
地西泮通过let-7a-5p/MYD88轴抑制LPS诱导的细胞焦亡和炎症从而缓解小鼠肺纤维化
2024-12-11 00:00:00宋端怡李赟唐雪芳李化陶康
南方医科大学学报 2024年11期


关键词:地西泮;let-7a-5p;MYD88;细胞焦亡;肺纤维化
肺纤维化是一种严重危害人类健康的常见呼吸道疾病[1],并伴有炎症反应[2]。然而,由于肺纤维化发展的复杂性,未知的潜在机制限制了其临床治疗靶点的发现。脂多糖(LPS)[3]或博来霉素(BLM)[4]是用于诱导肺纤维化动物模型进行研究的理想药物,已被证明可以完美模拟肺纤维化的发展。由于细菌感染通过分泌LPS诱导肺纤维化[5],因此有必要探索LPS诱导的肺纤维化的治疗药物。地西泮(Dia)是一类γ-氨基丁酸A受体(GABAA Rs)激动剂,它能抑制中枢神经系统的神经传递并调节大脑内的神经炎症[6]以及外周器官的炎症性疾病,例如哮喘[7]。但目前缺乏Dia对LPS诱导的炎症影响的报道。
细胞焦亡与多种疾病有关,如胰腺癌[8],肝纤维化[9, 10]和肺相关的疾病[11-13]。细胞焦亡通过分泌促炎细胞因子IL-18 和IL-1β加剧炎症相关疾病的进展[14, 15]。LPS诱导细胞焦亡并促进急性肺损伤和纤维化[13]。此外,LPS诱导巨噬细胞和癌细胞的Caspase-11依赖性细胞焦亡[16]。同样,目前Dia对LPS诱导的焦亡的影响研究较少。
microRNA(miRNA)是一类长度约为22 nt的内源性非编码单链小RNA,其参与多种生物功能并且在细胞间通讯中发挥重要作用[17]。let-7家族是miRNA中最为人熟知的一个家族,let-7 家族通常作为抑癌基因miRNA 发挥作用,以抑制肿瘤生长和转移[18]。let-7a-5p作为let-7家族的一员,也在调控细胞死亡途径中发挥作用。比如,let-7a-5p可以靶向BCL-xL影响肺癌细胞的自噬和凋亡[19]。还有富含let-7a-5p 的细胞外囊泡可以缓解急性肺损伤(ALI)中的炎症以及阻碍 ALI 纤维化过程[20]。目前没有let-7a-5p 对细胞焦亡影响的其他新报道,仍需继续let-7a-5p调控焦亡的机制研究。
MYD88是Toll样受体(TLR)和白细胞介素-1(IL-1)受体家族成员下游炎症信号通路的典型适配器[21]。MYD88在炎症反应[22]中充当中心枢纽,并且可以诱导来自位于质膜或内体的多种受体的信号传导。此外,MYD88信号传导可导致产生促炎或抗炎细胞因子以及I 型干扰素[23]。我们通过生物信息学网站TargetScan(http://www.targetscan.org/vert_72/)预测发现MYD88是let-7a-5p的一个靶标。基于此,我们推测let-7a-5p可能通过靶向作用调控MYD88的表达,进而影响LPS诱导的细胞焦亡和炎症反应。因此,本研究的目标是探讨Dia是否通过let-7a-5p/MYD88轴来抑制LPS诱导的细胞焦亡和炎症反应,从而缓解肺纤维化进程。
1 材料和方法
1.1 材料
C57BL/6小鼠[昆明医科大学实验动物部,合格证编号:SCXK(滇)K2020-0004];MRC-5细胞(中国科学院上海细胞库);LPS(Sigma);LipofectamineTM2000 转染试剂盒、Trizol 试剂、RT-qPCR 试剂盒(Invitrogen);ELISA试剂盒(上海酶联生物科技有限公司);Westernblotting 实验一抗Col‑Ⅰ、Col‑Ⅲ、α‑SMA、NLRP3、Caspase-1、MYD88、ASC、GSDMD和β‑actin(Abcam);CCK-8试剂盒、双荧光素酶报告基因检测试剂盒(上海翌圣生物科技股份有限公司)。
1.2 方法
1.2.1 动物模型 将体质量为18~22g 的C57BL/6 小鼠随机分为6 组。LPS组(n=15):小鼠分别通过肺注射、腹腔注射和气管内管注射3 次1.5 mg/kg LPS,间隔24 h[24]。正常对照组(n=15):注射等量生理盐水作为对照组。LPS+Dia组(n=10):LPS处理结束2 h后,小鼠腹腔注射Dia(0.7 mg/10 μL/g)。LPS+Dia+let-7a-5p mimic组:LPS+Dia 处理的小鼠尾静脉注射let-7a-5p mimic。LPS+Dia+ST2825(n=5):LPS+Dia 处理的小鼠腹腔注射20 mg/kg ST2825。LPS+Dia+let-7a-5p mimic+pc-DNA-MYD88(n=5):LPS+Dia+let-7a-5p mimic 处理的小鼠尾静脉注射pc-DNA-MYD88 质粒包装的慢病毒。在第21 天处死小鼠进行进一步实验。收集小鼠血清和肺组织,左肺组织用于组织学观察,右肺组织用于检测相关基因表达水平。所有动物实验通过了昆明医科大学动物伦理委员会的批准(伦理批号:kmmu20211589)。
1.2.2 细胞培养及转染 将人肺成纤维细胞MRC-5 细胞置于杜尔贝科改良的含有10%的胎牛血清(FBS)和有5%的CO2的完全培养基(DMEM)中,37 ℃的培养箱中培养。在细胞融合达到约70%~80%后,对细胞进行胰蛋白酶消化并稀释至1×104/mL。然后将细胞悬浮液按照1 mL/孔接种到六孔板中并培养24 h。随后,分别用浓度为0.1、0.5 和1 mg/mL的LPS 处理MRC-5细胞24 h。加入LPS 2 h 后,将细胞暴露于Dia(1、5 和10 μmol/L)12 h,并收集上述细胞用于进一步实验。在细胞培养至融合率达到60%~80% 时,使用LipofectamineTM2000 转染试剂盒将let-7a-5p mimic 和pc-DNA-MYD88 及阴性对照NC mimic,pc-DNA转染到MRC-5细胞中,用MYD88抑制剂ST2825(1 μmol/L)处理细胞4 h。
1.2.3 CCK-8实验 选择对数生长期的细胞,将细胞稀释至约5×104/mL后,接种于96孔板中。各组实验结束后,加入CCK-8试剂,10 μL/孔,37 ℃孵育3 h后,酶标检测吸光度值A450 nm,用每组吸光度值与正常组的吸光度值的比值来计算细胞增殖活力。
1.2.4 RT-qPCR实验 使用Trizol 试剂从MRC-5 细胞中提取细胞总RNA。取2 μg 总RNA 逆转录酶合成cDNA。PCR的两步法反应条件为:预变性(95 ℃下维持15 min),变性(95 ℃下维持10 s),退火(30 s)和延伸(30 s),循环40 次。以GAPDH(mRNA)和U6(miRNA)为内参。根据公式2-ΔΔCt值计算基因表达量,所得值即为基因的最终表达情况。在GenBank 上查找对应的基因序列,用Primer Premier 5设计特异性引物,引物由上海捷瑞生物工程有限公司合成(表1)。
1.2.5 Western blotting 实验 使用RIPA裂解缓冲液从细胞和组织中提取总蛋白。然后通过十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)分离目标蛋白,并转移到聚偏氟乙烯(PVDF)膜上,并用5%牛血清白蛋白封闭膜2 h。用稀释的一抗Col-Ⅰ(1∶5000)、Col-Ⅲ(1∶1000)、α‑SMA(1∶10000)、NLRP3(1∶1000)、Caspase-1(1∶10000)、ASC(1∶1000)、GSDMD(1∶1000)、MYD88(1∶1000)和β-actin(1∶5000)在4 ˚C摇床上孵育过夜,洗膜后将二抗(1∶4000)与膜一起孵育。随后,将膜暴露于增强化学发光底物中,使用Western blotting分析系统显示蛋白条带。
1.2.6 ELISA实验 收集每组细胞培养上清液和小鼠肺组织,按ELISA试剂盒说明进行操作,测量IL-4、IL-6、TGF-β和TNF-α水平,酶标仪测量A450值,通过A值计算出待测样本中各蛋白浓度。
1.2.7 免疫荧光染色 细胞在铺14 mm爬片的24 孔板中培养用于免疫荧光测定。对于α-SMA染色,将细胞用4% 多聚甲醛固定15 min 后,置于5% Triton×100PBS 中10 min,将细胞在10% 山羊血清中室温封闭30 min,与一抗α-SMA(1∶200)在4 ℃下孵育过夜,然后与 Alexa Fluor 488(Invitrogen/Life Technologies) 在室温下放置2 h。细胞核用DAPI复染。在显微镜下观察图像,并用ImagePro Plus 软件对图像进行分析。
1.2.8 双荧光素酶报告基因实验 TargetScan (http://www.targetscan.org/vert_72/) 用于预测 let-7a-5p 的潜在靶标,随后通过双荧光素酶报告基因检测进行确认。构建双荧光报告质粒,将对数生长期的MRC-5细胞以1×105/孔接种到24孔细胞板上,将NC-mimic、let-7a-5pmimic 与MYD88- 野生型(MYD88-WT)或突变型(MYD88-MUT)重组质粒按照LipofectamineTM 2000说明书步骤转染至MRC-5细胞。培养箱内培养48 h后,参照双荧光素酶报告基因检测试剂盒步骤检测荧光素酶活性。
1.2.9 Masson 染色 将小鼠肺组织固定于10%中性福尔马林固定液中,常规脱水包埋后切片。用Weiger 氏铁苏木素染色5~10 min,流水冲洗。将切片用1%盐酸酒精分化,并用流水冲洗10 min。用丽春红酸性品红液染对切片进行染色,染色时间为5~10 min,染色后用蒸馏水对切片进行快速冲洗。然后用1%磷钼酸水溶液处理约5 min,1%冰醋酸处理1 min,95%酒精脱水。最后用中性树胶封固,于显微镜下观察。
1.2.10 统计学分析 结果以均数±标准差表示。采用GraphPad Prism 7.0软件对数据进行分析及绘制相关定量图,所有数据在进行两组间比较时均采用t检验,所有数据在进行多组间比较时均采用单因素方差分析。当Plt;0.05时认为差异有统计学意义。
2 结果
2.1 LPS诱导细胞焦亡和炎症并且引起肺纤维化
细胞结果显示,LPS 以浓度依赖性的方式抑制MRC-5细胞的活力(图1A),因此,选择1 mg/mL的LPS用于后续实验。ELISA结果显示,LPS诱导后,MRC-5细胞中炎症相关因子IL-4、IL-6、TGF-β和TNF-α的浓度均升高(Plt;0.001,图1B),在小鼠肺组织中也得到了同样的结果(Plt;0.05,图1C)。Western blotting 结果显示,LPS 诱导后,MRC-5 细胞中的纤维化相关蛋白Col-Ⅰ、Col-Ⅲ、α-SMA和细胞焦亡相关蛋白ASC、GSDMD-N、NLRP3、Caspase-1、的表达量均升高(Plt;0.01,图1D、E),在动物实验中得到的也是相同的结果(Plt;0.05,图1F、G)。对纤维化相关蛋白α-SMA进行免疫荧光染色,发现LPS促进了MRC-5细胞和小鼠肺组织中α-SMA的含量(图1H、I)。Masson染色结果显示,LPS诱导了小鼠肺组织的纤维化(图1J)。
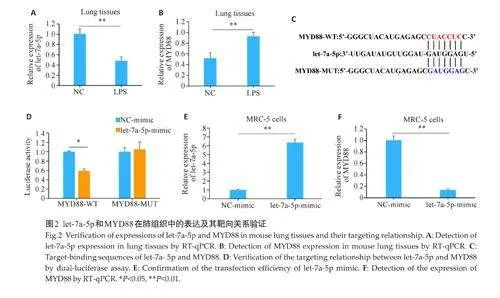
2.2 let-7a-5p和MYD88在肺组织中的表达及其靶向关系验证
RT-qPCR 检测发现,与NC 组相比,let-7a-5p 在LPS 组中表达下调(Plt;0.01,图2A),而MYD88 在LPS组中表达上调(Plt;0.01,图2B)。TargetScan预测结果显示MYD88是let-7a-5p的靶基因,二者之间有靶向结合序列(图2C)。双荧光素酶实验进一步对靶向关系进行验证,结果显示let-7a-5p mimic 降低了野生型MYD88(MYD88-WT)的荧光素酶活性(Plt;0.05,图2D),对突变型MYD88(MYD88-MUT)的荧光素酶活性无影响(图2D)。两者的靶向调控关系通过RT-qPCR实验进行确认。转染let-7a-5p mimic后,let-7a-5p mimic的表达上调(Plt;0.01,图2E),说明转染成功。RT-qPCR结果显示,与NC-mimic组相比,let-7a-5p mimic组MYD88的表达下调(Plt;0.01,图2F)。
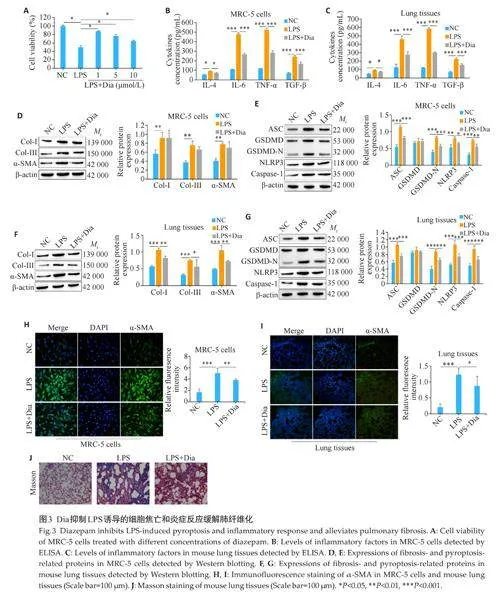
2.3 Dia抑制LPS诱导的细胞焦亡和炎症反应缓解肺纤维化
用不同浓度的Dia(1、5、10 μmol/L)处理LPS 诱导的MRC-5 细胞,CCK-8 结果显示,MRC-5 细胞对1 μmol/L的Dia最敏感(Plt;0.05,图3A),后续实验使用1 μmol/L 的Dia 进行。ELISA 结果显示,LPS 组中MRC-5细胞和小鼠肺组织中炎症相关因子IL-4、IL-6、TGF-β和TNF-α的浓度均高于NC组(Plt;0.001,图3B、C),而与LPS 组相比,LPS+Dia 组细胞和组织中IL-4、IL-6、TGF-β和TNF-α的浓度均降低(Plt;0.05,图3B、C)。Western blotting 结果显示,与NC组相比,LPS 组中MRC-5 细胞和小鼠肺组织中的纤维化相关蛋白Col-Ⅰ、Col-Ⅲ、α-SMA 和细胞焦亡相关蛋白ASC、GSDMD-N、NLRP3、Caspase-1 的表达量均升高(Plt;0.05,图3D~G);而与LPS 组相比,LPS+Dia 组细胞和组织中这些蛋白的表达量均降低(Plt;0.05,图3D~G)。α-SMA 的免疫荧光染色结果显示,LPS 组诱导增强的α-SMA的荧光强度,而在LPS+Dia组中荧光强度又减弱(图3H、I)。Masson 染色结果显示,LPS组诱导加重的纤维化程度而在LPS+Dia组中纤维化程度得到了一定的缓解(图3J)。
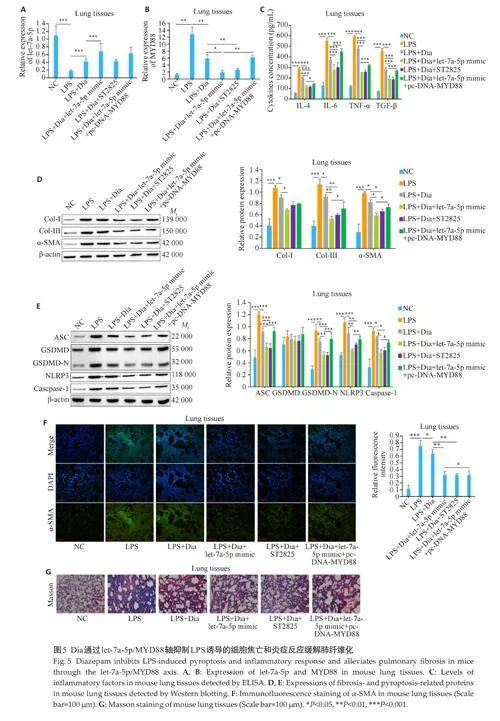
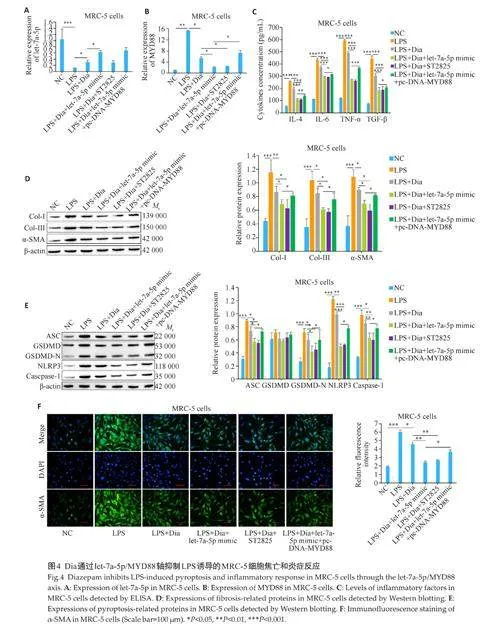
2.4 Dia通过let-7a-5p/MYD88轴抑制LPS诱导的细胞焦亡和炎症反应缓解肺纤维化
RT-qPCR结果显示,在MRC-5细胞和小鼠肺组织中,let-7a-5p在LPS组中的表达低于NC组(Plt;0.001),而在LPS+Dia组中的表达高于LPS组(Plt;0.05),在LPS+Dia+let-7a-5p mimic组中的表达进一步增加(Plt;0.05),而MYD88 抑制剂(ST2825)和过表达MYD88(pc-DNA-MYD88)的处理对Dia和Dia+let-7a-5p mimic 促进let-7a-5p 表达的作用不产生影响(图4A、5A)。MYD88在MRC-5细胞和小鼠肺组织中的表达与let-7a-5p 相反,并且ST2825 和pc-DNA-MYD88 的处理一定程度上可以逆转Dia和Dia+let-7a-5p mimic对MYD88表达的抑制作用(图4B、5B)。ELISA 和Westernblotting 结果显示,在MRC-5 细胞和小鼠肺组织中,炎症相关因子(IL-4、IL-6、TGF-β、TNF-α)和纤维化相关蛋白(Col-Ⅰ、Col-Ⅲ、α-SMA)以及细胞焦亡相关蛋白(ASC、GSDMD-N、NLRP3、Caspase-1)在LPS组中的含量和表达高于NC 组(Plt;0.05),而在LPS+Dia 组中的含量和表达低于LPS 组(Plt;0.05),在LPS+Dia+let-7a-5p mimic 组和LPS+Dia+ST2825 组中的含量和表达与LPS+Dia 组相比均下降(Plt;0.05),且这种降低在LPS+Dia+let-7a-5p mimic+pc-DNA-MYD88组得到一定的逆转(图4C~E、图5C~E)。α‑SMA 的免疫荧光染色结果显示,在MRC-5 细胞和小鼠肺组织中,α‑SMA 的荧光强度在LPS 组中高于NC 组(Plt;0.001),而在LPS+Dia 组中的荧光强度低于LPS 组(Plt;0.05),在LPS+Dia+let-7a-5p mimic 组和LPS+Dia+ST2825 组中的荧光强度与LPS+Dia 组相比均降低(Plt;0.01),且这种降低在LPS+Dia+let-7a-5p mimic+pc-DNA-MYD88 组得到一定的逆转(图4F、5F)。Masson 染色结果显示,在小鼠肺组织中,肺纤维化程度在LPS 组中高于NC组,而在LPS+Dia 组中的肺纤维化程度低于LPS组,在LPS+Dia+let-7a-5p mimic 组和LPS+Dia+ST2825 组中的肺纤维化程度与LPS+Dia 组相比均降低,且这种降低在LPS+Dia+let-7a-5pmimic+pc-DNA-MYD88组得到一定的逆转(图5G)。
3 讨论
肺纤维化是一种复杂的病理过程,目前临床上缺乏有效的治疗方法[1]。因此,深入揭示肺纤维化的潜在机制对于开发新的治疗策略至关重要。本研究通过建立LPS诱导的细胞和小鼠肺纤维化模型,模拟了细菌感染诱导的肺纤维化的发展过程。先前的研究表明,LPS通过抑制肺细胞增殖和促进肺细胞凋亡,以及诱导肺组织炎症,加剧了肺纤维化的进展[25, 26]。本研究进一步探讨了Dia对肺纤维化进展的影响及其分子机制。
Dia作为一种苯二氮卓类药物,已被证明能够通过影响先天免疫细胞减少促炎细胞因子的释放[27, 28],并在流感感染期间抑制炎症反应[29, 30]。鉴于肺纤维化是一种慢性炎症相关反应的表现,Dia可能作为肺纤维化的潜在治疗药物。我们先前研究已经证明了Dia 可以有效缓
解小鼠肺纤维化进程[24]。本研究再次证实Dia 能够抑制LPS 诱导的MRC-5 细胞中炎症相关因子(如IL-4、IL-6、TGF-β和TNF-α)的表达;还发现Dia 可以抑制NLRP3、Caspase-1、ASC 和GSDMD-N 的表达。NLRP3炎性小体激活是导致细胞焦亡的重要原因。焦亡过程中,ASC会被招募到炎症小体复合物中,促进Caspase-1 的活化,活化的Caspase-1 又会切割GSDMD,产生具有活性的GSDMD-N片段,GSDMDN会导致细胞膜通透性增加,最终引发细胞肿胀和破裂[31]。有报道称,应用焦亡抑制剂可以降低肺组织的炎症反应,抑制肺纤维化[32]。所以Dia对炎症因子和焦亡蛋白的抑制作用潜在说明了其可以通过抑制细胞焦亡和炎症反应,从而缓解肺纤维化。
此外,我们发现let-7a-5p 在LPS诱导的MRC-5 细胞中表达下调,而Dia 的处理可以恢复let-7a-5p 的表达。let-7a-5p 作为一种微小RNA,在多种疾病中发挥重要作用,尤其是在癌症中被广泛研究,被描述为肿瘤抑制因子或致癌miRNA[ 33]。有研究指出,在尘肺病中可以检测到外源性let-7a-5p的表达,并且通过增加let-7a-5p表达有可能降低尘肺合并肺癌死亡率[34]。我们的实验数据也显示,let-7a-5p 在肺纤维化中的表达降低,且过表达let-7a-5p可抑制细胞焦亡和炎症反应缓解肺纤维化。MYD88是一种关键的衔接蛋白,参与控制先天免疫反应和炎症的Toll样和IL-1受体家族信号传导,是炎症性肺病的治疗靶点[35]。MYD88依赖的信号通路可激活NF-κB,从而引起炎症因子释放[36]。研究表明,藻蓝蛋白通过在早期抑制 TLR2-MYD88-NF-κB 通路来减轻BLM诱导的肺损伤和纤维化,并在纤维化过程中减弱EMT[37]。我们的研究结果发现,MYD88在LPS诱导的肺纤维化中表达上调,在加入MYD88 抑制剂ST2825后LPS诱导的肺纤维化的细胞焦亡和炎症反应能得到缓解。我们还确定了let-7a-5p 与MYD88 之间具有靶向结合位点,过表达MYD88 可以有效逆转let-7a-5p对炎症和焦亡的抑制作用。说明let-7a-5p能通过调控MYD88 的表达来影响LPS诱导的细胞焦亡和炎症反应。
综上所述,Dia可以通过上调let-7a-5p的表达靶向负调控MYD88,从而抑制LPS诱导的细胞焦亡和炎症反应缓解肺纤维化。
猜你喜欢
中老年保健(2022年2期)2022-11-25 23:46:31
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:24:52
昆明医科大学学报(2022年4期)2022-05-23 13:04:50
医学综述(2022年7期)2022-04-19 12:31:12
昆明医科大学学报(2020年12期)2021-01-26 00:44:42
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:26
国际呼吸杂志(2019年21期)2019-11-25 09:52:20
中国报道(2018年2期)2018-04-20 04:12:46
金融博览(2016年7期)2016-08-16 18:44:41
绿色中国(2016年1期)2016-06-05 09:02:59
