252. WANG Z Y, CHEN X P, CHENG Y, et al. Leaf and fine root economics spectrum across 49 woody plant species in Wuyi mountains[J]. Chinese Journal of Plant Ecology,2021,45(3): 242-252.
[29] 李俊慧,彭国全,杨冬梅.常绿和落叶阔叶物种当年生小枝茎长度和茎纤细率对展叶效率的影响[J].植物生态学报, 2017,41(6):650-660. LI J H, PENG G Q, YANG D M. Effect of stem length to stem slender ratio of current-year twigs on the leaf display efficiency in evergreen and deciduous broadleaved trees[J]. Chinese Journal of Plant Ecology,2017,41(6):650-660.
[30] 车俭,郑洁,蒋娅,等.中国亚热带森林动态监测样地常绿和落叶木本被子植物谱系结构及生态习性差异[J].植物生态学报,2020,44(10):1007-1014. CHE J, ZHENG J, JIANG Y, et al. Separation of phylogeny and ecological behaviors between evergreen and deciduous woody angiosperms in the subtropical forest dynamics plots of China[J]. Chinese Journal of Plant Ecology,2020,44(10):1007-1014.
[本文编校:吴 毅]
猜你喜欢
中国公路(2022年15期)2022-09-25 09:18:16
建材发展导向(2022年18期)2022-09-22 07:14:02
湖南水利水电(2021年6期)2022-01-18 06:07:02
建材发展导向(2021年9期)2021-07-16 07:10:42
国防交通工程与技术(2020年2期)2020-05-25 06:46:18
中华建设(2019年2期)2019-08-01 05:57:58
山东工业技术(2016年15期)2016-12-01 05:32:02
水科学与工程技术(2016年3期)2016-07-10 15:12:44
水利科技与经济(2016年9期)2016-04-22 01:07:50
河南科技(2014年12期)2014-02-27 14:10:26
