
灰毡毛忍冬ERF 基因家族鉴定及其在衰老过程中的表达分析
2024-09-29 00:00:00刘燕妮曹正艳吴佩荫李清清陈泽雄唐宁
经济林研究 2024年2期


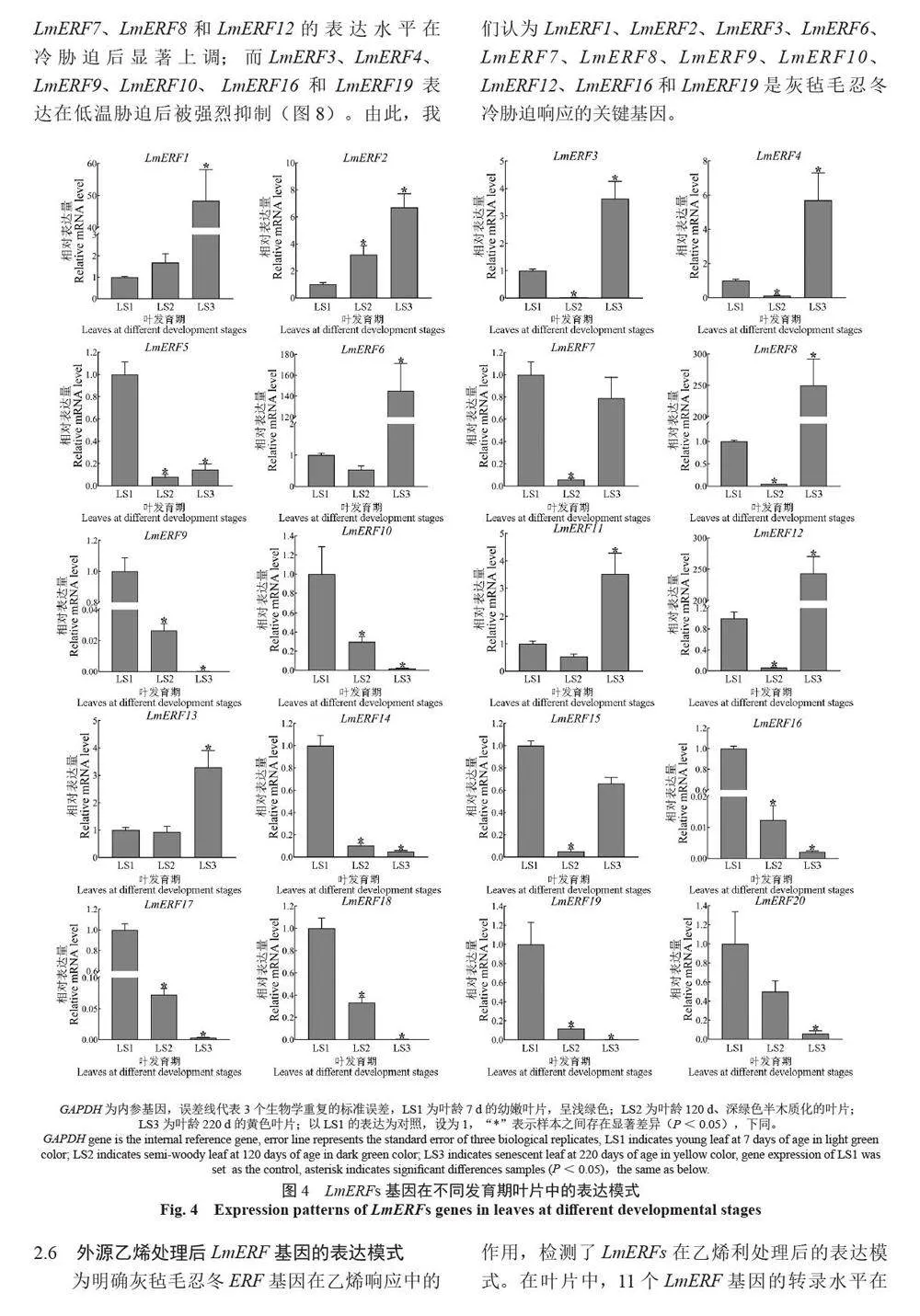

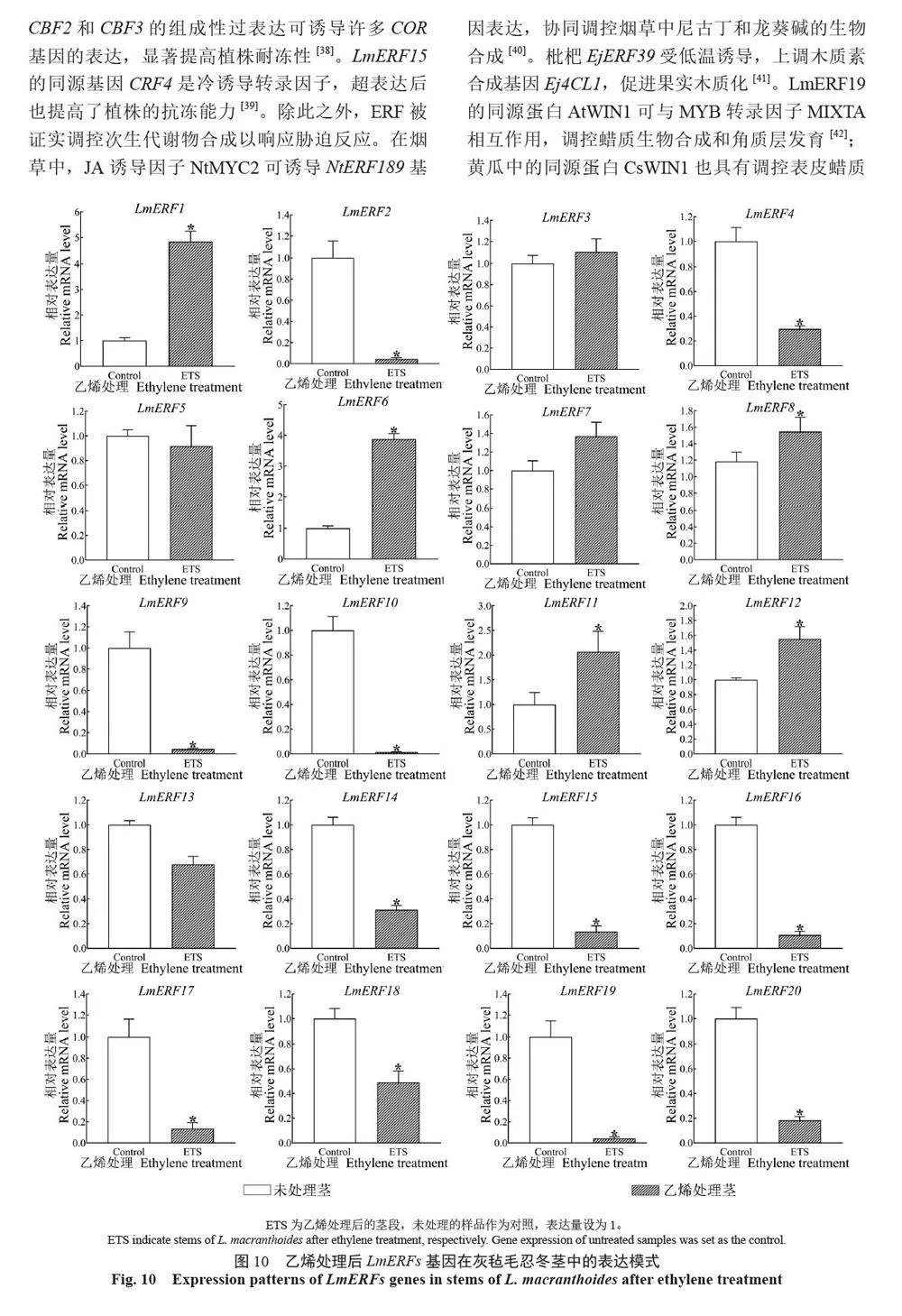









摘 要:【目的】灰毡毛忍冬Lonicera macranthoides 为我国南方地区大宗道地中药材和重要经济植物,绿原酸和木犀草苷等药效物质在器官衰老过程中积累且受转录因子调控。乙烯信号转导相关ERF 基因家族广泛参与植物的衰老、胁迫响应和次生代谢。鉴定灰毡毛忍冬ERF 家族成员并探究其表达模式,可为该类基因调控次生代谢的功能研究提供参考。【方法】基于转录组数据鉴定灰毡毛忍冬ERF 转录因子家族,通过系统发育树分析其进化关系,利用MEME 在线网站分析序列的结构基序。QRT-PCR 分析LmERFs 基因在不同器官衰老过程以及冷胁迫和乙烯处理后的表达模式。【结果】在灰毡毛忍冬中共鉴定出65 个具AP2 保守结构域的LmERFs 基因,可将其分为3 个亚组。利用RNA-Seq 技术筛选了在幼嫩和衰老叶片之间差异表达的20 个LmERFs。进一步qRT-PCR 分析显示,分别有18 个、13 个和11 个LmERFs 基因在叶片、茎和花的衰老过程显著差异表达,其中LmERF2、LmERF9、LmERF16、LmERF17 和LmERF19 基因表达在不同器官衰老过程中均具显著差异。冷胁迫后,叶和茎中7 个LmERFs(LmERF1、LmERF2、LmERF6、LmERF7、LmERF8、LmERF11 和LmERF12) 显著上调,LmERF3、LmERF9、LmERF16 和LmERF19 显著下调;而乙烯处理诱导了LmERF1、LmERF8 和LmERF12基因的表达,抑制了LmERF2、LmERF4、LmERF9、LmERF10、LmERF14、LmERF15、LmERF16、LmERF17、LmERF18 和LmERF19 基因的表达。【结论】8 个LmERFs(LmERF1、LmERF2、LmERF6、LmERF9、LmERF10、LmERF15、LmERF16 和LmERF19)响应了器官发育、冷胁迫和乙烯诱导的衰老过程,这为后续挖掘高效调控灰毡毛忍冬次生代谢的靶点LmERFs 提供了数据支持。
关键词:灰毡毛忍冬;ERF 基因家族;器官衰老;乙烯;冷害胁迫
中图分类号:S567.79 文献标志码:A 文章编号:1003—8981(2024)02—0013—16
灰毡毛忍冬Lonicera macranthoides Hand-Mazz又名大金银花,忍冬科忍冬属植物,为我国西南地区大宗道地中药材和特色经济植物,其次生代谢产物如绿原酸和木犀草苷等核心药效成分具有抗病毒、抗氧化、增强免疫力等多种功能[1]。目前,灰毡毛忍冬已被广泛应用于制药、保健品及化妆品等行业,市场巨大[2]。然而,灰毡毛忍冬存在品种混杂、药效成分含量参差不齐等问题,已成为制药、提取、加工等下游产业发展的瓶颈,挖掘调控次生代谢产物生物合成的关键因子是解决这一难题的重要途径。
研究表明,植物通过产生次生代谢物响应衰老信号、适应环境压力[3]。拟南芥叶片衰老过程中,花青素和吲哚衍生的甲基葡萄糖苷显著增加;而黄酮醇苷和芥子酸共轭物持续减少。随着器官的发育和衰老,灰毡毛忍冬绿原酸、绿原酸前体及异构体等苯丙烷类物质和木犀草苷等黄酮类物质含量发生显著变化[1,4]。Rezaie 等[5] 发现,冷胁迫显著影响了罗勒中苯丙素类化合物的含量。糖和次生代谢在柑橘类植株响应冷胁迫中发挥着重要作用[6-7]。乙烯可促进植物次生代谢物的产生,如长春花中酚酸、黄酮醇、萜类和吲哚类生物碱、乌龙茶中儿茶素和甜橙中瓦伦西亚橘烯[8]。由此可知,植物发育、激素或胁迫信号显著影响次生代谢物合成,探究灰毡毛忍冬响应衰老和胁迫机制有助于挖掘次生代谢物合成的关键基因,为利用基因工程手段培育新品种提供理论依据。
转录因子通过介导次生代谢物合成参与生物和非生物应激反应[9]。AP2/ERF 转录因子是植物最大的转录因子家族之一,成员含有1 个或2 个由60 ~ 70 个氨基酸构成的AP2 保守结构域,根据结构域数量和识别序列的不同可将其分为AP2、ERF、DREB、RAV 和Soloist5 个亚家族[10],广泛参与植物生长发育、初生和次生代谢调控、生物或非生物胁迫响应[11] 以及乙烯等植物激素信号传导途径[12-13]。其中ERF(ethylene response factor) 转录因子亚家族是乙烯信号通路下游的关键调控因子[14-15],只含有一个AP2 结构域,第14 位和第19 位氨基酸分别是丙氨酸和天冬氨酸,区别于其他亚家族成员。ERF 亚家族成员可特异性结合基因启动子区的GCC-box 元件(AGCCGCC),从而参与调控乙烯应答、生物及非生物胁迫和次生代谢[13,16]。胡俊东等[17] 研究表明,PheERF5 基因参与毛竹抗生物和非生物胁迫过程;小麦TaERF1基因在拟南芥中过表达可提高转基因植株对高盐和低温的耐受性[18];ERF- Ⅵ亚族成员CRF 具有正向调节植物渗透胁迫耐受性和抗冻性作用[18-19]。Lyu 等[20] 发现桦木中BpERF13 过表达可通过结合并上调冷胁迫关键转录因子CBF 的基因表达来增强耐寒性。拟南芥ERF 亚族转录因子ORA59基因受茉莉酸和乙烯的协同诱导,通过正向调控PDF1.2 的表达增加植物灰霉抗性,在茉莉酸与乙烯信号的交叉对话中起了重要的调控作用[21-22]。烟草中过表达NtERF19 可显著增加生物碱,特别是新烟碱的含量[23]。目前,鲜见忍冬属药用植物ERF 转录因子的研究,在灰毡毛忍冬中尚未见ERF 参与胁迫响应的研究报道。
