
盐碱胁迫下燕麦幼苗生长生理差异及耐盐碱阈值
2024-11-02 00:00:00张碧茹米俊珍赵宝平刘景辉
西北农业学报 2024年10期
摘 要 为研究混合盐碱胁迫对燕麦幼苗生长、生理的影响,计算耐盐碱阈值,选用相对盐碱敏感品种‘白燕5号’和相对耐盐碱品种‘坝莜18号’为试验材料,设置不同浓度的盐碱胁迫处理,分析燕麦生长、生理指标的变化。结果表明:(1)不同浓度盐碱胁迫对‘白燕5号’和‘坝莜18号’幼苗的株高、根长和生物量有不同程度的抑制作用,分别在150 mmol/L 和200 mmol/L胁迫时差异显著。(2)随着胁迫浓度的升高,‘白燕5号’SPAD值、Pn、Gs及Tr均呈下降趋势,在100 mmol/L胁迫时差异显著;‘坝莜18号’SPAD值、Pn、Gs、Tr则表现出先升后降的趋势,Ci呈先降后升的趋势,胁迫达到200 mmol/L时差异显著。(3)‘白燕5号’和‘坝莜18号’脯氨酸(Pro)和丙二醛(MDA)含量随着胁迫浓度的增加均呈上升趋势,过氧化氢酶(CAT)活性则呈相反趋势,超氧化物歧化酶(SOD)、过氧化物酶(POD)活性呈先升后降的趋势,‘白燕5号’和‘坝莜18号’SOD活性分别在150 mmol/L和200 mmol/L胁迫时达到峰值且差异显著。(4)以株高较对照降低50%为标准,得‘白燕5号’和‘坝莜18号’幼苗对盐碱胁迫的耐受阈值分别为163.4 mmol/L和200.0 mmol/L。综上所述,燕麦幼苗混合盐碱耐受阈值的确定,可为盐碱地燕麦栽培与推广应用提供理论依据。
关键词 燕麦;盐碱胁迫;生理差异;耐盐碱阈值
土壤盐碱化是影响植物生长发育和导致作物减产的主要非生物胁迫因子之一。中国约有 9 913 万 hm2 盐碱地,约占国土面积的1/4[1]。盐渍化土壤对植物的生长发育具有严重的负面影响。盐碱胁迫不仅会减缓植物吸收水分的速率, 抑制植物叶片和根系的生长速度, 盐分还可能参与蒸腾作用,对叶片细胞产生伤害[2],而且会破坏植物细胞或组织内外的渗透平衡、离子稳态和生长激素等的调节,从而抑制植物正常生长发育[3]。燕麦( Avena sativa L.)是中国北方地区优势特色粮饲兼用作物,具有抗寒、耐贫瘠和耐盐碱等优良特性,被称为是改良盐碱地的先锋作物[4],内蒙古种植燕麦面积居全国首位。王苗苗等[5]发现盐胁迫刺激燕麦合成更多的脯氨酸以维持渗透势平衡,从而抵抗盐胁迫造成的伤害。刘建新等[6-7]在对燕麦幼苗的盐碱胁迫试验中发现盐碱胁迫伤害了幼苗的光合系统,高浓度盐碱胁迫导致 Pn、Gs、Tr 和 Ls 显著降低及 Ci 明显提高,有机酸和脯氨酸可能是裸燕麦适应盐碱的主要渗透调节物质。目前,对盐、碱胁迫下燕麦的生长发育及生理响应机制研究较多,混合盐碱胁迫对燕麦生理响应的研究较少且多集中于一个品种的研究,但关于不同耐性品种的耐受阈值研究鲜有报道,因此,本研究以盐碱敏感品种‘白燕5号’ (Baiyan 5)和耐盐碱品种‘坝莜18号’(Bayou 18)为试验材料,研究不同混合盐碱胁迫对不同耐性燕麦幼苗生长和生理的响应,通过分析在不同浓度混合盐碱条件下燕麦幼苗生长、SPAD值、净光合速率等指标的变化及影响,明确不同耐性燕麦对混合盐碱的耐受阈值,为筛选燕麦耐盐碱品种及盐碱地合理利用提供理论依据。
1 材料与方法
1.1 试验材料
供试燕麦品种为前期萌发期试验筛选得到的相对盐碱敏感型‘白燕5号’(B5)和相对耐盐碱型‘坝莜18号’(B18),由吉林省白城市农业科学院和河北省张家口农科院提供。
1.2 试验方法
试验于2021-2022年在内蒙古农业大学燕麦产业研究中心温室进行,使用装有蛭石与泥炭土(按质量比1∶1的比例混合而成)的塑料盆(直径20 cm,高25 cm),每盆播种30粒,每盆定苗20株,在温室进行培养,温度20±5 ℃,定期浇营养液。幼苗三叶一心时(20 d),将每个品种燕麦幼苗分成6组,每组3盆,于18:00时开始如下处理:对照(CK):正常霍格兰营养液;混合盐碱(NaCl∶Na2SO4∶NaHCO3∶Na2CO3摩尔比为1∶9∶9∶1)溶液Y1:50 mmol/L、Y2:100 mmol/L、Y3:150 mmol/L、Y4:200 mmol/L、Y5:250 mmol/L;浇灌盐碱溶液时要注意逐级增加溶液浓度,可按每隔24 h增加50~150 mmol/L[JP]处理,防止发生盐激伤害,导致幼苗死亡。每个处理设置3次重复,处理第7 d时分别取各处理各品种幼苗叶片,进行生长及生理指标的测定。
1.3 测定指标及方法
1.3.1 形态指标与叶片相对含水量 株高:指植物根颈部到顶部之间的距离,其中顶部指主茎 顶部。
根长:指植物根茎部到顶部之间的距离,其中顶部指主根顶部。
鲜质量(FW)/干质量(DW):将地上部和地下部分开,并分别称量鲜质量(FW),然后将鲜样在105 ℃的烘箱中杀青20 min,再转到80 ℃烘干至恒量,并分别称量干质量(DW)[8]。
叶片相对含水量(RWC):称取植株新鲜叶片(0.5 g)以获得鲜质量(Wf),然后将叶片放在水面上4 h以获得饱和质量(WS),最后将叶片烘干(85 ℃)至恒量(24 h)以获得干质量(Wd)[9]。
叶片相对含水量=(Wf -Wd)/(WS-Wd)。
1.3.2 光合特性指标 SPAD值:用手持叶绿素仪(美国进口SPAD-502Plus:购自青岛聚创嘉恒分析仪器有限公司)测定叶片SPAD值。
气体交换参数:在晴天上午9:30-11:00时间段,使用CIRAS-3便携式光合作用测定系统(美国PP Systems公司),在大气CO2浓度为400 μmol/mol,温度为26 ℃,光强约为400 μmol/(m2·s)的温室中测定不同燕麦材料叶片的气孔导度(Gs)、净光合速率(Pn)、胞间CO2浓度(Ci)以及蒸腾速率(Tr)。分别选取对照和混合盐碱溶液处理后的材料从顶部开始第2个叶片进行测定,每个处理进行3次重复。
1.3.3 生理指标 脯氨酸(Pro) 含量:采用茚三酮法;丙二醛(MDA)含量:采用硫代巴比妥酸法;超氧化物歧化酶(SOD)活性:用氮蓝四唑(NBT)光还原法;过氧化物酶(POD)活性:用愈创木酚法;过氧化氢酶(CAT)活性:采用紫外吸收法,以上指标均参照李合生[10]的方法进行测定。
1.3.4 耐盐碱阈值计算 一般常采用植物生长量下降50%时的盐浓度作为其耐盐阈值[11-12]。本研究以株高为因变量、混合盐碱浓度为自变量构建拟合曲线,并建立回归方程,以株高下降50%的土壤含盐量作为其耐盐碱阈值。
1.4 数据处理
用Excel 2016整理数据和制图,用SAS 9.0和SPSS 22.0软件进行差异显著性和相关性分析。[JP]
2 结果与分析
2.1 混合盐碱胁迫下燕麦的幼苗生长及叶片相对含水量的变化
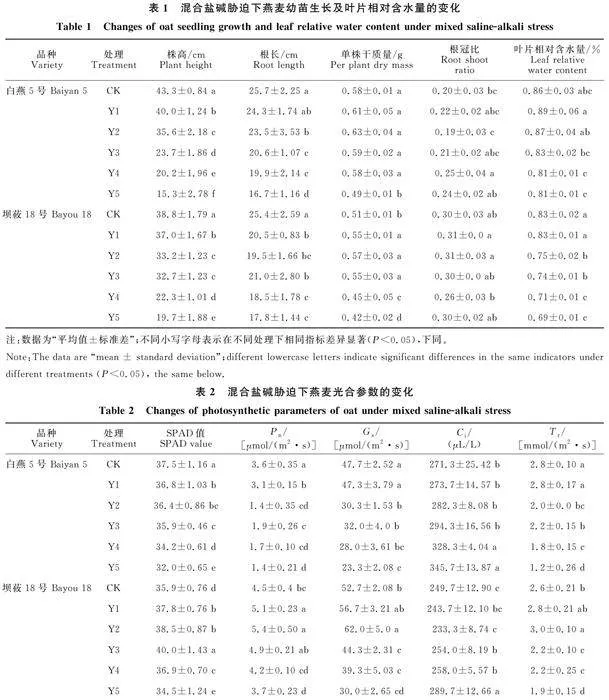
从表1中可以看出,随着混合盐碱浓度增大,‘白燕5号’和‘坝莜18号’幼苗株高和根长均呈下降趋势,较CK分别下降了7.6%~64.7%、 4.6%~23.5%和5.4%~35.0%、21.3%~ 49.2%。单株干质量和叶片相对含水量呈现先上升后下降的趋势,‘白燕5号’在胁迫浓度50、100、150 mmol/L时较CK提高均不显著(P< 0.05),而‘坝莜18号’单株干质量在胁迫浓度100 mmol/L时较CK提高了11.8%,差异显著;‘白燕5号’单株干质量和叶片相对含水量分别在胁迫浓度250 mmol/L 和200 mmol/L时较CK分别下降15.5%和5.8%,且达到显著差异水平,‘坝莜18号’单株干质量和叶片相对含水量分别在≥200 mmol/L和≥100 mmol/L下降达到显著差异水平,较CK分别下降了11.8%~17.6%和9.6%~16.9%。
2.2 混合盐碱胁迫下燕麦光合参数的变化
随着胁迫浓度的增加,‘白燕5号’SPAD值、Pn、Gs及Tr均呈下降趋势(表2),与CK相比,在100 mmol/L时光合参数变化达到显著差异水平(P<0.05),Ci呈上升趋势,在200 mmol/L时差异显著;而‘坝莜18号’SPAD值、Pn、Gs及Tr均则呈先上升后下降的趋势,Ci呈先下降后上升的趋势,胁迫达到250 mmol/L时SPAD值、Pn、Gs及Tr较CK均显著下降,Ci也上升至最大值且差异显著。
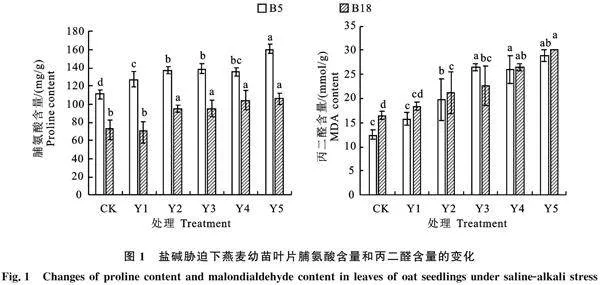
2.3 混合盐碱胁迫下燕麦叶片脯氨酸和丙二醛含量的变化
由图1可知,在不同浓度胁迫处理下‘白燕5号’叶片脯氨酸含量均高于‘坝莜18号’,且‘白燕5号’随着胁迫浓度增加脯氨酸含量也有不同程度的增加,较CK均达到显著差异水平,在250 mmol/L胁迫时增幅最大(43.7%);而‘坝莜18号’在50 mmol/L处理时脯氨酸含量较CK变化不显著,在100 mmol/L以上时脯氨酸含量显著增加。
随着胁迫浓度的增加,‘白燕5号’叶片丙二醛含量呈先上升后下降再上升的趋势,在100~150 mmol/L时升高达到显著差异,在200 mmol/L时开始降低,但差异不明显,在250 mmol/L时丙二醛含量最大,而‘坝莜18号’叶片丙二醛含量则呈上升趋势,在100~250 mmol/L时较CK明显升高,达到显著差异水平。
2.4 混合盐碱胁迫下燕麦叶片SOD、POD和CAT活性的变化
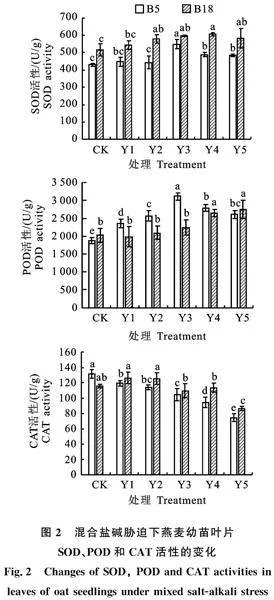
由图2可知,随着混合盐碱胁迫浓度的升高,‘白燕5号’叶片SOD和POD活性均呈先上升后下降的趋势,在150 mmol/L胁迫时活性均达到最高,较CK分别提高了27.4%和65.8%,且差异显著;而‘坝莜18号’叶片SOD活性均呈先上升后下降趋势,在200 mmol/L胁迫时SOD活性达到最高,较CK提高了17.7%,POD活性则随着胁迫浓度的升高而增加,在200 mmol/L胁迫时POD活性显著增加,与CK相比增幅达 31.0%。混合盐碱胁迫浓度提高,‘白燕5号’叶片CAT活性明显下降,而‘坝莜18号’叶片CAT活性与CK相比,在250 mmol/L时显著下降 25.3%。
2.5 燕麦耐盐碱指标选择
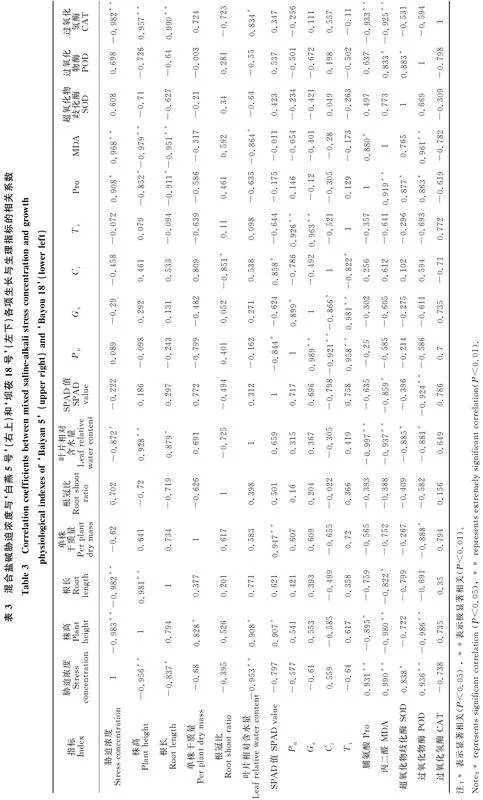
通过对混合盐碱胁迫浓度与两燕麦品种各项生长与生理指标的相关性进行分析,从而鉴定燕麦的耐盐碱性能(表3)。结果显示,‘白燕5号’和‘坝莜18号’的各项生长指标中株高相关系数绝对值最大,各项生理指标中丙二醛相关系数绝对值最大,分别与胁迫浓度呈极显著负相关和极显著正相关(P<0.01)。因此,株高可作为耐盐碱鉴定的生长指标,丙二醛可作为耐盐碱鉴定的生理指标。
2.6 确定燕麦的耐盐碱阈值
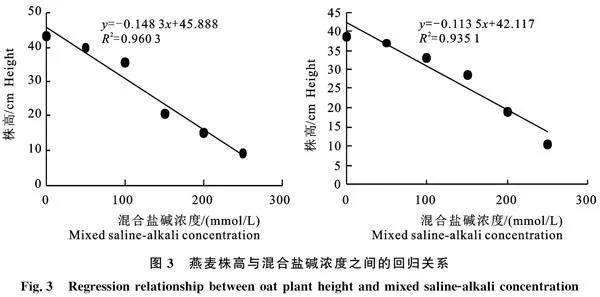
以株高为因变量(y),混合盐碱胁迫浓度为自变量(x)建立回归方程,结果见图3,以株高生长量下降50%时所对应的混合盐碱浓度作为‘白燕5号’和‘坝莜18号’幼苗对盐碱胁迫的耐受阈值,其值分别为163.4 mmol/L和200.0 mmol/L。
3 讨 论
作物的耐盐碱特性是受多基因控制的数量性状[13],且在不同生长时期的表现也存在差异[14],苗期是作物对盐碱胁迫最敏感的阶段之一[15]。已有研究发现,作物在盐碱胁迫下会通过自身生长的改变和生理生化响应的调节来适应,以及调整生物量的分配来构建和维持逆境下的存活与生长[16-17]。生物量是植物对盐碱胁迫响应的综合反映。本研究结果表明,随着混合盐碱胁迫浓度增大,幼苗叶片相对含水量和生物量明显下降,叶片相对含水量和生长量呈显著负相关(P<0.05),说明盐碱胁迫下水分亏缺是燕麦幼苗生长受抑的主要因素之一,这与王波等[18]和范远等[19]的结果一致。
叶片是植物光合作用主要器官,光合作用是植物最重要的代谢过程,因此测定植株光合指标是认识植物耐盐碱性的一个可靠途径。叶绿素是绿色植物进行光合作用的主要色素。在胁迫作用后,植物对胁迫的响应表现为光合作用下降和光合色素含量降低[5]。在本研究中,随着盐碱浓度的递增,相对盐敏感品种‘白燕5号’SPAD值呈下降趋势,相对耐盐碱品种‘坝莜18号’SPAD值呈先升后降的趋势。王晓春等[20]研究结论“低浓度的盐碱胁迫促进了苜蓿叶绿素的积累,但中高浓度盐碱胁迫致使叶绿素含量下降”与本研究中耐盐碱‘坝莜18号’变化一致,说明耐盐碱品种在受到盐碱胁迫后会通过自身防御提高光合作用,进而增强抗逆性。Farquhar等[21]认为净光合速率下降与气孔导度等有关,同时王旭明等[22]认为低盐胁迫下导致植物叶片光合速率下降、胞间CO2浓度上升的主要因素为气孔因素,但在高盐浓度下,植物叶片的非气孔因素会转为主导。本研究发现,随着胁迫浓度升高,‘白燕5号’Pn、Gs、Tr持续下降,Ci则持续上升,‘坝莜18号’Pn、Gs、Tr在胁迫浓度为100~150 mmol/L时达到最高值,250 mmol/L时最低,而Ci则变化趋势相反,说明非气孔因素是高浓度盐碱胁迫光合作用下降的主要原因,与刘建新等[7]研究结果一致。
盐碱胁迫对植物造成的伤害首先是渗透胁迫,进而导致活性氧代谢紊乱造成细胞膜脂过氧化,甚至诱导细胞死亡[23]。本研究表明,随着混合盐碱处理浓度增大,游离脯氨酸含量及膜脂氧化产物MDA含量均呈上升趋势,与张婷等[24]以枸杞为材料研究结果一致。可能是由于胁迫启动了体内的渗透调节机制,进而激发了其防御系统对外界的不良环境进行抵抗[25]。何淼等[26]研究表明,在受到复合盐碱胁迫处理时,芒幼苗丙二醛含量不断升高,也与本研究结果一致,说明燕麦体内的脯氨酸含量与丙二醛含量可以作为潜在的盐碱胁迫鉴定指标。因两燕麦品种耐性不同,所以渗透调节物质变化也不一致,同一浓度胁迫下‘白燕5号’脯氨酸含量明显高于‘坝莜18号’,而丙二醛含量表现为‘坝莜18号’高于‘白燕5号’,可能是因为丙二醛含量对盐碱胁迫的表现更敏感。
研究发现,逆境胁迫下植物叶片中 SOD、POD和CAT活性增加,或者严重胁迫时呈先增加后降低的规律[27-28]。在本试验中,随着盐碱胁迫浓度的增加,两燕麦品种CAT活性均呈下降趋势,而‘白燕5号’表现SOD和POD活性为先升后降的变化趋势,‘坝莜18号’则表现为上升趋势,表明‘坝莜18号’抗氧化酶活性更强,能更好地保持活性氧代谢平衡,以减缓活性氧对膜的伤害。张喜斌等[29]开展 6 个苜蓿品种的苗期耐盐性鉴定,在300 mmol/L NaCl胁迫下SOD、POD活性均出现了下降,但CAT活性却有不同程度的上升。谢楠等[30]以不同浓度盐胁迫对不同小黑麦品种进行胁迫,发现POD活性增高,而CAT在有的品种上表现为在低浓度盐胁迫下升高、在中高浓度盐胁迫下降低,有的品种却一直上升。这都与本研究结果有所不同,由此可见,抗氧化酶活性的变化较敏感,在植物种类、品种间差异 较大。
4 结 论
在高浓度混合盐碱胁迫下,与盐碱敏感型燕麦相比,耐盐碱型燕麦保持着较高的光合能力和抗氧化能力,主要通过合成叶绿素来维持其叶片的光合能力,同时通过调节高渗透调节物质含量和SOD、POD活性,降低氧化伤害,提高其抗 逆性。
研究以株高较对照下降50%时所对应的盐碱浓度作为衡量燕麦对盐碱胁迫的耐受阈值,最终确定‘白燕5号’幼苗对盐碱胁迫的耐受阈值为163.4 mmol/L,‘坝莜18号’幼苗对盐碱胁迫的耐受阈值为200.0 mmol/L。
参考文献 Reference:
[1] 刘小京,郭 凯,封晓辉,等.农业高效利用盐碱地资源探讨[J].中国生态农业学报,2023,31(3):345-353.
LIU X J,GUO K,FENG X H,et al.Discussion on efficient utilization of saline-alkali land resources in agriculture [J].Chinese Journal of Eco-Agriculture, 2023,31(3):345-353.
[2]赵天然,车路平,谢会成,等.流苏树对NaCl的生理响应及耐盐阈值分析[J].植物生理学报,2022,58(10):1935-1945.
ZHAO T R,CHE L P,XIE H CH,et al.Physiological response of tassel trees to nacl and analysis of salt tolerance threshold [J].Acta Phytophysiologica Sinica,2022,58(10):1935-1945.
[3]雷雪峰,赵宝平,刘景辉,等.燕麦对盐碱胁迫的响应机制及缓解措施研究进展[J].种子,2019,38(4):62-66.
LEI X F,ZHAO B P,LIU J H,et al.Research progress on response mechanism and mitigation measures of oat under saline-alkali stress [J].Seed,2019,38(4):62-66.
[4]张 波,任长忠.燕麦基因组学与分子育种研究进展[J].植物学报,2022,57(6):785-791.
ZHANG B,REN CH ZH.Advances in genomics and molecular breeding of oat [J].Bulletin of Botany,202,57(6):785-791.
[5]王苗苗,周向睿,梁国玲,等.5份燕麦材料苗期耐盐性综合评价[J].草业学报,2020,29(8):143-154.
WANG M M,ZHOU X R,LIANG G L,et al.Comprehensive evaluation of salt tolerance of five oat materials at seedling stage [J].Acta Prataculturae Sinica,2020,29(8):143-154.
[6]刘建新,王金成,王瑞娟,等.混合盐碱胁迫下裸燕麦的种子萌发和幼苗逆境生理特征[J].植物研究,2016,36(2):224-231.
LIU J X,WANG J CH,WANG R J,et al.Seed germination and seedling stress physiological characteristics of naked oat under mixed salt-alkali stress [J].Plant Research,2016,36(2):224-231.
[7]刘建新,王金成,王瑞娟,等.混合盐碱胁迫对燕麦幼苗矿质离子吸收和光合特性的影响[J].干旱地区农业研究,2017,35(1):178-184,239.
LIU J X,WANG J CH,WANG R J,et al.Effects of mixed salt-alkali stress on mineral ion absorption and photosynthetic characteristics of oat seedlings [J].Agricultural Research in the Arid Areas,2017,35(1):178-184,239.
[8]KHOSHBAKHT D,ASGHARI M R,HAGHIGHI M.Influence of foliar application of polyamines on growth,gas-exchange characteristics,and chlorophyll fluorescence in bakraii citrus under saline conditions [J].Photosynthetica,2018,56(2):731-742.
[9]FAROOQ M,WAHID A,LEE D J.Exogenously applied polyamines increase drought tolerance of rice by improving leaf water status,photosynthesis and membrane properties [J].Acta Physiologiae Plantarum,2009,31(5):937-945.
[10] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2002:134,164-167,199,258-260.
LI H SH.Experimental Principles and Techniques of Plant Physiology and Biochemistry [M].Beijing:Higher Education Press,2002:134,164-167,199,258-260.
[11]刘一明,程凤枝,王 齐,等.四种暖季型草坪植物的盐胁迫反应及其耐盐阈值[J].草业学报,2009,18(3):192-199.
LIU Y M,CHENG F ZH,WANG Q,et al.Salt stress response and salt tolerance threshold of four warm-season turfgrass plants [J].Acta Prataculturae Sinica,2009,18(3):192-199.
[12]范希峰,侯新村,朱 毅,等.盐胁迫对柳枝稷苗期生长和生理特性的影响[J].应用生态学报,2012,23(6):1476-1480.
FAN X F,HOU X C,ZHU Y,et al.Effects of salt stress on growth and ghysiological characteristics of switchgrass at seedling stage [J].Chinese Journal of Applied Ecology,2012,23(6):1476-1480.
[13]马晓军,金峰学,杨 姗,等.作物耐盐碱数量性状基因座(QTL) 定位[J].分子植物育种,2015,13(1):221-227.
MA X J,JIN F G,YANG SH,et al.Salt-tolerant crop quantitative trait loci(QTL) [J].Journal of Molecular Plant Breeding,2015,13(1):221-227.
[14]高彩婷,刘景辉,张玉芹,等.短期盐胁迫下燕麦幼苗的生理响应[J].草地学报,2017,25(2):337-343.
GAO C T,LIU J H,ZHANG Y Q,et al.Physiological response of oat seedlings to short-term salt stress [J].Acta Agrestia Sinica,2017,25(2):337-343.
[15]王宝山,赵可夫,邹 琦.作物耐盐机理研究进展及提高作物抗盐性的对策[J].植物学通报,1997,14(Z1):26-31.
WANG B SH,ZHAO K F,ZOU Q.Research progress of salt tolerance mechanism of crops and countermeasures to improve salt tolerance of crops [J].Botany Bulletin,1997,14(Z1):26-31.
[16]FALSTER D S,WESTOBY M.Plant height and evolutionary games[J].Trends in Ecology and Evolution,2003,18(7):337-343.
[17]KOYAMA M L,LEVESLEY A,KOEBNER R M.Ouantitative trait loci forcomponent physiological traits determining salt tolerance in rice [J].Plant Physiology,2001,125(1):406 422.
[18]王 波,张金才,宋凤斌,等.燕麦对盐碱胁迫的生理响应[J].水土保持学报,2007,21(3):86-89.
WANG B,ZHANG J C,SONG F B,et al.Physiological response of oat to salt alkali stress [J].Journal of Soil and Water Conservation,2007,21(3):86-89.
[19]范 远,任长忠,李品芳,等.盐碱胁迫下燕麦生长及阳离子吸收特征[J].应用生态学报,2011,22(11):2875-2882.
FAN Y,REN C ZH,LI P F,et al.Growth and cationic absorption characteristics of oat under saline-alkali stress [J].Chinese Journal of Applied Ecology,2011,22(11):2875-2882.
[20]王晓春,杨天辉,王 川,等.混合盐碱胁迫对紫花苜蓿的生长和生理指标的影响[J].中国农学通报,2022,38(19):139-145.
WANG X CH,YANG T H,WANG CH,et al.Effects of mixed saline-alkali stress on growth and physiological indexes of alfalfa [J].Chinese Agricultural Science Bulletin,2022,38(19):139-145.
[21]FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33(1):317-345.
[22]王旭明,赵夏夏,周鸿凯,等.NaCl胁迫对不同耐盐性水稻某些生理特性和光合特性的影响[J].热带作物学报,2019,40(5):882-890.
WANG X M,ZHAO X X,ZHOU H K,et al.Effects of NaCl stress on some physiological and photosynthetic characteristics of rice with different salt tolerance [J].Journal of Tropical Crops,2019,40(5):882-890.
[23]闫永庆,王文杰,朱 虹,等.混合盐碱胁迫对青山杨渗透调节物质及活性氧代谢的影响[J].应用生态学报,2009,20(9):2085-2091.
YAN Y Q,WANG W J,ZHU H,et al.Effects of mixed salt-alkali stress on osmotic regulatory substances and active oxygen metabolism of poplar [J].Chinese Journal of Applied Ecology,2009,20(9):2085-2091.
[24]张 婷,徐 捷,李名扬,等.4种野生百合对盐碱胁迫生理响应的初步研究[J].西南林业大学学报(自然科学),2021,41(6):22-30.
ZHANG T,XU J,LI M Y,et al.Physiological responses of 4 species of wild lily to salt-alkali stress [J].Journal of Southwest Forestry University(Natural Science),2021,41(6):22-30.
[25]李赵嘉,左永梅,宋明月,等.盐胁迫对大叶蒲公英生长生理指标及耐盐阈值的影响[J].中药材,2020,43(7):1558-1562.
LI ZH J,ZUO Y M,SONG M Y,et al.Effects of salt stress on growth physiological indexes and salt tolerance threshold of dandelion macrophylla [J].Chinese Herbal Medicine,2020,43(7):1558-1562.
[26]何 淼,王 欢,徐鹏飞,等.模拟复合盐碱胁迫对芒幼苗生理特性的影响[J].草业科学,2016,33(7):1342-1352.
HE M,WANG H,XU P F,et al.Effects of simulated combined saline-alkali stress on physiological characteristics of aemus sinensis seedlings [J].Pratacultural Science,2016,33(7):1342-1352.
[27]李玉梅,郭修武,代汉萍.牛叠肚幼苗对盐碱胁迫的生理响应及其耐盐阈值[J].西北植物学报,2014,34(6):1213-1219.
LI Y M,GUO X W,DAI H P.Physiological response of tripe tripe seedlings to salt-alkali stress and threshold of salt tolerance [J].Acta Botanica Boreali-Occidentalis Sinica,2014,34(6):1213-1219.
[28]刘 欢,张玉霞,孙明雪,等.混合盐碱胁迫对饲用高粱幼苗生理特性的影响[J].内蒙古民族大学学报(自然科学版),2022,37(2):143-148.
LIU H,ZHANG Y X,SUN M X,et al.Effects of mixed salt-alkali stress on physiological characteristics of forage sorghum seedlings [J].Journal of Inner Mongolia University for Nationalities(Natural Science Edition),2022,37(2):143-148.
[29]张喜斌,秦 楚,麻冬梅,等.紫花苜蓿苗期耐盐性鉴定[J].北方园艺,2017(22):43-50.
ZHANG X B,QIN CH,MA D M,et al.Salt tolerance identification of alfalfa seedling stage [J].Northern Horticulture,2017(22):43-50.
[30]谢 楠,赵海明,李 源,等.饲用黑麦、小黑麦品种苗期耐盐性评价及盐胁迫下的生理响应[J].草地学报,2016, 24(1):84-92.
XIE N,ZHAO H M,LI Y,et al.Salt tolerance evaluation of forage rye and triticale varieties and physiological response to salt stress at seedling stage [J].Acta Agrestia Sinica,2016,24(1):84-92.
Physiological Difference and Saline-alkali Tolerance Threshold of Oat Seedlings under Saline-alkali Stress
ZHANG Biru,MI Junzhen,ZHAO Baoping and LIU Jinghui
(National Outstanding Talents in Agricultural Research and Their Innovation Team/Inner Mongolia University
Oat Engineering Research Center/Cooperative Innovation Center of Mixed Cereal Industry,
Inner Mongolia Agricultural University, Hohhot 010019,China)
Abstract To investigate the effects of mixed salt-alkali stress on the growth and physiology of oat seedlings and to calculate the threshold of saline-alkali tolerance, two contrasting oat (Avena sativa L.) genotypes, ‘Baiyan 5’ (sensitive) and ‘Bayou 18’(tolerant), were grown under a series of salt-alkaline concentrations, and a number of growth and physiological indicators were measured. The results indicated that (1) The plant height, root length, and biomass of the genotypes were differentially affected by the stresses, and the differences were statistically significant at 150 and 200 mmol/L. (2) The levels of SPAD, Pn, Gs, and Tr in ‘Baiyan 5’ decreased as the salt-alkaline concentration increased, and the decreases became statistically significant at 100 mmol/L. In contrast, the levels of SPAD, Pn,Gs, and Tr in ‘Bayou 18’ increased at low levels of salt-alkaline concentrations before they decreased at higher levels of concentrations, and the decreases became statistically significant at 200 mmol/L. Opposite responses to the increase of salt-alkaline concentrations was found for Ci. (3) In both genotypes, the levels of proline(Pro)and Malondialdehyde(MDA)increased while that of catalase activity (CAT) decreased as the salt-alkaline concentration increased. The activities of superoxide dismutase (SOD) and peroxidase (POD) increased first to reach a maximum and then decreased as the salt-alkaline concentration increased. The maximum levels of SOD and POD were achieved at 150 and 200 mmol/L for ‘Baiyan 5’ and ‘Bayou 18’,respectively. (4) The level of salt-alkaline concentration to reduce the plant height by 50% was proposed as the salt-alkaline tolerance threshold, which was estimated as 163.4 and 200.0 mmol/L for ‘Baiyan 5’ and ‘Bayou 18’, respectively. In conclusion, the determination of oat seedling tolerance thresholds provides a theoretical basis for the cultivation and popularization of oats in salt-alkali soil.
Key words Oat; Saline-alkali stress; Physiological difference; Saline-alkali tolerance threshold
Received 2023-06-08 Returned 2023-08-01
Foundation item National Key R&D Program-Key Special Project for International Cooperation (No.2018YFE0107900); National Oat Buckwheat Industry Technology System(No.CARS-07);Science and Technology Innovation Team for Oat Whole Industry Chain(No.BR22-12-05);Inner Mongolia Oat and Quinoa Industry Technology Innovation and Promotion System.
First author ZHANG Biru, female, doctoral student. Research area:crop cultivation and farming system.E-mail:a1345639940@163.com
Corresponding author LIU Jinghui, male, Ph.D,professor,doctoral supervisor. Research area:farming system and agroecosystem. E-mail:cauljh@163.com
(责任编辑:成 敏 Responsible editor:CHENG Min)
基金项目:国家重点研发计划国际合作重点专项(2018YFE0107900);国家燕麦荞麦产业技术体系项目(CARS-07);燕麦全产业链科技创新团队(BR22-12-05);内蒙古自治区燕麦藜麦产业技术创新推广体系。
第一作者:张碧茹,女,博士研究生,主要从事作物栽培学与耕作学研究。E-mail:a1345639940@163.com
通信作者:刘景辉,男,博士,教授,博士生导师,主要从事耕作制度与农业生态系统研究。E-mail:caulih@163.com
