
STING高表达通过调控TLR4/NF-κB/NLRP3通路和影响炎症与凋亡水平促进小鼠肾脏缺血再灌注损伤
2024-11-03 00:00:00陶怀祥骆金光闻志远虞亘明苏萧王鑫玮关翰陈志军
南方医科大学学报 2024年7期
摘要:目的 探讨STING在肾缺血再灌注损伤(IRI)中的表达水平以及相关作用机制。方法 在体内水平,将24只C57BL/6小鼠分为假手术组(Sham)、IRI组、IRI+药物溶剂组(IRI+DMSO)、IRI+SN-011组,6只/组。通过肾动脉夹闭方法建立IRI模型,通过血清肌酐和尿素氮检测、PAS 染色检测肾组织损伤变化,采用RT-qPCR、ELISA、Western blotting 和IHC 法检测肾组织中STING、KIM-1、Bcl-2、Bax、caspase-3、TLR4、P65、NLRP3、caspase-1、CD68、MPO、 IL-1β、IL-6、TNF-α 的水平。在体外水平,将HK-2细胞分为对照组、缺氧复氧(H/R)组、H/R+药物溶剂组(H/R+DMSO)、H/R+SN-011组,用厌氧包模拟缺氧环境,RT-qPCR和Western blotting 法检测STING表达水平,流式细胞术检测各组细胞凋亡率。结果 在体内水平,与Sham组相比,IRI 组的PAS 染色显示组织损伤增加(Plt;0.05),小鼠血清肌酐、尿素氮含量以及组织KIM-1、STING、TLR4、P65、NLRP3、caspase-1、caspase-3、Bax、CD68、MPO、IL-1β、IL-6、TNF-α 表达水平升高(Plt;0.05),Bcl-2 水平降低(Plt;0.05),SN-011 抑制STING表达后,逆转了上述结果(Plt;0.05)。在体外水平,与对照组相比,H/R组STING的mRNA与蛋白水平升高(Plt;0.05),流式细胞仪检测显示细胞凋亡率上升(Plt;0.05),SN-011抑制STING表达,细胞凋亡率下降(Plt;0.05)。结论 STING在肾脏IRI中表达水平上升,且可通过作用于TLR4/NF-κB/NLRP3通路以及影响炎症与凋亡水平促进肾损伤。
关键词:STING;TLR4;NF-κB;NLRP3;肾缺血再灌注;炎症;凋亡
急性肾损伤(AKI)发病率死亡率较高,且其极易发展为慢性肾脏病或终末期肾脏病[1, 2]。肾缺血再灌注损伤(RIRI)被视为AKI 发生的主要原因之一[3]。既往研究显示,RIRI过程中,包括炎症级联反应和细胞凋亡在内的多种重要机制扮演了关键角色[4-6]。但目前有关RIRI中炎症和细胞凋亡的详细分子机制尚未被充分解明,而这些机制的深入了解对于开发新的治疗靶点与方法具有重要意义。干扰素基因刺激因子(STING)是一种与内质网相关的免疫衔接蛋白,可影响诸多细胞因子或免疫细胞产生与成熟,如干扰素I、巨噬细胞和T细胞等,在多种生理与病理情况下发挥作用,包括抗病毒反应、自身免疫性疾病以及肿瘤免疫等,但其在RIRI中的相关作用以及深入机制尚不清楚。研究显示,STING发生活化后,可激活下游STING-IKK-NF-κB 通路和STING-TBK1-IRF3通路,这使其在炎症反应与细胞死亡等诸多病理或生理过程中发挥重要作用[7, 8]。由于RIRI在发生发展中具有炎症反应与肾小管上皮细胞死亡相互促进等特点,故STING在RIRI中的调控作用值得进一步探讨。本研究通过建立肾脏IRI模型以及细胞H/R模型,探究STING在RIRI中表达水平变化,并采用新型抑制剂SN-011抑制STING表达,观察肾脏损伤程度以及炎症凋亡水平变化,探讨STING相关作用机制。
1 材料和方法
1.1 材料
1.1.1 实验动物
雄性SPF 级C57BL/6 小鼠24 只,6~8周龄,20~24 g,购自杭州子源实验动物公司。饲养条件:温度23±2 ℃,相对湿度(55±5)%,12 h昼夜交替,自由饮食进水条件下,饲养1周进行实验。本研究所有动物实验经蚌埠医学院第一附属医院伦理委员会批准通过(伦理批号:伦动科批字[2022]第066号)。
1.1.2 主要试剂与仪器
STING抑制剂SN-011(MCE);STING、TLR4、caspase-3、NF-κB p65抗体(Proteintech);KIM-1 抗体(Santa);Bcl-2、Bax、caspase-1 抗体(HUABIO);NLRP3(Abcam);β-Tubulin、GAPDH抗体以及羊抗小鼠二抗、羊抗兔二抗(Beyotime);PCR逆转录试剂盒、PerfectStart Green Qpcr SuperMix(TransgenBiotech);蛋白定量试剂盒、PAGE凝胶快速制备试剂盒(雅酶);ELISA试剂盒(Mlbio);细胞凋亡检测试剂盒(Servicebio);垂直电泳仪、曝光仪(Bio-Rad);荧光定量PCR仪(Roche);厌氧产气袋(三菱)。
1.2 方法
1.2.1 小鼠肾缺血再灌注模型的制备
10%戊巴比妥钠按照4 mL/kg 剂量麻醉小鼠,剃毛器剃净小鼠背部毛发,进行碘伏消毒,然后在肾区分层游离皮肤、筋膜与肌肉,游离双侧肾脏,暴露肾蒂。将双侧肾动脉用动脉夹夹持,肾脏由粉红转为紫黑提示血流阻断成功,动脉夹夹持30 min 后取下,肾脏由紫转红提示血流再灌注成功,随后逐步分层缝合伤口,背部消毒后放回鼠笼正常饮食饮水饲养。再灌注72 h后处死,取出双侧肾脏。
1.2.2 细胞缺氧复氧模型构建
取生长状态良好、处于对数生长期的人肾小管上皮细胞(HK-2),吸弃培养基,PBS清洗2 次,更换为无糖无血清的不全培养基,将细胞置于厌氧罐,随后立即放入厌氧产气袋,盖好密封盖,放进培养箱培养12 h后取出细胞,更换为DMEM完全培养基置于培养箱常态培养复氧6 h[9]。
1.2.3 实验分组与处理
体内实验中,24只雄性C57BL/6 鼠被随机分为4 组:假手术(Sham)组、IRI 组、IRI+DMSO组、IRI+SN-011组,6只/组。IRI组:按上述肾缺血再灌注模型制备方法处理;Sham组:仅游离肾蒂,不作其他特殊处理;IRI+SN-011组:手术同IRI组,在夹闭双肾动脉30 min 后,参考文献[10]方法,术后立即注射5 mg/kg的STING抑制剂SN-011,以后每24 h注射1次同样剂量SN-011,直至再灌注72 h 处死;IRI+DMSO组:手术同IRI组,在夹闭双肾动脉30 min后,每天注射与SN-011等量5%的DMSO,直至处死。
体外试验中,将HK-2细胞分为4组:对照组、H/R组、H/R+DMSO组、H/R+SN-011组。对照组:正常条件培养,不做任何处理;H/R组:处理方法同细胞缺氧模型构建;H/R+SN-011 组:H/R同时加入5 μmol/L SN-011;H/R+DMSO组:H/R同时加入与SN-011等量5%的DMSO。
1.2.4 血清肌酐及尿素氮检测
再灌注72 h后,取各组小鼠眼球血,4 ℃、8000 r/min离心10 min,取上层血清,使用全自动生化分析仪进行检测。
1.2.5 过碘酸雪夫(PAS)染色
小鼠取眼球血后行颈脱臼处死,迅速取双侧肾脏,小心去除肾包膜后PBS清洗,多聚甲醛固定3 d后,乙醇脱水,再用二甲苯透明,经石蜡包埋后进行切片,厚度5 μm,再经烤片、二甲苯脱蜡、梯度乙醇水化后放入高碘酸染液和雪夫染液中染色,再用苏木素染液染色、氨水返蓝后封片,镜检观察不同处理下肾组织结构情况。
1.2.6 免疫组织化学(IHC)染色
制备石蜡切片,再烤片、脱蜡、乙醇水化,一抗在4 ℃条件下孵育过夜,次日使用二抗孵育,加显色剂反应,再经苏木素染液复染,1%的盐酸酒精分化,随后洗涤干净后氨水返蓝,最后脱水封片,镜检观察不同处理下肾组织目的蛋白表达情况。
1.2.7 Western blotting
STING、NLRP3 兔源一抗按1∶2000比例稀释,TLR-4、KIM-1、p65、caspase-1兔源一抗按1∶1000比例稀释,山羊抗兔和抗鼠二抗按1∶2000比例稀释待用;提取肾组织蛋白并进行BCA定量后,再按80 V、30 min跑浓缩胶,110 V、60 min跑分离胶条件进行常规SDS-PAGE胶电泳,使用PVDF膜在200 mA、120 min 条件下转膜,再经5%脱脂牛奶封闭4 h 以上、4 ℃一抗孵育过夜,二抗常温孵育2 h后,洗膜曝光。采用Image J软件分析蛋白相对表达量。
1.2.8 ELISA检测
取所需量的肾脏组织块后,用预冷PBS 缓冲液冲洗。再使用超声充分匀浆组织,然后在4 ℃条件下以5000×g 的速度离心5 min。取上清液后,根据ELISA试剂盒的操作说明书,检测IL-1β、IL-6、TNF-α的分泌水平。主要步骤为制备ELISA标准品后与样品加入包被好的酶标板,静置过夜。弃去上清后,用PBST 洗涤3 次。加入生物素标记抗体,室温孵育1 h,然后弃去上清,再次用PBST洗涤3 次。加入亲和链霉素-HRP后,孵育30 min,再次洗涤3次。加入TMB溶液进行显色,终止反应后使用酶标仪在450 nm波长测定吸光度值(A450nm),绘制标准曲线以计算样品浓度。
1.2.9 流式细胞仪检测
将经过上述实验分组处理的细胞,经过无EDTA胰酶处理后,与预先收集的细胞培养上清混合来终止消化,4 ℃、800 r/min 条件下离心收集细胞后再用PBS清洗,随后同等条件下再次离心,随后使用Binding 缓冲液重悬细胞。取细胞悬液并加入Annexin V-FITC和PI染料,在避光条件下孵育10 min,然后进行上机检测。
1.2.10 生物信息学分析
从GEO数据库下载GSE98622全基因微阵列表达谱数据集,使用limma包对Sham组和IRI组进行差异分析。以Plt;0.05 和|logFC|gt;1.5 的标准对分析结果进行筛选。为进行京都基因与基因组百科全书(KEGG)富集分析,采用了\"clusterProfiler\"和\"org.Hs.eg.db\" R包。最后,采用\"ggplot2\" R包制作了热图、火山图和气泡图展示结果。
1.2.11 RT-qPCR实验
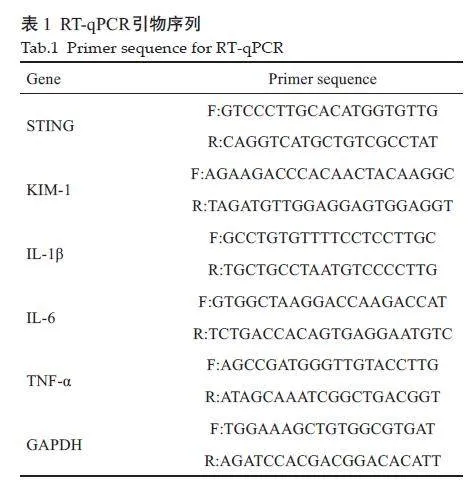
采用Trizol 法提取组织或细胞RNA后,按照Transgen反转录试剂盒说明书将RNA逆转录,再按照PerfectStart Green Qpcr SuperMix说明书配置反应混合液,按规定条件进行反应,2-ΔΔCt法(内参GAPDH)计算目的基因表达水平。引物序列见表1。
1.2.12 统计学分析
采用Graphpad Prism8.0软件进行统计学分析,符合正态分布的计量资料以均数±标准差表示,两组间比较用独立样本t检验,多组间比较用单因素方差分析,所有实验均独立重复3次,以Plt;0.05为差异有统计学意义。
2 结果
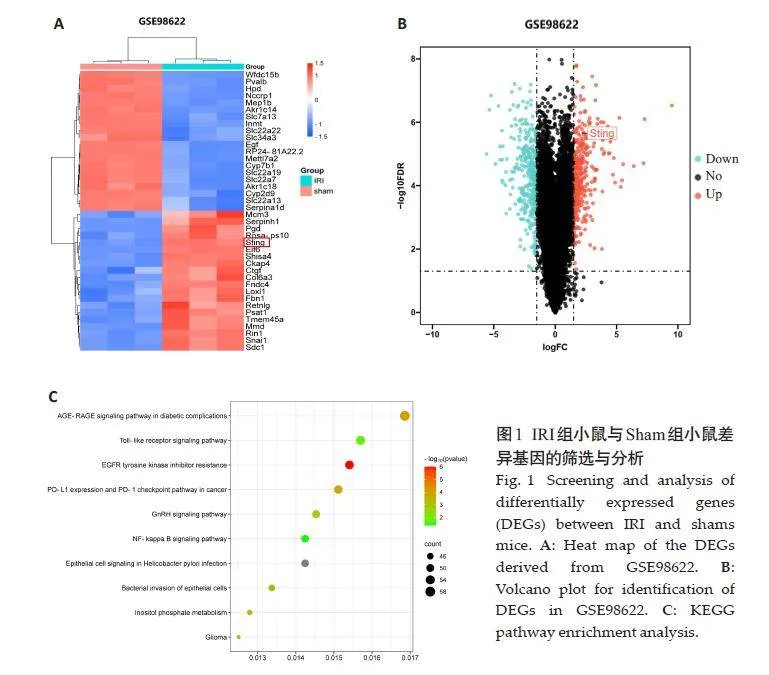
2.1 生物信息学分析
GSE98622数据集共筛选出681个差异基因,其中344 个上调基因,337 个下调基因,热图和火山图显示STING在数据集IRI 组表达水平明显上升(图1A、B)。KEGG富集分析结果显示差异基因富集于Toll样受体信号通路以及NF-κB通路(图1C)。
2.2 小鼠肾缺血再灌注模型构建效果验证
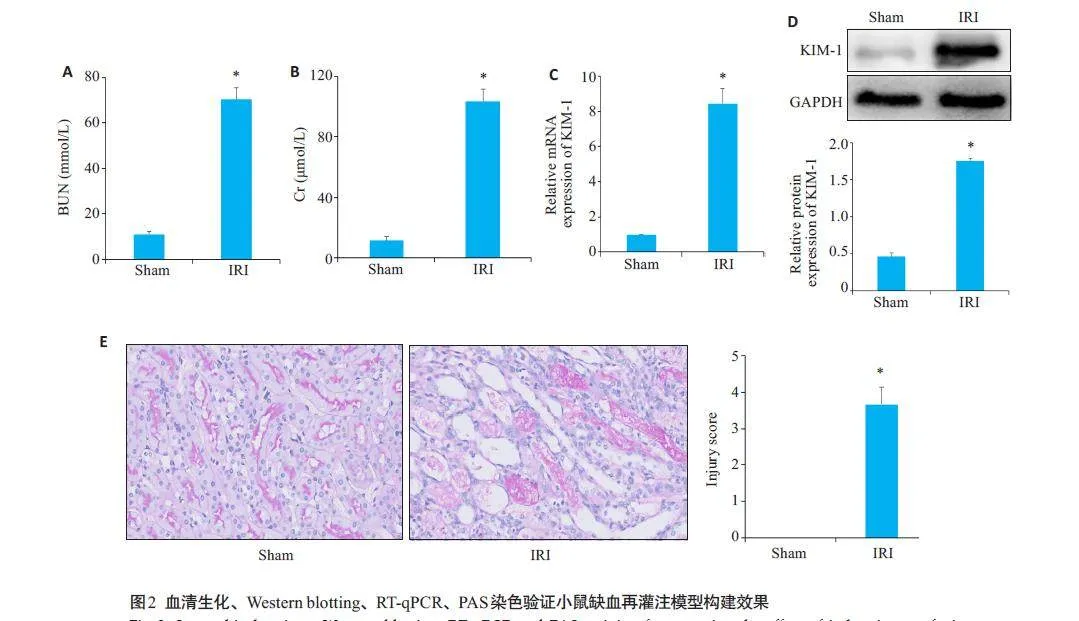
与Sham组相比,IRI组血清肌酐、尿素氮水平上升(Plt;0.05,图2A、B);RT-qPCR及Western blotting结果显示,IRI组KIM-1 mRNA和蛋白相对表达量上升(Plt;0.05,图2C、D);组织PAS染色结果显示,Sham组肾组织整体形态无明显异常,IRI组可见肾小管扩张,肾小管上皮细胞有脱落死亡、蛋白质管型增加(图2E)。与Sham组相比,IRI组肾组织损伤程度明显加重(Plt;0.05),小鼠肾缺血再灌注模型构建成功。
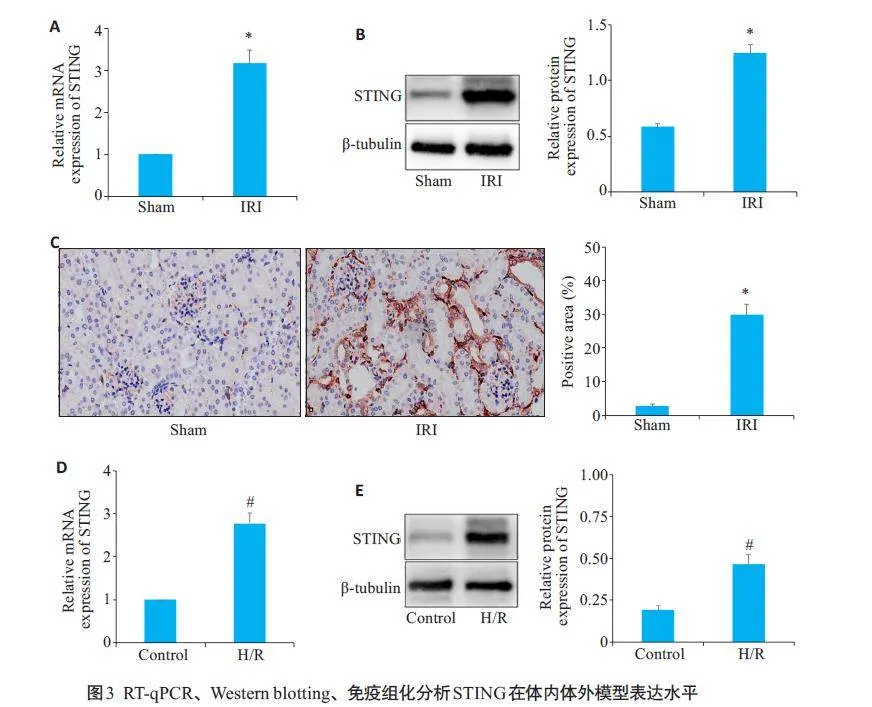
2.3 STING在体内体外肾缺血和缺氧模型中表达水平
在体内小鼠IRI模型以及体外HK-2细胞H/R模型中,RT-qPCR结果显示,与Sham组相比,IRI 组STING的mRNA相对表达量上升(Plt;0.05,图3A),与对照组相比,H/R组STING的mRNA水平上升(Plt;0.05,图3D);Western blotting 及免疫组织化学结果显示,与Sham组相比,IRI 组STING的蛋白表达量升高(Plt;0.05,图3B、C),且在免疫组织化学中,STING主要表达于肾小管上皮细胞以及肾小球基底膜。Western blotting结果显示,与对照组相比,H/R组STING的蛋白相对表达量上升(Plt;0.05,图3E)。
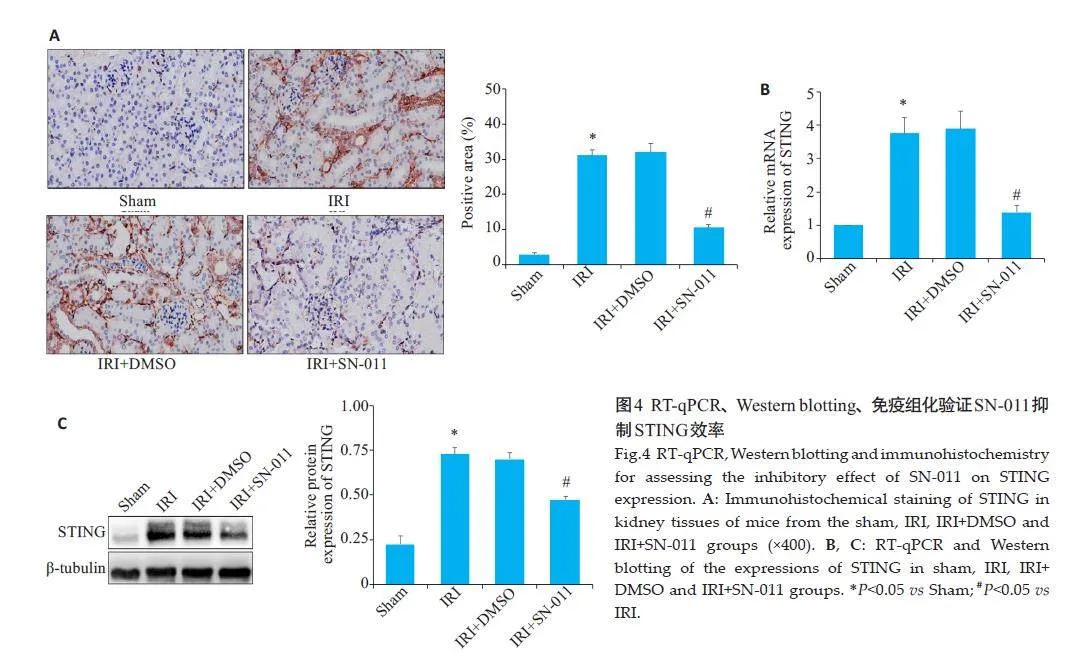
2.4 SN-011对STING表达水平抑制效率验证
RT-qPCR检测显示,IRI组与Sham组相比,肾组织中STING的mRNA水平升高(Plt;0.05);与IRI+DMSO组的差异无统计学意义(Pgt;0.05);与IRI组相比,SN-011可明显抑制STING mRNA 水平(Plt;0.05,图4B);Western blotting 和免疫组织化学检测显示,与Sham组相比,IRI 组肾组织中STING蛋白相对表达量上升(Plt;0.05);与IRI+DMSO 相比,差异无统计学意义(Pgt;0.05);与IRI组相比,IRI+SN-011组的STING 蛋白相对表达量下降(Plt;0.05,图4A、C)。
2.5 SN-011抑制STING表达后对肾组织损伤程度影响
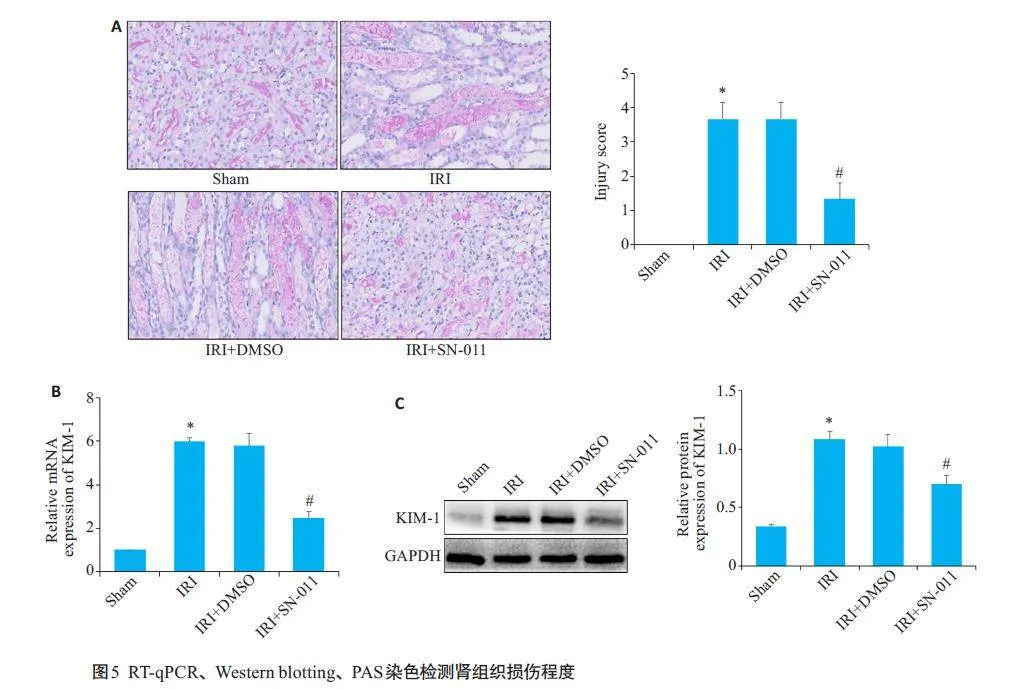
PAS染色结果显示,Sham组肾小管组织形态正常,但IRI组肾组织中可见肾小管扩张,肾小管上皮细胞坏死脱落,有管型形成;经SN-011处理后,肾组织损伤程度明显改善(图5A);RT-qPCR及Western blotting 结果显示,与Sham组相比,IRI组KIM-1 mRNA和蛋白相对表达量上升(Plt;0.05),与IRI+DMSO的差异无统计学意义(P>0.05),而与IRI组相比,IRI+SN-011组KIM-1mRNA和蛋白相对表达量下降(Plt;0.05,图5B、C)。
2.6 SN-011抑制STING表达后对肾组织炎症水平影响
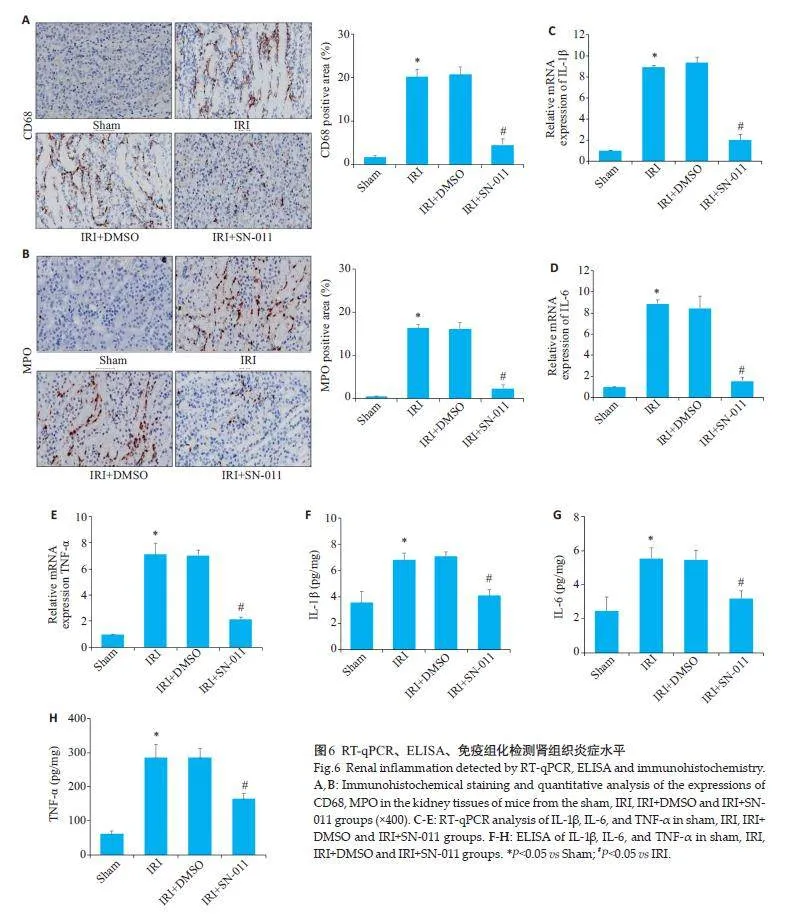
免疫组织化学检测显示,Sham组肾组织中CD68和MPO几乎不表达,而IRI 组中肾组织CD68 和MPO阳性表达增多(Plt;0.05),IRI+DMSO 组与IRI 组表达的差异无统计学意义(Pgt;0.05),经SN-011 处理后,肾组织中CD68和MPO阳性表达下降(Plt;0.05,图6A、B);RT-qPCR 以及ELISA 结果显示,与Sham组相比,IRI组肾组织中IL-1β、IL-6 和TNF-α炎性因子水平上升(Plt;0.05),与IRI+DMSO组的差异无统计学意义(Pgt;0.05),与IRI组相比,IRI+SN-011组肾组织中IL-1β、IL-6和TNF-α水平降低(Plt;0.05,图6C~H)。
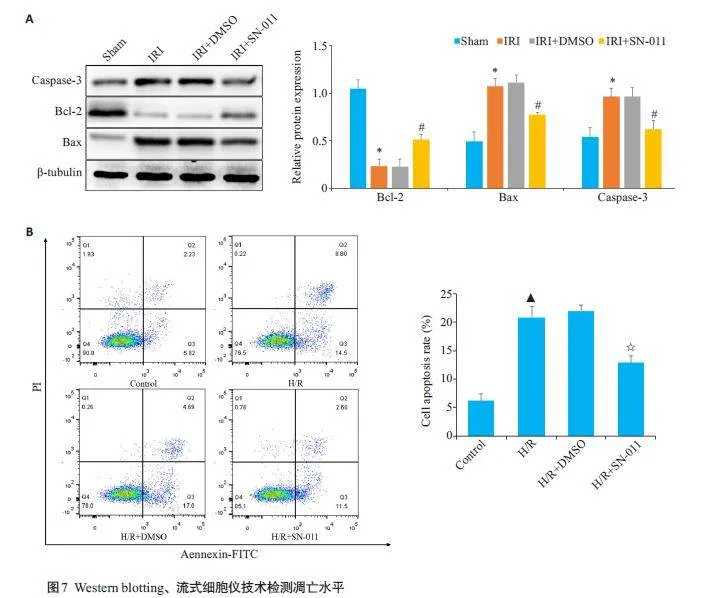
2.7 SN-011抑制STING表达后对凋亡水平影响
Western blotting检测显示,与Sham组相比,IRI组Bax、caspase-3蛋白相对表达量上升(Plt;0.05),Bcl-2蛋白相对表达量下降(Plt;0.05),与IRI+DMSO组的差异无统计学意义(Pgt;0.05);与IRI组相比,IRI+SN-011组Bax、caspase-3蛋白相对表达量下降(Plt;0.05),Bcl-2蛋白相对表达量上升(Plt;0.05,图7A)。流式细胞仪检测各组细胞凋亡率结果显示,H/R组凋亡率高于对照组(Plt;0.05),而与H/R+DMSO组的差异无统计学意义(Pgt;0.05),加入SN-011后的细胞凋亡率又下降(Plt;0.05,图7B)。
2.8 SN-011 抑制STING 表达后对TLR4/NF- κB 及NLRP3通路影响
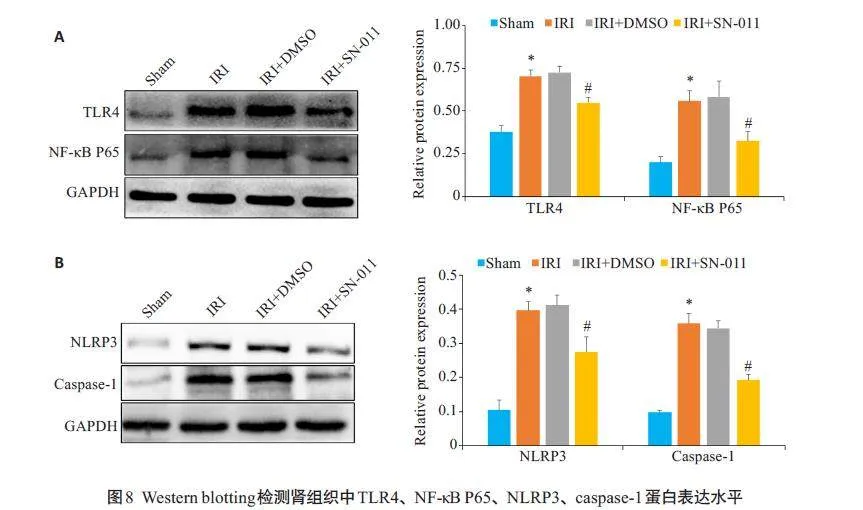
Western blotting检测显示,与Sham组相比,IRI组TLR4、p65 蛋白相对表达量上升(Plt;0.05),且NLRP3、Caspase-1 蛋白相对表达量上升(Plt;0.05),和IRI+DMSO组的差异无统计学意义(Pgt;0.05);经SN-011 处理后,TLR4、p65、NLRP3、Caspase-1 蛋白表达被抑制(Plt;0.05,图8)。
3 讨论
RIRI存在于各种器官移植、休克复苏之中,是导致AKI 主要原因之一,对患者危害巨大[11, 12]。目前由于RIRI病理生理机制复杂,涉及氧化应激[13]、免疫反应[14]、细胞凋亡与修复[15]等各个方面,且分子机制尚未完全阐述清楚。既往诸多研究致力于研究各种分子靶点对RIRI影响,如G蛋白偶联受体激酶4可与信号传导转录激活因子结合,引发坏死性凋亡,加重RIRI,其下调可能为肾脏保护提供潜在的治疗策略[16];人类尿液衍生干细胞外泌体来源的miR-146a-5p为其无创治疗RIRI提供一种新思路[17];甲基-CpG结合蛋白可减轻RIRI中肾细胞死亡、炎症和纤维化的影响,发挥肾脏保护分子的作用[18]。本研究通过验证肾损伤功能指标以及重要标志物KIM-1[19],在成功构建RIRI 模型的基础上,发现STING在小鼠RIRI模型表达水平异常增高,和细胞缺氧模型中变化水平一致,这符合前期生物信息学分析结果,提示其或许可能作为一种与以往不同的致病分子靶点,在肾IRI模型中发挥相关作用。
研究提示,STING作为免疫系统的关键组分,在诸如系统性红斑狼疮、结肠炎以及前列腺癌等多种自身免疫性疾病、炎症性疾病以及肿瘤中扮演重要角色[20-23]。也有研究表明,其作用机制通常与促进一型干扰素以及其他炎症相关因子或细胞产生、调节机体细胞凋亡或细胞焦亡等过程相关[24]。而RIRI 中,涉及多种炎性细胞和炎性因子的炎症级联反应,以及内质网应激、DNA损伤等多种途径介导的细胞凋亡同样起到重大作用[25-29]。本研究构建RIRI模型并抑制STING表达后,RT-qPCR以及ELISA检测IL-1β、IL-6、TNF-α炎性因子分泌水平变化,并且通过Western blotting和流式技术检测凋亡水平,结果与推测相符,STING在RIRI中同样可以促进肾组织中炎症与凋亡水平,这与在其他疾病中发挥的作用相似。此外,为了进一步明确STING对炎性细胞产生与成熟的影响,本研究同时还通过免疫组织化学检测发现STING可影响IRI 肾组织中CD68、MPO水平,从另一方面再次说明STING影响炎性细胞的产生与成熟在RIRI炎症反应中发挥作用。
既往对STING 作用的研究中,常选用C-176、H-151 等STING抑制剂来抑制STING表达与激活,探讨相关研究机制[30-33]。而作为一种新型且有效的STING抑制剂,本研究中的SN-011可与环二核苷酸(CDN)竞争性结合STING二聚体,阻断CDN结合和STING激活,具有更好的特异性与安全性,以及更高的抑制效率[34]。与既往研究相比,本研究构建小鼠IRI模型注射SN-011 后,STING的mRNA和蛋白水平均下降,较低的抑制剂剂量在体内小鼠模型中即具有更好的抑制效率,也验证了SN-011 的高效性,这可能与SN-011 对CDN结合更高的亲和性以及与既往不同的针对STING二聚体形成的特殊机制相关。抑制STING表达后,与IRI组相比,IRI+SN-011组PAS染色显示组织结构损伤减轻,KIM-1在mRNA和蛋白水平上均下降,提示肾损伤程度有效减轻,推测SN-011除在包括STING相关婴儿期起病血管病在内等多种自身免疫性疾病与炎症性疾病中发挥作用外[35-38],或许在RIRI相关治疗研究领域中也有一定前景。
TLR是一种横跨细胞膜的模式识别受体,可以通过髓样分化因子88途径激活TLR4/NF-κB信号通路[39]。而NLRP3炎性小体复合物由NLRP3、凋亡相关斑点样蛋白和caspase-1 组成,可感知外源性微生物或内源危险信号,亦可在炎症调节中发挥重要作用。既往研究认为,RIRI 是一种涉及炎症性反应以及肾小管细胞死亡机制的疾病,TLR4/NF-κB以及NLRP3 均可在RIRI扮演重要角色[40-44]。本研究通过KEGG富集分析,以及检测RIRI模型中相关通路标志蛋白表达,也验证了这一结果正确性。但前述结果中已发现STING可影响炎症与凋亡反应促进RIRI,具体机制尚不明确。STING可与多条经典信号通路产生交互作用[7, 45]。为了揭示STING影响炎症与凋亡促进RIRI的调控机制,本研究使用STING抑制剂抑制STING激活后,明显降低了TLR4/NF-κB/NLRP3 通路标志蛋白相对表达量,这提示STING调节TLR4/NF-κB/NLRP3 通路或许是影响RIRI的潜在机制。
综上所述,本实验证实了STING在RIRI中表达水平上升,且可通过促进炎症、凋亡以及影响TLR4/NF-κB/NLRP3 通路加重RIRI。SN-011 作为一种新型的STING抑制剂,除了在自身免疫性疾病中发挥重要作用外,或许也可通过抑制STING表达,从而减轻炎症与凋亡反应,在缓解肾缺血再灌注中发挥作用,具有一定治疗潜力。本研究可为STING在RIRI 相关作用机制的深入探究提供借鉴意义,对未来寻找RIRI治疗相关分子靶点以及治疗药物提供重要帮助。
参考文献:
[1] Hoste EAJ, Kellum JA, Selby NM, et al. Global epidemiology andoutcomes of acute kidney injury[J]. Nat Rev Nephrol, 2018, 14(10):607-25.
[2] Zuk A, Bonventre JV. Acute kidney injury[J]. Annu Rev Med, 2016,67: 293-307.
[3] Arai S, Kitada K, Yamazaki T, et al. Apoptosis inhibitor ofmacrophage protein enhances intraluminal debris clearance andameliorates acute kidney injury in mice[J]. Nat Med, 2016, 22(2):183-93.
[4] Inagi R, Ishimoto Y, Nangaku M. Proteostasis in endoplasmicreticulum: new mechanisms in kidney disease[J]. Nat Rev Nephrol,2014, 10(7): 369-78.
[5] Yan MJ, Tang CY, Ma ZW, et al. DNA damage response innephrotoxic and ischemic kidney injury[J]. Toxicol ApplPharmacol, 2016, 313: 104-8.
[6] Malek M, Nematbakhsh M. Renal ischemia/reperfusion injury; frompathophysiology to treatment[J]. J Renal Inj Prev, 2015, 4(2): 20-7.
[7] Fang R, Wang CG, Jiang QF, et al. NEMO-IKKβ are essential forIRF3 and NF- κB activation in the cGAS-STING pathway[J]. JImmunol, 2017, 199(9): 3222-33.
[8] Fang R, Jiang QF, Guan YK, et al. Golgi apparatus-synthesizedsulfated glycosaminoglycans mediate polymerization and activationof the cGAMP sensor STING[J]. Immunity, 2021, 54(5): 962-75.e8.
[9] Ren P, Cao JL, Lin PL, et al. Molecular mechanism of luteolinregulating lipoxygenase pathway against oxygen-glucosedeprivation/reperfusion injury in H9c2 cardiomyocytes based onmolecular docking[J]. Zhongguo Zhong Yao Za Zhi, 2021, 46(21):5665-73.
[10]Bi R, Yang YL, Liao HW, et al. Porphyromonas gingivalis inducesan inflammatory response via the cGAS-STING signaling pathwayin a periodontitis mouse model[J]. Front Microbiol, 2023, 14:1183415.
[11] Pressly JD, Park F. DNA repair in ischemic acute kidney injury[J].Am J Physiol Renal Physiol, 2017, 312(4): F551-5.
[12]Hu HL, Zou C. Mesenchymal stem cells in renal ischemiareperfusioninjury: biological and therapeutic perspectives[J]. CurrStem Cell Res Ther, 2017, 12(3): 183-7.
[13] Inagi R. Endoplasmic reticulum stress in the kidney as a novelmediator of kidney injury[J]. Nephron Exp Nephrol, 2009, 112(1):e1-9.
[14]Cao Q, Wang YP, Niu ZG, et al. Potentiating tissue-resident type 2 innate lymphoid cells by IL-33 to prevent renal ischemiareperfusioninjury[J]. J Am Soc Nephrol, 2018, 29(3): 961-76.
[15]Havasi A, Borkan SC. Apoptosis and acute kidney injury[J]. KidneyInt, 2011, 80(1): 29-40.
[16]Yang DH, Tang M, Zhang MM, et al. Downregulation of G proteincoupledreceptor kinase 4 protects against kidney ischemiareperfusioninjury[J]. Kidney Int, 2023, 103(4): 719-34.
[17]Li XR, Liao J, Su XJ, et al. Human urine-derived stem cells protectagainst renal ischemia/reperfusion injury in a rat model viaexosomal miR-146a-5p which targets IRAK1[J]. Theranostics,2020, 10(21): 9561-78.
[18]Wang J, Xiong MR, Fan Y, et al. Mecp2 protects kidney fromischemia-reperfusion injury through transcriptional repressing IL-6/STAT3 signaling[J]. Theranostics, 2022, 12(8): 3896-910.
[19] van Timmeren MM, van den Heuvel MC, Bailly V, et al. Tubularkidney injury molecule-1 (KIM-1) in human renal disease[J]. JPathol, 2007, 212(2): 209-17.
[20]Gkirtzimanaki K, Kabrani E, Nikoleri D, et al. IFNα impairsautophagic degradation of mtDNA promoting autoreactivity of SLEmonocytes in a STING-dependent fashion[J]. Cell Rep, 2018, 25(4): 921-33.e5.
[21]Gao YP, Zhang NN, Zeng ZH, et al. LncRNA PCAT1 activatesSOX2 and suppresses radioimmune responses via regulating cGAS/STING signalling in non-small cell lung cancer[J]. Clin TranslMed, 2022, 12(4): e792.
[22]Li X, Liu YJ, Wang Y, et al. Epoxy triglyceride enhances intestinalpermeability via caspase-1/NLRP3/GSDMD and cGAS-STINGpathways in dextran sulfate sodium-induced colitis mice[J]. J AgricFood Chem, 2023, 71(10): 4371-81.
[23]Wu JJ, Zhao L, Hu HG, et al. Agonists and inhibitors of the STINGpathway: potential agents for immunotherapy[J]. Med Res Rev,2020, 40(3): 1117-41.
[24]Barber GN. STING: infection, inflammation and cancer[J]. Nat RevImmunol, 2015, 15(12): 760-70.
[25]Lu L, Zhou HM, Ni M, et al. Innate immune regulations and liverischemia-reperfusion injury[J]. Transplantation, 2016, 100(12):2601-10.
[26]DeWolf SE, Kasimsetty SG, Hawkes AA, et al. DAMPs releasedfrom injured renal tubular epithelial cells activate innate immunesignals in healthy renal tubular epithelial cells[J]. Transplantation,2022, 106(8): 1589-99.
[27]Raup-Konsavage WM, Wang YM, Wang WW, et al. Neutrophilpeptidyl arginine deiminase-4 has a pivotal role in ischemia/reperfusion-induced acute kidney injury[J]. Kidney Int, 2018, 93(2): 365-74.
[28]Salvadori M, Rosso G, Bertoni E. Update on ischemia-reperfusioninjury in kidney transplantation: Pathogenesis and treatment[J].World J Transplant, 2015, 5(2): 52-67.
[29]Liu CH, Wang QD, Niu L. Sufentanil inhibits Pin1 to attenuate renaltubular epithelial cell ischemia-reperfusion injury by activating thePI3K/AKT/FOXO1 pathway[J]. Int Urol Nephrol, 2023, 55(8):1903-16.
[30]Wu B, Xu MM, Fan C, et al. STING inhibitor ameliorates LPSinducedALI by preventing vascular endothelial cells-mediatedimmune cells chemotaxis and adhesion[J]. Acta Pharmacol Sin,2022, 43(8): 2055-66.
[31]Liu R, Li JY, Shao JC, et al. Innate immune response orchestratesphosphoribosyl pyrophosphate synthetases to support DNA repair[J]. Cell Metab, 2021, 33(10): 2076-89.e9.
[32]Yang B, Li X, Fu Y, et al. MEK inhibition remodels the immunelandscape of mutant KRAS tumors to overcome resistance to PARPand immune checkpoint inhibitors[J]. Cancer Res, 2021, 81(10):2714-29.
[33]Zhang YN, Dong YL, Hao WP, et al. Increased cGAS/STINGsignaling components in patients with Mooren's ulcer[J]. Int JOphthalmol, 2021, 14(11): 1660-5.
[34]Hong Z, Mei JH, Li CH, et al. STING inhibitors target the cyclicdinucleotide binding pocket[J]. Proc Natl Acad Sci U S A, 2021, 118(24): e2105465118.
[35]Diao FF, Bai J, Jiang CL, et al. The papain-like protease of porcinereproductive and respiratory syndrome virus impedes STINGtranslocation from the endoplasmic reticulum to the Golgi apparatusby deubiquitinating STIM1[J]. J Virol, 2023, 97(4): e0018823.
[36]Yang BX, Xie XR, Wu ZY, et al. DNA damage-mediated cellularsenescence promotes hand-foot syndrome that can be relieved bythymidine prodrug[J]. Genes Dis, 2022, 10(6): 2557-71.
[37]Ablasser A, Chen ZJ. cGAS in action: expanding roles in immunityand inflammation[J]. Science, 2019, 363(6431): eaat8657.
[38]Gulen MF, Koch U, Haag SM, et al. Signalling strength determinesproapoptotic functions of STING[J]. Nat Commun, 2017, 8(1): 427.
[39]Lehnardt S, Massillon L, Follett P, et al. Activation of innateimmunity in the CNS triggers neurodegeneration through a Toll-likereceptor 4-dependent pathway[J]. Proc Natl Acad Sci USA, 2003,100(14): 8514-9.
[40]Wang L, Yang JW, Lin LT, et al. Acupuncture attenuatesinflammation in microglia of vascular dementia rats by inhibitingmiR-93-mediated TLR4/MyD88/NF‑κB signaling pathway[J].Oxid Med Cell Longev, 2020, 2020: 8253904.
[41]Zhang NX, Guan C, Liu ZY, et al. Calycosin attenuates renalischemia/reperfusion injury by suppressing NF‑κB mediatedinflammation via PPARγ/EGR1 pathway[J]. Front Pharmacol,2022, 13: 970616.
[42]Alaaeldin R, Bakkar SM, Mohyeldin RH, et al. Azilsartan modulatesHMGB1/NF- κB/p38/ERK1/2/JNK and apoptosis pathways duringrenal ischemia reperfusion injury[J]. Cells, 2023, 12(1): 185.
[43]Ding HS, Huang Y, Qu JF, et al. Panaxynol ameliorates cardiacischemia/reperfusion injury by suppressing NLRP3-inducedpyroptosis and apoptosis via HMGB1/TLR4/NF- κB axis[J]. IntImmunopharmacol, 2023, 121: 110222.
[44]Liu YY, Lei ZL, Chai H, et al. Salidroside alleviates hepaticischemia-reperfusion injury during liver transplant in rat throughregulating TLR-4/NF‑κB/NLRP3 inflammatory pathway[J]. SciRep, 2022, 12(1): 13973.
[45]Li N, Zhou H, Wu HM, et al. STING-IRF3 contributes tolipopolysaccharide-induced cardiac dysfunction, inflammation,apoptosis and pyroptosis by activating NLRP3[J]. Redox Biol,2019, 24: 101215.
(编辑:郎 朗)
基金项目:安徽省自然科学基金重点项目(2008085QH358);慢性疾病免疫学基础与临床安徽省重点实验室开放课题基金(AHIAI2022K01)
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21 02:14:52
感染、炎症、修复(2021年1期)2021-07-28 06:17:48
学苑创造·A版(2020年9期)2020-10-13 09:41:02
小学生学习指导(低年级)(2017年10期)2017-10-10 01:00:05
感染、炎症、修复(2016年4期)2016-04-17 05:32:05
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:55
中国病理生理杂志(2015年8期)2015-12-21 12:38:06
医学研究杂志(2015年3期)2015-06-10 06:41:52
西南军医(2015年3期)2015-04-23 07:28:32
创业家(2015年1期)2015-02-27 07:52:02
