
肿瘤相关成纤维细胞上调hsa-miR-18b-5p 靶向FBXL3 促进前列腺癌的增殖及转移
2024-11-03 00:00:00骆金光陶怀祥闻志远陈龙胡昊关翰
南方医科大学学报 2024年7期





摘要:目的 探讨受肿瘤相关成纤维细胞(CAFs)调控的hsa-miR-18b-5p在前列腺癌(PCa)中的表达水平及作用,并研究对其在PCa 发生发展过程中的分子机制。方法 利用生物信息学技术分析在PCa 中高表达的miRNA,构建肿瘤相关成纤维细胞并与PCa细胞系共培养,验证CAFs对PCa细胞增殖和迁移的影响及对hsa-miR-18b-5p的调控作用;RT-qPCR验证20例患者癌组织及癌旁正常组织、前列腺正常上皮增生细胞及PCa 各细胞系中hsa-miR-18b-5p 的表达水平;通过脂质体法将阴性对照及hsamiR-18b-5p 抑制物分别转染入PCa 细胞C4-2、LNCAP中,分为NC inhibitor 组和hsa-miR-18b-5p inhibitor 组;采用细胞集落形成、CCK-8 实验、划痕愈合、Transwell、IC50实验、流式细胞术分别检测C4-2、LNCAP细胞增殖、迁移、侵袭、耐药能力、凋亡和周期;建立PCa裸鼠移植瘤模型,定期测量移植瘤的质量和体积,Kaplan-Meier生存曲线分析裸鼠生存情况;利用靶基因预测分析网站(Targetscan、Mirtarbase、miRDB、miRDIP)预测has-miR-18b-5p 的靶基因,并通过双荧光素酶报告基因分析验证hsa-miR-18b-5p与靶基因的靶向关系,RT-qPCR、Western blotting检测各组细胞靶基因的表达水平。结果 生物信息学分析结果显示,在PCa 中高表达的miRNA有17 个,结合差异表达程度以及相关文献筛选出拟研究的基因:miR-148a、miR-17、miR-18b-5p、miR-770、miR-297-3p。CAFs与PCa细胞系共培养后hsa-miR-18b-5p的表达水平升高(Plt;0.01),且促进PCa细胞的增殖与迁移(Plt;0.01),敲除has-miR-18b-5p可抵消CAFs对PCa细胞增殖及迁移的影响,确定has-miR-18b-5p为研究基因。与正常前列腺上皮细胞或肿瘤旁组织相比,PCa细胞或肿瘤组织中hsa-miR-18b-5p的表达升高(Plt;0.05);敲除hsa-miR-18b-5p可抑制C4-2、LNCAP细胞的增殖、迁移、侵袭以及耐药(Plt;0.05);敲除hsa-miR-18b-5p可抑制裸鼠移植瘤质量和体积的增长,增加裸鼠的生存时间(Plt;0.05)。靶基因预测分析网站显示,FBXL3 是hsa-miR-18b-5p 的一个潜在作用靶点,双荧光素酶报告基因证实has-miR-18b-5p 与FBXL3基因间存在结合位点,且敲除hsa-miR-18b-5p可增加C4-2、LNCAP细胞中FBXL3的蛋白表达(Plt;0.05)。结论 CAFs上调前列腺癌细胞hsa-miR-18b-5p的表达水平,后者可能通过靶向调控FBXL3基因的表达促进PCa细胞的增殖和转移。
关键词:hsa-miR-18b-5p;肿瘤相关成纤维细胞;FBXL3;前列腺癌;促癌
在男性人群中,前列腺癌(PCa)是仅次于肺癌的最常见的恶性肿瘤[1, 2],也是全球第5大致死性疾病[3]。据统计,2020年泌尿系统癌症占全球1930万新发癌症的13.1%[4],其中仅PCa 就占美国男性新诊断癌症的21%[5]。通常,PCa 的治疗方式主要包括放射治疗、手术、激素治疗和冷冻手术等[6],然而,这些方法经常与各种不良副作用相关[7]。尽管对PCa的系统化以及个体化治疗研究已取得巨大进展[8-10],但PCa 患者的预后仍不稳定[11]。因此,明确PCa肿瘤的发生、发展以及转移的原因和分子机制对于肿瘤的早期发现、早期诊断和个体化治疗具有重要意义。
microRNA(miRNAs)是一类来源于内源性转录本含有18~22 个核苷酸的非编码RNA,它们可以通过转录后基因沉默促进基因组的调控[12]。通常,这些短RNA能够结合到其靶信使RNA(mRNA)的3'非翻译区的特定位点,并通过完全或不完全碱基配对介导mRNA降解或阻断基因翻译[13, 14],超过60%的哺乳动物mRNA是miRNA的靶点[15]。因此,miRNA可调节各种细胞过程,例如存活、生长和分化[16]。具体而言,miRNA可以作为肿瘤抑制因子或致癌基因,其参与人类癌症的发展[17, 18]。有研究证明,miRNAs在肺癌、胰腺癌、卵巢癌、胃肠癌、PCa 等多种癌症的发生发展中发挥着重要作用[19-23]。
CAFs是肿瘤微环境的重要组成部分,特别是在基质细胞中[24],可以构建和重塑细胞外基质,并且与肿瘤发生、肿瘤转移、肿瘤预后和抗肿瘤免疫应答密切相关[25]。在肿瘤微环境中,成纤维细胞调节肿瘤细胞的血管生成和转移,并且miRNA是CAFs 对肿瘤细胞起促进功能的关键调节剂[26]。据报道,在永生化成纤维细胞中诱导miR-21过表达可导致α-平滑肌肌动蛋白的表达上调,表明miR-21过表达驱动成纤维细胞向肌成纤维细胞转化并且miR-21过表达的成纤维细胞可保护结直肠癌细胞免受奥沙利铂诱导的凋亡,并增加其增殖能力[27];miR-205通过阻断肿瘤细胞和相关成纤维细胞之间回路的传入和传出,从而中断由反应性基质参与的促氧化和促炎回路,作为针对PCa转移的制动器[28]。
hsa-miR-18b-5p在肿瘤发生中起重要作用,包括乳腺癌、结直肠癌、胆囊癌、卵巢癌以及肺腺癌等。例如,has-miR-18b-5p 可以通过与转录延伸因子A 样7 的3'UTR 特异性结合,通过激活核因子-κB促进核Snail异位激活,从而诱导上皮间质转化并促进肿瘤细胞的侵袭和转移[29];非编码RNA LOXL1-AS1通过调节hsa-miR-18b-5p/VMA21轴在卵巢癌中表现出致癌活性[30];长的非编码RNA AC073284.4 通过在耐紫杉醇的乳腺癌细胞中调控hsa-miR-18b-5p来抑制上皮-间质转化[31]。然而,hsa-miR-18b-5p在PCa进展中的潜在作用国内外尚未有相关报道。因此,本研究旨在探讨hsa-miR-18b-5p与CAFs 之间的调控关系以及hsa-miR-18b-5p 在PCa细胞生长和转移中的作用及其可能的分子机制,为临床治疗PCa提供潜在的新靶点。
1 材料和方法
1.1 材料
1.1.1 实验动物
雄性SPF级裸鼠30只(杭州子源实验动物科技有限公司),6~8周龄,体质量20~24 g,生产许可证号:SCXK(浙)2019-0004。饲养条件:温度23±2 ℃,相对湿度(55±5)%,12 h 昼夜交替,自由饮食进水条件下,饲养1周进行实验。本研究所有动物实验经蚌埠医科大学第一附属医院伦理委员会批准通过(伦理批号:伦动科批字[2022]第 066 号)。
1.1.2 主要试剂与仪器
人PCa细胞株(中国科学院(上海)典型培养物保藏委员会); NC inhibitor 及 hsa-miR-18b-5p inhibitor、F-框亮氨酸丰富重复蛋白3(FBXL3)双荧光素酶报告载体(上海吉玛基因);Lipofectamine2000(Invitrogen);兔抗人Vimentin 抗体、鼠抗人α-SMA抗体、兔抗人FBXL3 抗体(武汉三鹰生物技术有限公司);Dual-Glo RLuciferase Assay System试剂盒(Promega);PCR逆转录试剂盒、PerfectStart GreenQpcr SuperMix(Transgen Biotech);蛋白定量试剂盒、PAGE 凝胶快速制备试剂盒(雅酶);垂直电泳仪、曝光仪(Bio-Rad);荧光定量 PCR仪(Roche)。
1.2 方法
1.2.1 数据收集及分析
使用GEO数据库(http://www.ncbi.nlm.nih.gov/geo)获取PCa肿瘤组织样本和癌旁组织样本的差异表达的miRNA,使用R语言相应分析包对获取的数据进行分析,筛选差异基因的标准为:|log2Fld changez|≥1.0,同时Plt;0.05。
1.2.2 差异miRNA靶基因的预测及mRNA的确定
使用miRTarBase(http://mirtarbase.mbc.nctu.edu.tw/php/index. php)、TargetScan (https://www. targetscan. org/vert_80/)、miRDIP (http://ophid. utoronto. ca/mirDIP/index.jsp)、和miRDB(https://mirdb.org/index.html)等数据库预测差异miRNA的靶基因,并将4 个数据库的靶基因取交集,结合前期研究,确定后续研究的mRNA。
1.2.3 CAFs 与NFs 的构建与共培养
从临床PCa 根治性切除的标本中获取癌组织进行实验,前列腺增生标本来自耻骨上前列腺剜除或前列腺电切的患者组织。将组织块无菌条件下剪碎至离心管中,PBS清洗后置于超声波细胞破碎仪进行破碎后加入胶原酶于37 ℃恒温下消化15 min。将细胞放入培养瓶中培养,待细胞长满后胰酶消化细胞,贴壁15 min 以纯化成纤维细胞,利用Transwell小室间接共培养CAFs细胞与PCa细胞48 h。
1.2.4 组织采集
纳入2020~2023年在蚌埠医科大学第一附属医院泌尿外科为PCa 患者行手术时获得的PCa患者的肿瘤组织及癌旁非癌组织(正常组织)。所有肿瘤和正常组织样本在使用前均经过病理学确认。试验已获得蚌埠医科大学第一附属医院伦理委员会批准(批准号:2020-029)。
1.2.5 细胞培养
人PCa C4-2、LNCAP、PC3、22RV1、DU145 细胞使用含10%胎牛血清的 RPMI 1640 培养基,人BPH-1 细胞使用含10%胎牛血清的KSFM培养基,将细胞置于37 ℃、5%CO2的培养箱中培养。
1.2.6 细胞转染
取对数生长期的C4-2、LNCAP分别细胞接种于6 孔板,待细胞融合度达到50% 时,按照Lipofectamine 2000转染试剂盒说明书,分别取5 μL浓度为20 μmol/L hsa-miR-18b-5p inhibitor(hsa-miR-18b-5p inhibitor 组)和NC inhibitor(NC inhibitor 组)溶于250 μL Opti-MEM™ I 减血清培养基中;将 5 μLLipofectamine 2000 溶于250 μL Opti-MEM™ I 减血清培养基中,室温孵育5 min。分别将hsa-miR-18b-5pinhibitor 和NC inhibitor 与Lipofectamine 2000 混合后室温静置15 min。6 孔板培养基换成低血清培养基(1%),转染24 h后更换含血清培养液,继续培养48 h后进行实验。
1.2.7 RT-qPCR法
Trizol 法提取组织或细胞RNA后,按照 Transgen反转录试剂盒说明书将RNA逆转录,再按照 PerfectStart Green Qpcr SuperMix 说明书配置反应混合液,按规定条件进行反应,2-ΔΔCt 法(内参U6、GAPDH)计算目的基因表达水平,引物设计见表1。
1.2.8 Western blotting 检测
分别收集转染后的C4-2和LNCAP细胞,加入RIPA细胞裂解液(含混合蛋白酶抑制剂)裂解细胞,离心后提取总蛋白并经BCA法测定蛋白浓度;将蛋白与上样缓冲液混匀后放入100 ℃金属浴10 min灭活,SDSPAGE分离胶电泳后转至PVDF膜上;5%脱脂奶粉封闭2 h,清洗后加入一抗于4 ℃冰箱孵育过夜;加入二抗,室温杂交1 h 后于曝光仪上进行曝光。
1.2.9 CCK8检测
将转染后的C4-2和LNCAP细胞接种于96孔板,每组3个复孔,每孔培养液总量为100 μL。严格按照试剂盒说明书步骤操作,用酶标仪测定每孔吸光度值A450nm,实验重复3次。
1.2.10 细胞集落形成实验
细胞集落形成:将转染后的C4-2和LNCAP细胞按每组3个复孔、1000/孔接种于6孔板中,放入CO2培养箱培养。培养1~2 周后,肉眼可见集落形成时弃除培养液,加入1 mL/孔多聚甲醛固定30 min后弃除,加入1 mL0.1%结晶紫染色10 min,清洗后光镜下计数集落,实验重复3次。
1.2.11 细胞划痕实验
将各组C4-2 和LNCAP细胞培养于6孔板中,用高压灭菌的200 μL枪头在孔内轻轻划3道痕,用PBS清洗划下的细胞,无血清培养基培养24 h后在显微镜下拍照,计算相对划痕宽度。实验重复3次。
1.2.12 Transwell 实验
迁移实验:在Transwell 小室的下室加入600 μL完全培养基,取100 μL无血清的细胞悬液种植于Transwell小室的上室,每组3个复孔。将上室置于下室中,细胞培养箱中培养8~14 h后取出上室,擦去上室表面的细胞,用95%甲醇固定细胞20 min,用0.2%结晶紫染色30 min,显微镜下计数,实验重复3次。侵袭实验:在Transwell小室的下室加入600 μL完全培养基,取100 μL无血清的细胞悬液种植于Transwell小室的上室,每组3个复孔。将小室放在细胞培养箱中培养18~24 h后取出上室,擦去上室表面的细胞和基质胶,用95%甲醇固定细胞20 min,再用0.2%结晶紫染色30 min,显微镜下计数,实验重复3次。
1.2.13 IC50实验
将转染后的各组细胞消化离心沉淀后使用血球计数板计数细胞,将细胞以2000个/孔接种于96孔板中,待其贴壁后, 分别加入恩杂鲁胺使其终浓度达到0、2、4、8、16、32、64 μmol/L,每组设5个复孔,每孔终体积为100 μL。在每孔加入CCK-8试剂10 μL,浓度为5 mg/mL,将孔板放入37 ℃培养箱孵育1 h后用酶标仪检测每孔的OD值,吸收波长450 nm,计算增殖抑制率=[(对照组OD-实验组OD)/对照组OD]×100%,绘制IC50曲线。
1.2.14 流式细胞术检测
凋亡:将转染好的细胞消化、离心,使用1xbinding Buffer 100 mL重悬细胞沉淀,转移至1.5 mL离心管中,加入5 μL PI染液,染色3 min后加入5 μL AnnexinV-FITC染液共同染色5 min,染色结束后加400 μL的1xbinding Buffer 终止染色,使用流式细胞仪上机检测。周期:将转染好的细胞消化、离心,加入1 mL预冷的70%乙醇中,轻轻吹打混匀,4 ℃固定24 h。1000 g离心5 min沉淀细胞后去上清,加入预冷的PBS重悬细胞,离心后去上清。每管细胞样品中加入0.5 mL碘化丙啶染色液,37 ℃避光孵育30 min,使用流式细胞仪上机检测。
1.2.15 动物实验
分别将干扰has-miR-18b-5p 的慢病毒质粒LV-anti-has-miR-18b-5p(LV-anti-has-miR-18b-5p组)及其对照质粒LV-vector(LV-vector组)按病毒感染复数MOI=20感染的C4-2细胞(1.0×106)皮下注射裸鼠的左侧下腹部。每组15只小鼠,5只小鼠用于测量肿瘤的体积和质量,10只小鼠用于生存分析。5周后通过颈椎脱位法对小鼠实施安乐死,并称量肿瘤组织的质量和计算肿瘤体积。
1.2.16 双荧光素酶报告基因分析
分别构建含有荧光素酶报告基因的载体pGL 3 的野生型FBXL3-Wt和突变型FBXL3-Mut重组质粒,并将重组质粒与hsa-miR-18b-5p Inhibitor或NC inhibitor共转染至C4-2、LNCAP细胞中,置于37 ℃培养箱继续培养48 h,收集并裂解细胞,使用双荧光素酶报告基因测定系统测量荧光素酶活性。
1.3 统计学分析
采用 Graphpad Prism 9.0统计软件进行数据分析,采用均数±标准差表示符合正态分布的计量资料,两组间比较采用独立样本t检验,采用单因素方差分析进行多组间统计分析,所有实验均独立重复3次。以Plt;0.05为差异有统计学意义。
2 结果
2.1 生物信息学分析
从GEO数据库下载全基因微阵列表达谱数据集GSE36802,热图及火山图均显示hsa-miR-18b-5p在数据集prostate cancer组表达水平明显上升(图1)。
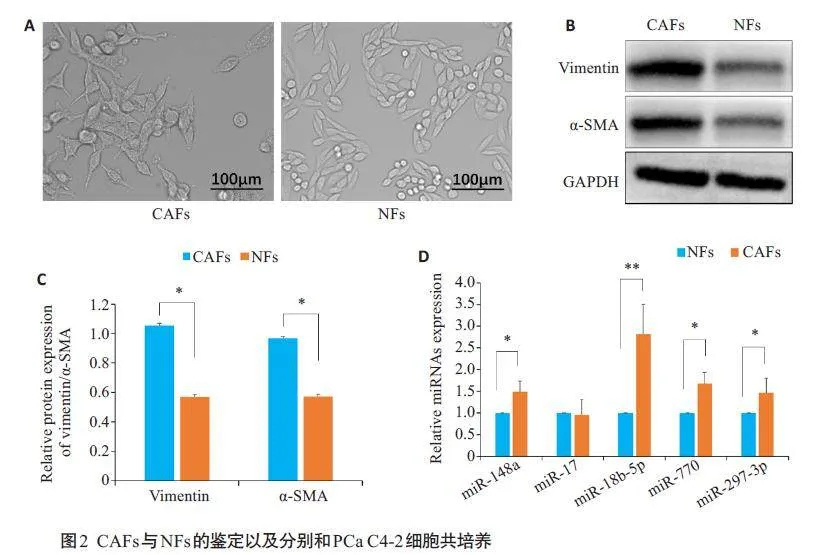
2.2 CAFs 与NFs 的鉴定以及分别和PCa C4-2 细胞共培养
光学显微镜下观察分离的CAFs 及NFs 的细胞形态均呈现典型的梭形(图2A)。Western blotting 检测CAFs及NFs特异性蛋白的表达显示,Vimentin蛋白和α-SMA蛋白在两者中均有表达,在CAFs中的表达明显强于NFs(Plt;0.05,图2B、C)。CAFs与NFs分别与PCaC4-2细胞非接触性共培养后,RT-qPCR结果显示,相对于NFs,CAFs 组hsa-miR-18b-5p 的表达水平显著升高(Plt;0.01,图2D)。
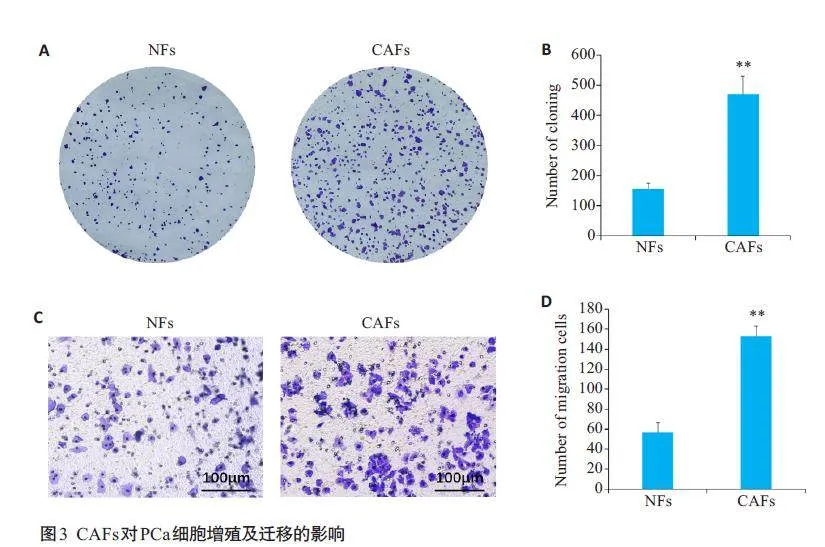
2.3 CAFs对PCa细胞增殖及迁移的影响
细胞集落形成实验结果显示,与NFs组相比,CAFs与PCa C4-2细胞共培养后可明显促进C4-2细胞的增殖(Plt;0.01,图3A、B);Transwell 迁移实验结果显示,与NFs组相比,CAFs与PCa C4-2细胞共培养后可明显促进C4-2细胞的迁移(Plt;0.01,图3C、D)。
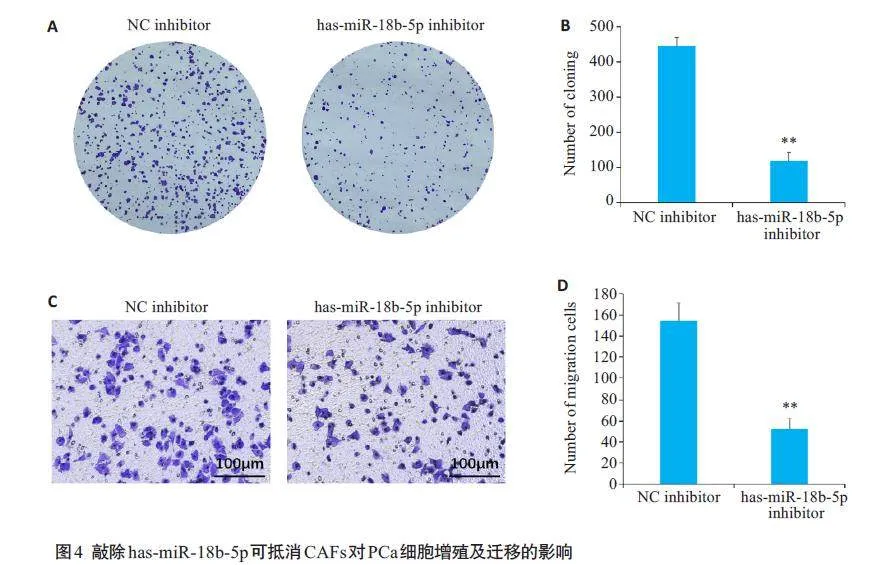
2.4 敲除has-miR-18b-5p可抵消CAFs对PCa细胞增殖及迁移的影响
细胞集落形成实验结果显示,与NC inhibitor组相比,hsa-miR-18b-5p inhibitor组的细胞增殖明显受到抑制(Plt;0.01,图4A、B);Transwell迁移实验结果显示,与NC inhibitor 组相比,hsa-miR-18b-5p inhibitor 组的细胞迁移明显受到抑制(Plt;0.01,图4C、D)。
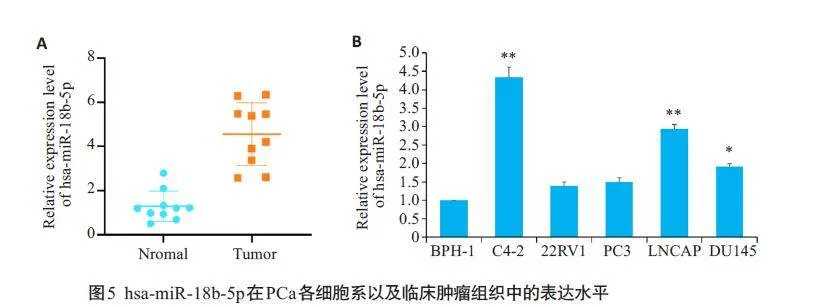
2.5 hsa-miR-18b-5p 在PCa 各细胞系以及临床肿瘤组织中的表达水平
RT-qPCR 实验结果显示, 与癌旁组织相比,hsamiR-18b-5p在PCa组织中上调(Plt;0.05,图5A);与正常前列腺增生细胞BPH-1中hsa-miR-18b-5p的表达量相比,hsa-miR-18b-5p在PCa细胞系C4-2、LNCAP、DU145细胞中表达量增高(Plt;0.05,图5B)。
2.6 hsa-miR-18b-5p对PCa细胞增殖的影响
CCK8检测结果显示,与NC inhibitor组相比,hsamiR-18b-5p inhibitor 组的细胞增殖明显受到抑制(Plt;0.05,图6A);细胞集落形成实验结果显示,与NCinhibitor 组相比,hsa-miR-18b-5p inhibitor 组的细胞集落形成的数量明显减少(Plt;0.01,图6B、C)。
2.7 hsa-miR-18b-5p对PCa细胞迁移和侵袭的影响
细胞划痕实验结果显示,与NC inhibitor 组相比,hsa-miR-18b-5p inhibitor 组的细胞迁移能力明显减弱(Plt;0.05,图7A、B);Transwell 迁移实验结果显示,与NC inhibitor 组相比,hsa-miR-18b-5p inhibitor 组的细胞发生迁移的细胞数明显降低(Plt;0.05,Plt;0.01,图7C、E);Transwell侵袭实验结果显示,与NC inhibitor组相比,hsa-miR-18b-5p inhibitor组的细胞侵袭能力明显减弱(Plt;0.05,Plt;0.01,图7D、F)。
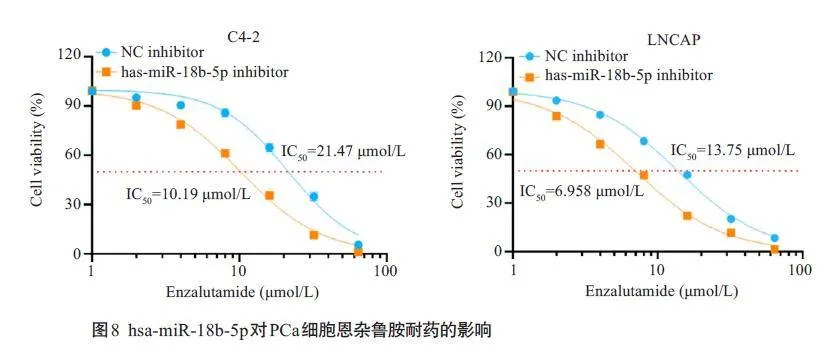
2.8 hsa-miR-18b-5p对PCa细胞恩杂鲁胺耐药的影响
IC50 测试结果显示,与NC inhibitor 组相比,hsamiR-18b-5p inhibitor组的细胞IC50值明显降低,对恩杂鲁胺的耐药能力减弱(21.47 μmol/L vs 10.19 μmol/L,13.75 μmol/L vs 6.958 μmol/L,图8)。
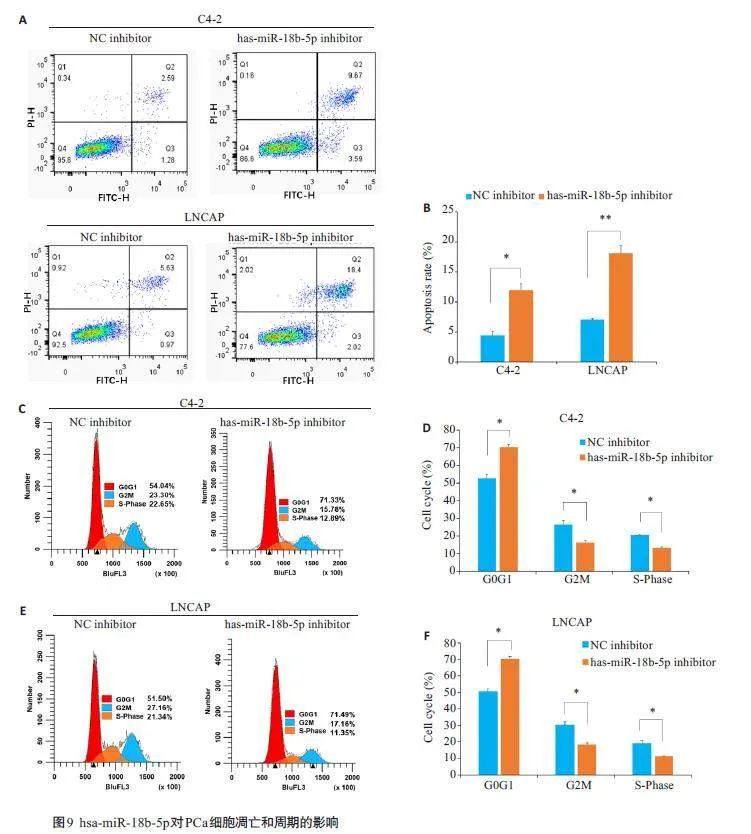
2.9 hsa-miR-18b-5p对PCa细胞的凋亡和周期的影响
流式细胞术凋亡实验结果显示,与NC inhibitor组相比,hsa-miR-18b-5p inhibitor组的细胞凋亡率明显上升(Plt;0.01,图9A、B);细胞周期实验结果显示,与NCinhibitor 组相比,hsa-miR-18b-5p inhibitor 组的G0G1期细胞比例明显增加,G2M、S-phase 期细胞比例减少(Plt;0.05,图9C~F)。
2.10 hsa-miR-18b-5p对裸鼠体内瘤负荷的影响
PCa 异种移植小鼠模型的结果显示,与LV-vector组相比,LV-anti-has-miR-18b-5p 组小鼠的肿瘤质量和体积降低(Plt;0.05,图10A~C);小鼠Kaplan-Meier生存曲线分析结果显示,与LV-vector 组相比,LV-anti-hasmiR-18b-5p组小鼠的存活时间增长(Plt;0.05,图10D)。
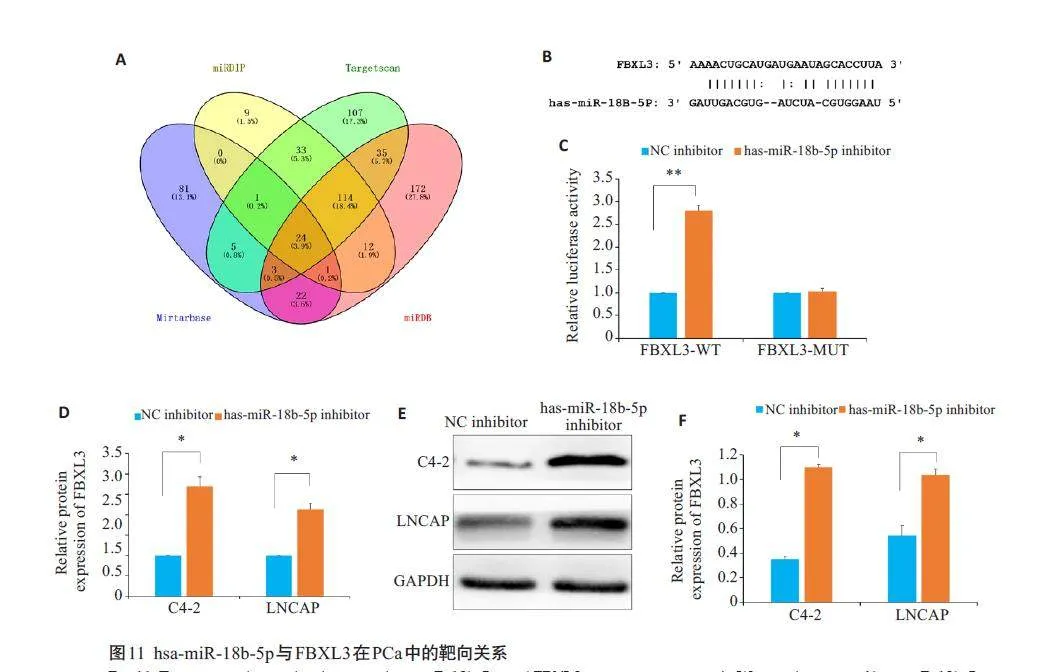
2.11 hsa-miR-18b-5p与FBXL3在PCa中的靶向关系
将miRTarBase、TargetScan 、miRDIP和miRDB等数据库筛选出hsa-miR-18b-5p 的靶基因作韦恩图(图11A);ENCORI 数据库发现hsa-miR-18b-5p 和FBXL3具有结合位点(图11B);双荧光素酶报告基因分析结果显示,hsa-miR-18b-5p inhibitor 转染增加了含有wt-FBXL3 的细胞的荧光素酶活性(Plt;0.01,图11C);RTqPCR实验结果显示, 与NC inhibitor组相比,hsa-miR-18b-5p inhibitor组细胞的FBXL3基因的表达明显升高(Plt;0.05,图11D);Western blot 实验结果显示,与NCinhibitor 组相比,hsa-miR-18b-5p inhibitor 组细胞的FBXL3蛋白的表达升高(Plt;0.05,图11E、F)。
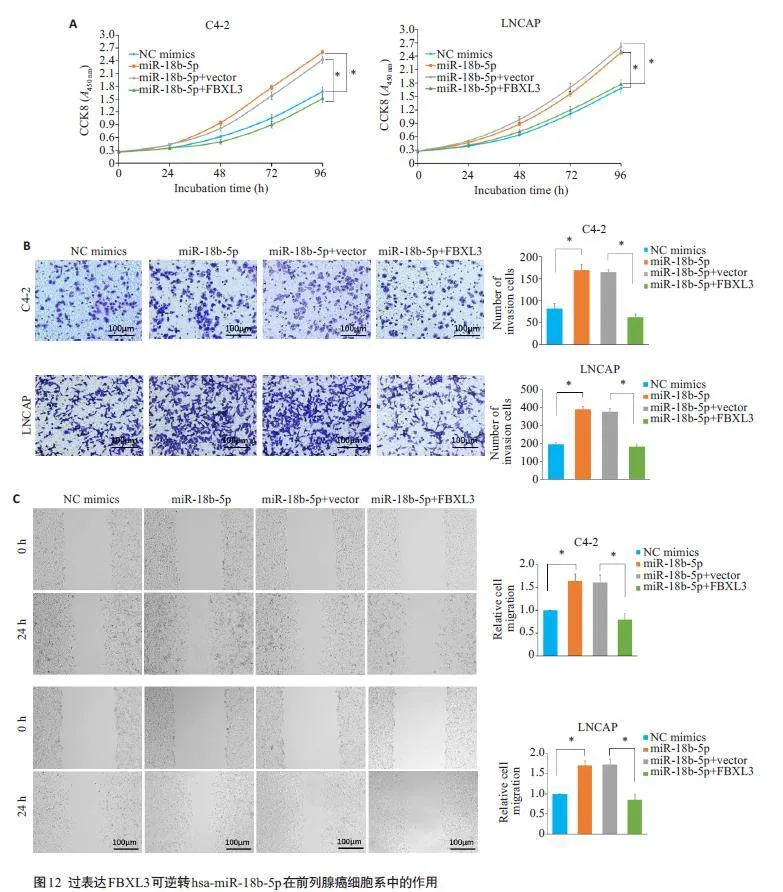
2.12 过表达FBXL3 可逆转hsa-miR-18b-5p 在前列腺癌细胞系中的作用
通过CCK8、细胞划痕实验和transwell 测定,结果显示,FBXL3的过表达能够抵消has-miR-18b-5 p模拟物在PCa细胞上的加速增殖、迁移和侵袭(图12)。
3 讨论
PCa仍然是全世界男性癌症死亡的第3大原因[32]。据研究,miRNAs 可充当基因表达的调节剂,并且具有一系列生物学功能,例如调节细胞存活、增殖、凋亡、肿瘤生长和转移[33]。miRNA与其互补的mRNA序列结合,并最终导致其翻译抑制、降解或切割。在癌症研究领域,miRNA的异常表达可能有助于人体免疫系统识别癌症和非癌症组织[34]。许多基础研究和临床研究的文章发现miRNA在癌症领域发挥着重要作用,近50%的miRNA靶基因的序列位于人类基因组与癌症相关的基因区域[35]。根据目前的研究发现,在不同癌症中发生差异表达的miRNAs不尽相同,甚至同一种miRNA在不同癌症的表达也存在差异。例如有研究指出miRNAs 的改变参与了不同类型的人类癌症的建立和发展,miRNAs 数量和质量的变化与癌症的发生,进展和转移有关[36];有研究发现miR-339 通过开启GPER1增强子来上调乳腺癌细胞中GPER1的表达,而GPER1增强子可以通过CRISPR/Cas9 系统删除增强子来阻断[37];有研究证明突变体KRAS激活PI3K-STAT3信号,从而抑制miR-34a的表达并减轻miR-34a对CD47的转录后抑制,从而逃避肺腺癌的先天免疫监视[38]。本研究通过生物信息学分析在前列腺癌中差异表达的miRNAs,发现共有17个高表达的miRNA,并通过后续在临床组织样本以及癌细胞系中验证筛选出研究基因has-miR-18b-5p。
近年来,对于miRNAs与PCa恶性进展的相关研究越来越广泛且深入。研究指出miR-138-5p 通过靶向APOBEC3B的3'-UTR来调节PCa 细胞的增殖和对化疗耐药[39];有研究阐明了miR-34a 是肿瘤抑制性miRNA,通过靶向许多对CSC存活和功能至关重要的分子发挥作用,这使其可能成为抗PCSC的治疗剂[40];研究证实缺氧是导致前列腺肿瘤中miR-21过度表达的关键因素,miR-21随后可以通过抑制肿瘤抑制基因Ras同系物家族成员B的表达来促进PCa 的进展[41]。本研究发现has-miR-18b-5p在前列腺癌中高表达,可促进前列腺癌细胞的增殖、迁移、侵袭以及对药物的耐药性,并且抑制了前列腺癌细胞的凋亡。后续对has-miR-18b-5p发生促癌作用的机制进行探究发现has-miR-18b-5p可直接靶向FBXL3,高表达的has-miR-18b-5p 显著抑制了FBXL3的表达。
大量证据支持侵袭性癌的进展受到微环境因素的强烈影响,包括缺氧、酸度、细胞外基质的组成和宿主基质细胞,统称为“反应性基质”[42]。在基质细胞中,CAFs无论是驻留的还是从循环的骨髓来源的间充质细胞中招募的,都有被报道在恶性进展中起关键作用[43]。此外,CAF与癌细胞进行双向相互作用,通过所谓的“传出方式”作用于癌细胞,从而增强其恶性程度[28]。
目前,越来越多的研究涉及miRNAs对肿瘤微环境的影响。研究发现与来自正常邻近组织的成纤维细胞相比,miR-31是在从子宫内膜癌分离的CAF中下调最多的miRNA,异位过表达miR-31 的CAFs 的基质减少了子宫内膜癌细胞的迁移和侵袭,表明miR-31可能靶向负责分泌与促进肿瘤细胞发芽有关的可溶性因子的基因[44];研究证明了乳腺基质成纤维细胞中miR-320的下调通过激活促癌分泌蛋白组来重新编程肿瘤微环境[45];有研究也证明了可以使用miR-21 的特异性反义抑制剂来防止肌成纤维细胞从祖细胞成纤维细胞转分化为TGF-β[46]。类似的,在本研究中,通过分离与纯化CAFs 和NFs 并且分别与PCa C4-2 细胞非接触性共培养。实验结果表明,相对于NFs,C4-2 细胞与CAFs 共培养后hsa-miR-18b-5p的表达水平显著升高且可促进C4-2细胞的增殖和迁移。结合本研究hsa-miR-18b-5p在PCa的发展中发挥的是促癌作用,表明本研究的实验结果与目前主流研究关于肿瘤微环境中CAFs 起促癌作用的结论相一致。本研究是第一个全面评估hasmiR-18b-5p 与CAFs 之间的相互影响以及其在PCa 中作用及机制的研究。
已有研究报道has-miR-18b-5p 通过靶向TCEAL7、TERT、VMA21、NLRP7、KLK3、OLFM3、POSTN、MAGED4B、KIR3DL3 和CEACAM5 发挥癌基因或肿瘤抑制基因的作用。本研究中,FBXL3 被确定为has-miR-18b-5p的直接靶点。FBXL3基因位于细胞核中,编码的蛋白质属于Fbls类,除了F-box外,还包含几个串联的富含亮氨酸的重复序列。在已有的研究中,FBXL3发挥肿瘤抑制基因作用。例如,有研究通过Kaplan-Meier 生存曲线分析FBXL3 与NSCLC患者总生存率以及在功能上进行了MTT、集落形成测定和transwell测定确定FBXL3抑制NSCLC细胞增殖、迁移和侵袭,表明FBXL3在NSCLC中具有潜在的肿瘤抑制作用[47];有文献阐述了FBXL3 和CRY2 协同降级细胞增殖的关键调节剂c-MYC,从而抑制肿瘤细胞的增殖,发挥肿瘤抑制基因的作用[48];我们首次证实了受hasmiR-18b-5p调控的FBXL3能够抑制PCa细胞的增殖、迁移和侵袭,并在PCa中发挥抑癌作用。
综上所述,本研究首次提示CAFs可上调PCa细胞has-miR-18b-5p 的表达,敲低has-miR-18b-5p 可抵消CAFs 对PCa 细胞增殖及迁移的影响;而has-miR-18b-5p可抑制FBXL3的表达,从而促进PCa细胞增殖和转移,同时抑制PCa细胞的凋亡,提示受CAFs上调的hasmiR-18b-5p可通过靶向FBXL3调控PCa的恶性进展,这可为PCa治疗策略提供新的见解。
参考文献:
[1] Sehn JK. Prostate cancer pathology: recent updates and controversies[J]. Mo Med, 2018, 115(2): 151-5.
[2] Kimura T, Egawa S. Epidemiology of prostate cancer in Asiancountries[J]. Int J Urol, 2018, 25(6): 524-31.
[3] Zelic R, Garmo H, Zugna D, et al. Predicting prostate cancer deathwith different pretreatment risk stratification tools: a head-to-headcomparison in a nationwide cohort study[J]. Eur Urol, 2020, 77(2):180-8.
[4] Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020:GLOBOCAN estimates of incidence and mortality worldwide for 36cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209-49.
[5] Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020[J]. CACancer J Clin, 2020, 70(1): 7-30.
[6] Taylor LG, Canfield SE, Du XL. Review of major adverse effects ofandrogen-deprivation therapy in men with prostate cancer[J].Cancer, 2009, 115(11): 2388-99.
[7] Zarour L, Alumkal J. Emerging therapies in castrate-resistantprostate cancer[J]. Curr Urol Rep, 2010, 11(3): 152-8.
[8] Boulos S, Mazhar D. The evolving role of chemotherapy in prostatecancer[J]. Future Oncol, 2017, 13(12): 1091-5.
[9] Sebesta EM, Anderson CB. The surgical management of prostatecancer[J]. Semin Oncol, 2017, 44(5): 347-57.
[10]Shevach J, Chaudhuri P, Morgans AK. Adjuvant therapy in high-riskprostate cancer[J]. Clin Adv Hematol Oncol, 2019, 17(1): 45-53.
[11] Chistiakov DA, Myasoedova VA, Grechko AV, et al. Newbiomarkers for diagnosis and prognosis of localized prostate cancer[J]. Semin Cancer Biol, 2018, 52(Pt 1): 9-16.
[12]Michlewski G, Cáceres JF. Post-transcriptional control of miRNAbiogenesis[J]. RNA, 2019, 25(1): 1-16.
[13]Liu T, Zhang Q, Zhang JK, et al. EVmiRNA: a database of miRNAprofiling in extracellular vesicles[J]. Nucleic Acids Res, 2019, 47(D1): D89-93.
[14]Ali Syeda Z, Langden SSS, Munkhzul C, et al. Regulatorymechanism of microRNA expression in cancer[J]. Int J Mol Sci,2020, 21(5): 1723-9.
[15]Friedman RC, Farh KKH, Burge CB, et al. Most mammalianmRNAs are conserved targets of microRNAs[J]. Genome Res,2009, 19(1): 92-105.
[16]Cocks A, Del Vecchio F, Martinez-Rodriguez V, et al. Pro-tumoralfunctions of tumor-associated macrophage EV-miRNA[J]. SeminCancer Biol, 2022, 86(Pt 1): 58-63.
[17]Del Valle-Morales D, Le P, Saviana M, et al. The epitranscriptome inmiRNAs: crosstalk, detection, and function in cancer[J]. Genes,2022, 13(7): 1289-95.
[18]Smolarz B, Durczyński A, Romanowicz H, et al. miRNAs in cancer(review of literature)[J]. Int J Mol Sci, 2022, 23(5): 2805-13.
[19] Iqbal MA, Arora S, Prakasam G, et al. MicroRNA in lung cancer:role, mechanisms, pathways and therapeutic relevance[J]. MolAspects Med, 2019, 70: 3-20.
[20]Kt RD, Karthick D, Saravanaraj KS, et al. The roles of microRNA inpancreatic cancer progression[J]. Cancer Invest, 2022, 40(8): 700-9.
[21] Ismail A, Abulsoud AI, Fathi D, et al. The role of miRNAs in ovariancancer pathogenesis and therapeutic resistance-A focus on signalingpathways interplay[J]. Pathol Res Pract, 2022, 240: 154222.
[22]Hwang GR, Yuen JG, Ju JF. Roles of microRNAs in gastrointestinalcancer stem cell resistance and therapeutic development[J]. Int JMol Sci, 2021, 22(4): 1624-36.
[23]Doghish AS, Ismail A, El-Mahdy HA, et al. A review of thebiological role of miRNAs in prostate cancer suppression andprogression[J]. Int J Biol Macromol, 2022, 197: 141-56.
[24]Mao XQ, Xu J, Wang W, et al. Crosstalk between cancer-associatedfibroblasts and immune cells in the tumor microenvironment: newfindings and future perspectives[J]. Mol Cancer, 2021, 20(1): 131-8.
[25]Tsoumakidou M. The advent of immune stimulating CAFs in cancer[J]. Nat Rev Cancer, 2023, 23(4): 258-69.
[26]Wang YF, Liang HY, Zheng J. Exosomal microRNAs mediatingcrosstalk between cancer cells and cancer-associated fibroblasts inthe tumor microenvironment[J]. Pathol Res Pract, 2022, 239:154159.
[27]Bullock MD, Pickard KM, Nielsen BS, et al. Pleiotropic actions ofmiR-21 highlight the critical role of deregulated stromal microRNAsduring colorectal cancer progression[J]. Cell Death Dis, 2013, 4(6):e684-92.
[28]Gandellini P, Giannoni E, Casamichele A, et al. MiR-205 hinders themalignant interplay between prostate cancer cells and associatedfibroblasts[J]. Antioxid Redox Signal, 2014, 20(7): 1045-59.
[29]Yan ZQ, Sheng ZM, Zheng YH, et al. Cancer-associated fibroblastderivedexosomal miR-18b promotes breast cancer invasion andmetastasis by regulating TCEAL7[J]. Cell Death Dis, 2021, 12(12):1120-32.
[30]Xue F, Xu YH, Shen CC, et al. Non-coding RNA LOXL1-AS1 exhibits oncogenic activity in ovarian cancer via regulation of miR-18b-5p/VMA21 axis[J]. Biomedecine Pharmacother, 2020, 125:109568.
[31]Wang YY, Yan L, Yang S, et al. Long noncoding RNA AC073284.4suppresses epithelial-mesenchymal transition by sponging miR-18b-5p in paclitaxel-resistant breast cancer cells[J]. J Cell Physiol, 2019,234(12): 23202-15.
[32]Komura K, Sweeney CJ, Inamoto T, et al. Current treatmentstrategies for advanced prostate cancer[J]. Int J Urol, 2018, 25(3):220-31.
[33]Swarbrick S, Wragg N, Ghosh S, et al. Systematic review of miRNAas biomarkers in Alzheimer's disease[J]. Mol Neurobiol, 2019, 56(9): 6156-67.
[34]Pottoo FH, Iqubal A, Iqubal MK, et al. miRNAs in the regulation ofcancer immune response: effect of miRNAs on cancerimmunotherapy[J]. Cancers, 2021, 13(23): 6145-56.
[35]Khan AQ, Ahmed EI, Elareer NR, et al. Role of miRNA-regulatedcancer stem cells in the pathogenesis of human malignancies[J].Cells, 2019, 8(8): 840-8.
[36]Hussen BM, Hidayat HJ, Salihi A, et al. MicroRNA: a signature forcancer progression[J]. Biomedecine Pharmacother, 2021, 138:111528.
[37]Liang Y, Lu Q, Li W, et al. Reactivation of tumour suppressor inbreast cancer by enhancer switching through NamiRNA network[J]. Nucleic Acids Res, 2021, 49(15): 8556-72.
[38]Hu HH, Cheng RJ, Wang YB, et al. Oncogenic KRAS signalingdrives evasion of innate immune surveillance in lungadenocarcinoma by activating CD47[J]. J Clin Invest, 2023, 133(2):e153470.
[39]Liu LN, Zhang Y, Hu X, et al. MiR-138-5p inhibits prostate cancercell proliferation and chemoresistance by targeting APOBEC3B[J].Transl Oncol, 2023, 35: 101723-36.
[40]Li WJ, Liu XZ, Dougherty EM, et al. MicroRNA-34a, prostatecancer stem cells, and therapeutic development[J]. Cancers, 2022,14(18): 4538-50.
[41]Angel CZ, Stafford MYC, McNally CJ, et al. MiR-21 is induced byhypoxia and down-regulates RHOB in prostate cancer[J]. Cancers,2023, 15(4): 1291-309.
[42]Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation[J]. Cell, 2011, 144(5): 646-74.
[43]Cirri P, Chiarugi P. Cancer-associated-fibroblasts and tumour cells: adiabolic liaison driving cancer progression[J]. Cancer MetastasisRev, 2012, 31(1/2): 195-208.
[44]Aprelikova O, Yu X, Palla J, et al. The role of miR-31 and its targetgene SATB2 in cancer-associated fibroblasts[J]. Cell Cycle, 2010, 9(21): 4387-98.
[45]Bronisz A, Godlewski J, Wallace JA, et al. Reprogramming of thetumour microenvironment by stromal PTEN-regulated miR-320[J].Nat Cell Biol, 2011, 14(2): 159-67.
[46]Yao Q, Cao SY, Li C, et al. Micro-RNA-21 regulates TGF-β-inducedmyofibroblast differentiation by targeting PDCD4 in tumor-stromainteraction[J]. Int J Cancer, 2011, 128(8): 1783-92.
[47]Wang DZ, Han X, Li C, et al. FBXL3 is regulated by miRNA-4735-3p and suppresses cell proliferation and migration in non-small celllung cancer[J]. Pathol Res Pract, 2019, 215(2): 358-65.
[48]Huber AL, Papp SJ, Chan AB, et al. CRY2 and FBXL3cooperatively degrade c-MYC[J]. Mol Cell, 2016, 64(4): 774-89.
(编辑:林 萍)
基金项目:安徽省自然科学基金(2008085QH358);安徽省高等学校自然科学研究重点项目(2023AH051942);蚌埠医学院2023 年度研究生科研创新计划项目(Byycx23114);慢性疾病免疫学基础与临床安徽省重点实验室开放课题基金(AHIAI2022K01)
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13 08:59:50
云南医药(2021年3期)2021-07-21 05:40:30
宁夏医学杂志(2020年3期)2021-01-21 08:23:24
天然产物研究与开发(2018年4期)2018-05-07 06:48:03
中国报道(2018年2期)2018-04-20 04:12:46
金融博览(2016年7期)2016-08-16 18:44:41
绿色中国(2016年1期)2016-06-05 09:02:59
中国现代医学杂志(2015年26期)2015-12-23 11:04:22
中国当代医药(2015年33期)2015-03-01 02:09:20
中国医疗美容(2015年5期)2015-02-03 03:02:09
