
EMS诱变的水稻不同位点突变体对ALS抑制剂类除草剂抗性差异
2024-09-26 00:00:00周佳伟王歆凯唐兆成凌溪铁张保龙
江苏农业学报 2024年7期


摘要:稻田杂草严重威胁水稻生长,培育抗除草剂水稻新品种,是高效解决稻田杂草危害的重要途径之一。本研究利用甲基磺酸乙酯(EMS)对华占水稻进行化学诱变,创建突变体,利用乙酰乳酸合成酶(ALS)抑制剂类除草剂甲咪唑烟酸(20 g/hm2,有效成分)对突变体进行筛选,获得抗甲咪唑烟酸突变体材料,并对上述突变体的OsALS基因序列进行分析,明确其突变类型。最后对不同突变类型对甲氧咪草烟和甲基二磺隆的交互抗性进行分析。结果表明,华占水稻经EMS诱变和甲咪唑烟酸筛选后共获得11株抗甲咪唑烟酸突变体材料,其突变类型为Pro171Leu、Ser627Cys、Ser627Asn和Gly628Glu 4种。其中,Ser627Cys为首次发现的突变类型。4种突变类型对甲咪唑烟酸的抗性水平存在一定的差异,Ser627Asn、Ser627Cys、Pro171Leu、Gly628Glu突变类型的抗性指数分别为8.71、5.16、3.28和3.69。4种突变类型对甲氧咪草烟和甲基二磺隆的抗性亦存在显著差异。其中,Ser627Asn突变类型对甲氧咪草烟的抗性最高,抗性指数为20.57,Gly628Glu对甲基二磺隆抗性最高,抗性指数为5.11。综上,本研究通过EMS诱变获得了4种抗ALS抑制剂类除草剂的华占水稻突变类型,并鉴定了其对甲咪唑烟酸的抗性差异及对甲氧咪草烟和甲基二磺隆的交互抗性,为进一步的抗除草剂水稻品种培育和除草剂轮换使用提供依据。
关键词:水稻;ALS抑制剂类除草剂;EMS诱变;抗除草剂突变体
中图分类号:S511.035.2文献标识码:A文章编号:1000-4440(2024)07-1153-08Differences in ALS inhibitor herbicide resistance among rice mutants with different loci induced by EMS mutagenesisZHOU Jiawei WANG Xinkai TANG Zhaocheng LING Xitie ZHANG Baolong
(1.College of Tropical Agriculture and Forestry, Hainan University, Haikou 570228, China;2.Institute of Germplasm Resources and Biotechnology, Jiangsu Academy of Agricultural Sciences/Jiangsu Provincial Key Laboratory of Agrobiology, Nanjing 210014, China)
Abstract:Weeds in paddy fields seriously threaten the growth of rice. Breeding new varieties of herbicide-resistant rice is one of the important ways to effectively solve the harm of weeds in paddy fields. In this study, ethyl methane sulfonate (EMS) was used as a chemical mutagen to induce Huazhan rice to create mutants. The mutants were screened by the acetolactate synthase (ALS) inhibitor herbicide imazapic (20 g/hm2) to obtain resistant mutant materials. The OsALS gene sequences of the above mutants were analyzed to clarify the mutation type. Finally, the cross-resistance of different mutant types to imazamox and mesosulfuron-methyl was analyzed. The results showed that 11 imazapic-resistant mutants were obtained from Huazhan rice by EMS mutagenesis and imazapic screening. The mutation types were Pro171Leu, Ser627Cys, Ser627Asn and Gly628Glu. Among them, Ser627Cys mutation type was first reported. The resistance levels of the four mutation types to imazapic were different. The resistance indices of Ser627Asn, Ser627Cys, Pro171Leu and Gly628Glu were 8.71, 5.16, 3.28 and 3.69, respectively. There were also significant differences in the resistance of the four mutation types to imazamox and mesosulfuron-methyl. The resistance of Ser627Asn to imazamox was the highest, and the resistance index was 20.57. The resistance of Gly628Glu to mesosulfuron-methyl was the highest, and the resistance index was 5.11. In summary, four Huazhan rice mutant types resistant to ALS inhibitor herbicides were obtained by EMS mutagenesis, and their resistance differences to imazapic and the cross-resistance to imazamox and mesosulfuron-methyl were identified, which provided a basis for further cultivation of herbicide-resistant rice varieties and rotation of herbicides.
Key words:rice;ALS inhibitor herbicide;EMS mutagenesis;herbicide-resistant mutant
水稻(Oryza sativa L.)是全球最重要的粮食作物之一。稻田杂草与水稻争夺生长发育所需的营养成分是制约水稻生产的重要生物因子,且同为稻属的杂草稻不受现有稻田除草剂影响,严重影响水稻的产量和品质[1-2]。目前稻田杂草防除主要通过化学除草的方法实现。化学除草具有效率高、成本低的特点,但单一除草剂的长期使用容易导致杂草产生抗药性,同时除草剂的过量使用还会对水稻造成药害[3]。目前生产中,常通过除草剂的轮换和混合使用延缓杂草对除草剂抗性的产生。近年来,抗除草剂水稻种质的培育为稻田杂草的防除提供了新的手段[4-5]。
缬氨酸、亮氨酸和异亮氨酸等支链氨基酸(Branched chain amino acids,BCAA)是植物生长发育必需的氨基酸,乙酰乳酸合酶(ALS)是合成BCAA的关键酶[6]。ALS抑制剂类除草剂能与植物ALS形成复合物,阻碍其与底物结合,抑制ALS活性,从而减少支链氨基酸的合成,最终导致植物死亡。ALS抑制剂类除草剂主要包括咪唑啉酮类(Imidazolidinones,IMI)、磺酰脲类(Sulfonylureas,SU)、三唑并嘧啶类(Triazole pyrimidines,TP)、嘧啶水杨酸类(Pyrimidinylthio-benzoates,PTB)、磺酰胺基羰基三唑并嘧啶类(Sulfonylamino-carbonyltriazolinones,SCT)[7-8]。
ALS氨基酸突变能使植物获得对ALS抑制剂类除草剂的抗性。水稻ALS基因第96、122、171、197、548、627和628位点突变能使水稻对ALS抑制类除草剂产生抗性[9-10]。Croughan[11]和Powles等[12]通过化学诱变AS3510和Cypress 2个水稻品种,分别用咪唑乙烟酸和甲咪唑烟酸对M2代进行筛选,获得ALS基因第627位点突变(编码的氨基酸由丝氨酸突变为天冬酰胺)的抗除草剂水稻品种CL121、CL141和第628位点突变(编码的氨基酸由甘氨酸突变为谷氨酸)的抗除草剂水稻品种CL161、XL8,并用于商业化生产。Shoba等[13]通过EMS诱变旱稻品种N22,获得第152位点突变(编码的氨基酸由甘氨酸突变为谷氨酸)的抗咪唑啉酮类除草剂咪草烟材料HTM-N22。Gealy等[14]通过EMS诱变水稻品种IRGA417,获得第96位点突变(编码的氨基酸由丙氨酸突变为苏氨酸)的抗咪唑啉酮类除草剂水稻品种PUIT INTA CL。Okuzaki等[15]利用双草醚筛选水稻花药衍生的愈伤组织,获得第95位点突变(编码的氨基酸由甘氨酸突变为丙氨酸)的抗磺酰脲类除草剂水稻品种。Chen等[16]通过EMS诱变水稻品种黄华占获得第548位点、第96位点、第627位点、第628位点突变的抗除草剂水稻系列品种,其中,第548位点突变的材料经过回交和除草剂筛选获得能稳定遗传的抗除草剂水稻品种JTD-001。Piao等[17]利用CRISPR/Cas9技术定点突变南粳9108的OsALS基因,获得第628位点突变(编码的氨基酸由甘氨酸突变为色氨酸)的抗除草剂水稻品种;王芳权等[18]利用除草剂对7 000多个水稻品种进行筛选,获得1个抗咪唑乙烟酸除草剂水稻品种金粳818,其ALS基因突变类型为Ser627Asn。
目前对于抗ALS抑制剂类除草剂水稻的研究主要集中在材料创制方面,而同一突变位点对不同ALS抑制剂类除草剂是否存在交互抗性、同一遗传背景下的ALS不同突变位点对ALS抑制剂类除草剂的抗性是否存在差异的研究较少。本研究利用甲基磺酸乙酯(EMS)对恢复系水稻品种华占进行化学诱变,构建突变体库,并利用甲咪唑烟酸对突变体库进行筛选,获得抗甲咪唑烟酸突变体材料,并对上述突变体的OsALS基因序列进行分析,明确其突变类型。最后分析不同类型突变体对甲咪唑烟酸、甲氧咪草烟、甲基二磺隆的抗性,为抗ALS抑制剂类除草剂水稻种质的培育和除草剂轮换使用提供依据。
1材料与方法
1.1材料与试剂
供试水稻材料为籼稻品种华占(品种权号:CNA20080059.0)。化学诱变剂甲基磺酸乙酯(EMS)购自Sigma-aldrich公司,4%甲氧咪草烟(咪唑啉酮类除草剂)购自江苏省农用激素工程技术研究中心有限公司,30 g/L甲基二磺隆(磺酰脲类除草剂)购自富美实农业科技有限公司,240 g/L甲咪唑烟酸(咪唑啉酮类除草剂)购自德国巴斯夫公司。DNA提取和PCR扩增所用试剂购自南京诺维赞科技有限公司,引物由南京金斯瑞生物科技有限公司合成,DNA测序由南京擎科生物科技有限公司完成。
1.2抗甲咪唑烟酸除草剂水稻突变体筛选
华占突变体种子库的构建:选取颗粒饱满华占水稻种子200 kg晾晒后用清水浸种2 h,再用质量浓度5 g/L的EMS溶液常温浸种14 h,然后加入25 g/L的硫代硫酸钠进行中和反应15 min,倒掉废液,用自来水清洗种子5次,每次清洗5 min。EMS诱变处理后的种子按60 kg/hm2播种量直播在大田,正常肥水管理,于成熟期混收种子,收获的种子即为华占突变体种子库。
从EMS诱变获得的华占突变体种子库中随机选取243 kg种子,以900 kg/hm2高密度撒播于大田苗床,播种面积为0.27 hm2。于秧苗3叶期全田喷施有效成分为20 g/hm2的甲咪唑烟酸,喷药后14 d观察苗床植株存活情况,生长正常的植株作为具有抗性的突变体单株移栽至温室,单株收种用于后续试验。
1.3抗除草剂水稻突变体OsALS基因序列分析
单株剪取筛选得到的突变体及野生型华占植株叶片1 g,使用DNA试剂盒提取突变体及野生型DNA。根据在NCBI(美国国家生物信息中心)网站上检索得到的水稻OsALS基因保守序列,设计特异性引物 OsALS-F和OsALS-R,引物序列分别为5′-TCGCCCAAACCCAGAAACCC-3′和5′-CTCTTTATGGGTCATTCAGGTC-3′。利用B-96型梯度PCR仪(杭州博日科技股份有限公司产品)对突变体和野生型植株的OsALS基因进行PCR扩增,PCR扩增体系为:DNA 1.0 μL、引物OsALS-F、OsALS-R各1.0 μL、Phanta Max 高保真 DNA 聚合酶1.0 μL、2×Phanta Max Buffer 25.0 μL、dNTP Mix(10 mmol/L)1.0 μL、5% 二甲基亚砜(DMSO)2.5 μL、双蒸水(ddH2O)补至50.0 μL。扩增程序为预变性95 ℃ 2 min;变性95 ℃ 15 s,退火58 ℃ 15 s ,延伸72 ℃ 2 min,35个循环。扩增产物送南京擎科生物科技有限公司测序,采用SnapGene 4.1.8软件对测序结果进行分析比对。
1.4抗性突变体对甲咪唑烟酸的抗性鉴定
参照文献[19]的方法对突变体和野生型水稻对甲咪唑烟酸的抗性水平测定。选取均匀一致的突变体种子、野生型水稻种子,播种到盛有土壤的穴盘中,每穴播种10粒。在水稻2叶期间苗,每穴保留生长一致的幼苗5株。于水稻3叶期时,利用3WP-2000型行走式喷雾塔进行甲咪唑烟酸喷雾处理,喷液压力0.275 mPa,喷液量450 L/hm2。野生型水稻喷施甲咪唑烟酸剂量(有效成分)分别为0 g/hm2、14.4 g/hm2、21.6 g/hm2、32.4 g/hm2、48.6 g/hm2、72.6 g/hm2、109.4 g/hm2 、164.0 g/hm2 8个处理,突变体喷施甲咪唑烟酸剂量(有效成分)分别为0 g/hm2、14.4 g/ hm2、21.6 g/hm2、32.4 g/hm2、48.6 g/hm2、72.6 g/hm2、109.4 g/hm2、164.0 g/hm2、246.0 g/hm2、369.1 g/hm2 、553.6 g/hm2共11个处理,每个处理4个重复。施药后21 d时剪取各处理的植株地上部存活组织,称取鲜重[20]。
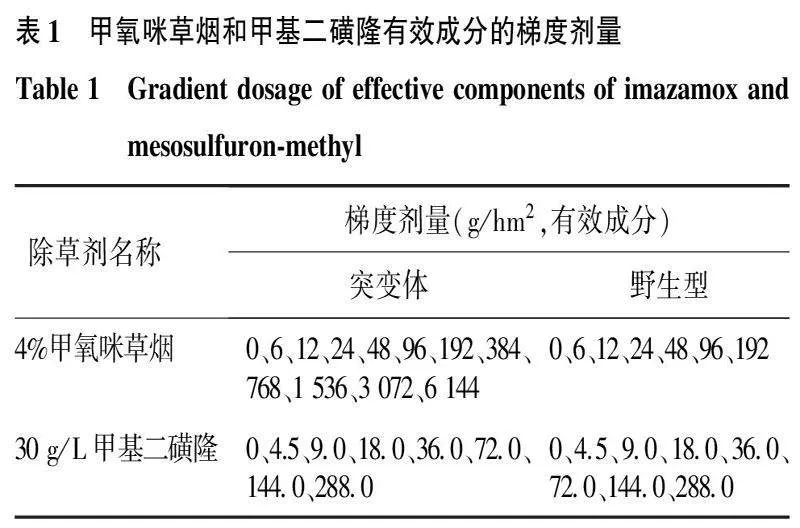
1.5抗性突变体和野生型植株对甲氧咪草烟和甲基二磺隆的交互抗性鉴定参照抗性突变体对甲咪唑烟酸的抗性水平测定方法,于水稻3叶期开展突变体和野生型水稻对甲氧咪草烟和甲基二磺隆的交互抗性鉴定。甲氧咪草烟和甲基二磺隆有效成分的梯度处理如表1所示。每个处理设4个重复。
1.6数据处理及分析
利用Sigmaplot 14. 0软件进行图、表的制作。利用Logistic方程拟合不同处理下突变体和野生型水稻除草剂剂量与鲜重抑制率的关系,进行鲜重抑制中量(GR50)的计算,并根据的突变体材料的GR50与野生型水稻的GR50比值,计算不同突变体材料的抗性指数(RI),以明确突变体水稻对除草剂的抗性强弱。
2结果与分析
2.1抗性突变体的筛选获得
华占突变体群体喷施甲咪79b35e2446514cb1067fa0eb099b377e唑烟酸后,植株生长受抑制,14 d后非抗性植株基本死亡,本次筛选共得到11个存活单株,如图1所示。对存活单株进行移栽和单株收种,用于进一步的突变类型及除草剂抗性鉴定试验。
2.2抗性突变体OsALS基因序列分析
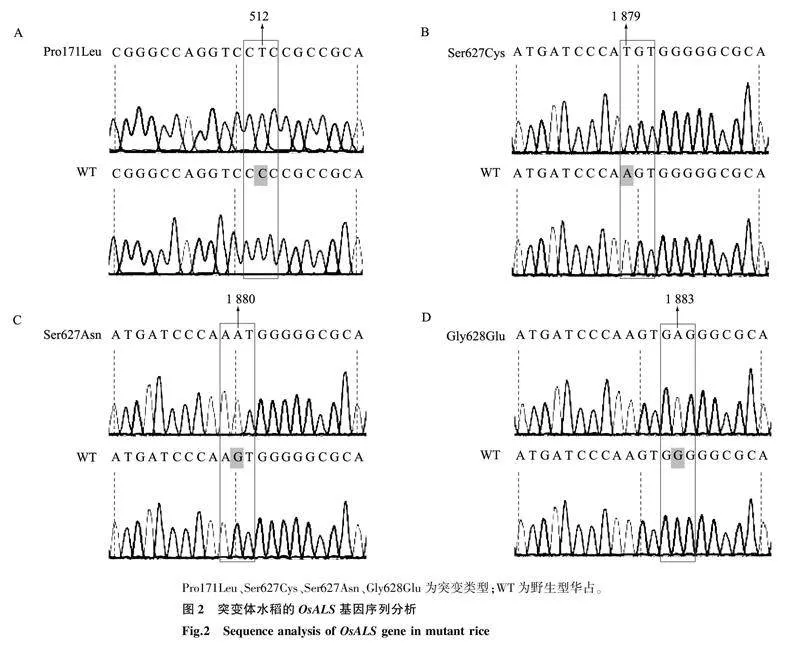
获得的11个抗性突变体与野生型OsALS基因序列差异结果如图2所示。11个突变体单株OsALS基因的突变类型有4种,其中有3个抗性单株的OsALS基因第512碱基由C突变为T,1个抗性单株的OsALS基因第1 879碱基由A突变为T,4个抗性单株的OsALS基因第1 880碱基由G突变为A,3个抗性单株的OsALS基因第1 883碱基由G突变为A。11个突变体对应的氨基酸突变类型分别为Pro171Leu、Ser627Cys、Ser627Asn、Gly628Glu 4种。其中,Ser627Cys为本研究发现的新突变类型。
2.3抗性突变体对甲咪唑烟酸的抗性水平
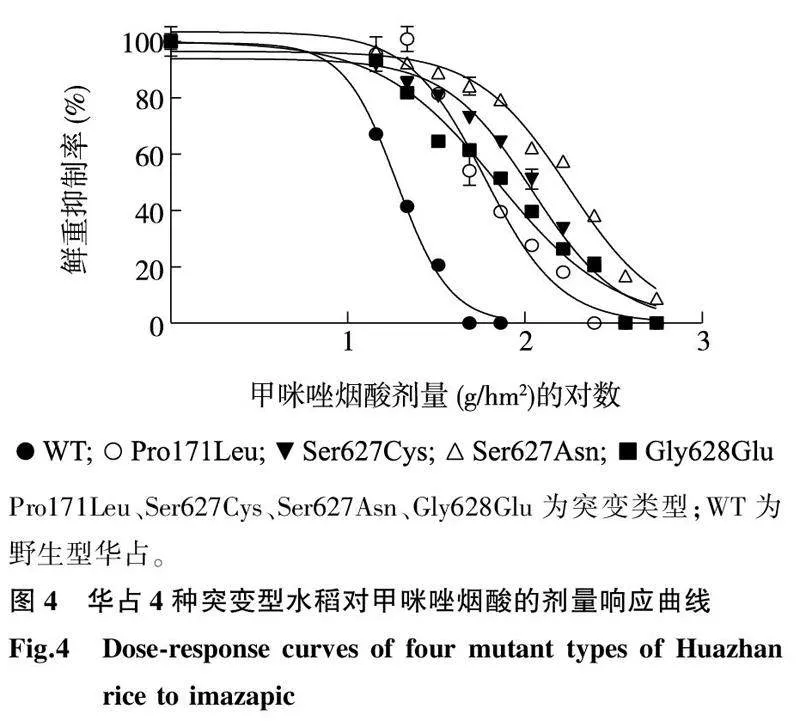
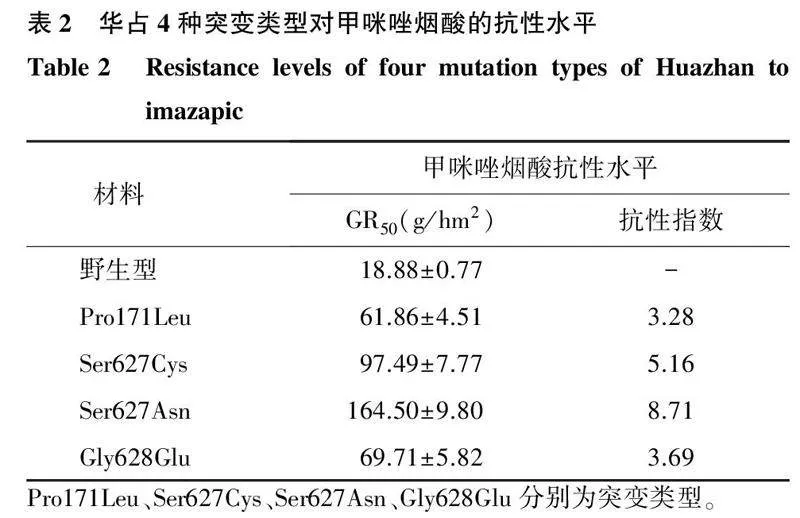
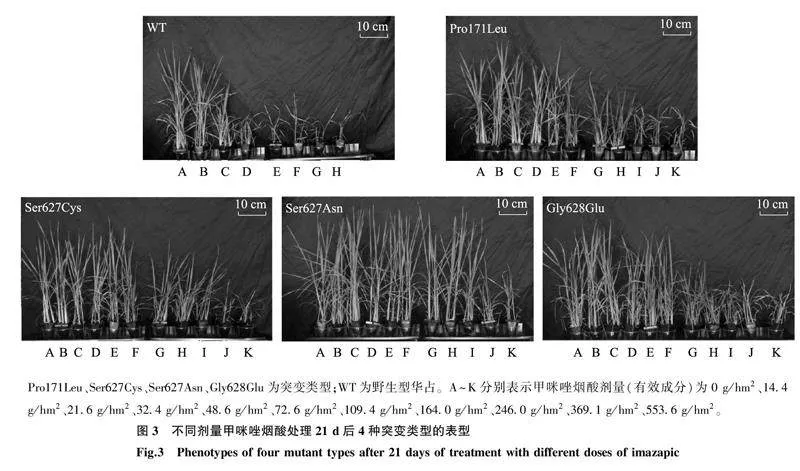
与野生型相比, 4种华占ALS突变类型均对甲咪唑烟酸表现出明显的抗性(图3、图4、表2)。野生型华占对甲咪唑烟酸的GR50值为18.88 g/hm2,Pro171Leu突变类型对甲咪唑烟酸的GR50值为61.86 g/hm2;Ser627Cys突变类型对甲咪唑烟酸的GR50值为97.49 g/hm2;Ser627Asn突变类型对甲咪唑烟酸的GR50值为164.50 g/hm2;Gly628Glu突变类型对甲咪唑烟酸的GR50值为69.71 g/hm2。4种突变类型中,以Ser627Asn突变类型对甲咪唑烟酸的抗性最好,抗性指数为8.71;其次是Ser627Cys突变类型,抗性指数为5.16;Pro171Leu与Gly628Glu两种突变类型对甲咪唑烟酸的抗性水平相当,抗性指数分别为3.28和3.69。
Pro171Leu、Ser627Cys、Ser627Asn、Gly628Glu为突变类型;WT为野生型华占。A~K分别表示甲咪唑烟酸剂量(有效成分)为0 g/hm2、14.4 g/hm2、21.6 g/hm2、32.4 g/hm2、48.6 g/hm2、72.6 g/hm2、109.4 g/hm2、164.0 g/hm2、246.0 g/hm2、369.1 g/hm2、553.6 g/hm2。
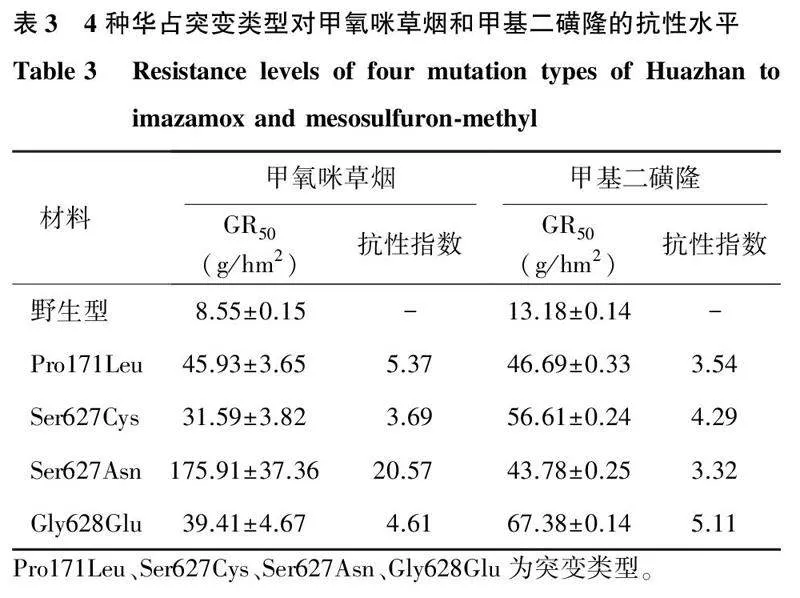
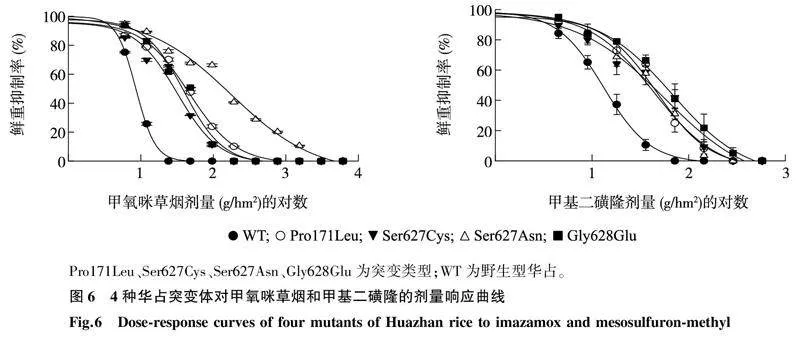
2.44种抗性突变体对甲氧咪草烟和甲基二磺隆的交互抗性与野生型相比, 4种华占ALS突变类型均对甲氧咪草烟和甲基二磺隆表现出不同的抗性水平(如图5、图6和表3)。野生型华占对甲氧咪草烟和甲基二磺隆的GR50值分别为8.55 g/hm2和13.18 g/hm2,Pro171Leu突变类型对甲氧咪草烟和甲基二磺隆的GR50值分别为45.93 g/hm2和46.69 g/hm2;Ser627Cys突变类型对甲氧咪草烟和甲基二磺隆的GR50值分别为31.59 g/hm2和56.61 g/hm2; Ser627Asn突变类型对甲氧咪草烟和甲基二磺隆的GR50值分别为175.91 g/hm2和43.78 g/hm2; Gly628Glu突变类型对甲氧咪草烟和甲基二磺隆的GR50值分别为39.41 g/hm2和67.38 g/hm2。4种突变类型中,Ser627Asn突变类型对甲氧咪草烟的抗性最高,抗性指数为20.57;其次是Pro171Leu和Gly628Glu突变类型,抗性指数为5.37和4.61;Ser627Cys突变类型对甲氧咪草烟的抗性水平最低,抗性指数为3.69。甲基二磺隆喷施处理下,Gly628Glu突变类型的抗性最高,抗性指数为5.11,其次为Ser627Cys突变类型,抗性指数为4.29;Pro171Leu与Ser627Asn两种突变类型对甲基二磺隆的抗性水平相当,抗性指数分别为3.54和3.32。
3讨论
ALS抑制剂类除草剂是一种低毒、高效的除草剂,常用于水稻、小麦、玉米等作物中各类杂草的防治。但田间长期使用单一除草剂会导致抗除草剂杂草大量产生,严重危害作物的生长发育以及产量。不同类型除草剂的轮换使用对于延缓抗除草剂杂草产生和控制抗除草剂杂草进化具有重要意义。甲氧咪草烟属于咪唑啉酮类(IMI)除草剂,主要用于大豆田,甲基二磺隆是磺酰脲类(SU)除草剂,主要用于小麦田,这2种除草剂主要用于苗后除草,可以防除多种禾本科以及阔叶类杂草,目前这2种除草剂并未在稻田杂草上登记使用[21-22]。本研究通过检验抗除草剂水稻突变体对这2种除草剂的交互抗性水平为这2种除草剂在水稻田上的使用以及抗除草剂水稻育种提供一定的理论依据。
随着除草剂的大量应用,研究人员对抗除草剂水稻新品种的创制越发重视。水稻对除草剂产生耐受性的机制主要包括2个方面:靶点抗性(Target-site resistance,TSR)和非靶点抗性(Nontarget-site resistance,NTSR)。目前,抗除草剂水稻新品种的研究主要集中于TSR[23]。本研究利用除草剂对EMS诱变的M2代水稻进行筛选,获得4种抗除草剂突变类型,并对其相关OsALS基因序列进行克隆分析,鉴定到Pro171Leu、Ser627Cys、Ser627Asn、Gly628Glu 4种ALS突变类型。其中,Pro171Leu、Ser627Asn、Gly628Glu 3种突变类型已有报道,而Ser627Cys为新发现的突变类型[4]。Powles等[12]研究认为水稻对除草剂产生耐受性与ALS活性变化有关,即基因突变导致ALS结构发生变化,从而使得植株与除草剂的结合能力有所改变。由于不同类型的ALS抑制剂与酶的结合位点不同,导致不同突变位点对于不同类型ALS抑制剂类除草剂耐受性具有差异。Ser627Asn突变类型对IMI类除草剂的耐受性更强,而Gly628Glu突变类型对SU类除草剂的耐受性更强[10,24-25],这与本研究的结果基本一致,但这4种突变类型的ALS活性差异以及突变位点导致的酶结构差异还需进一步进行研究。
目前使水稻产生除草剂抗性的主要手段包括化学诱变、转基因及基因编辑技术[26-27]。转基因技术可以赋予作物新性状,在植物抗逆性研究中广泛应用。Dong等[28]将荧光假单胞菌(Pseudomonas fluorescens)中的G2-EPSPS基因转入水稻中花11中,转基因水稻对草甘膦的耐受性比野生型提高100倍以上。但由于转基因作物对人体和环境的安全性还缺乏全面认证,一定程度上限制了转基因作物的推广。通过化学诱变得到的水稻突变体,比转基因水稻更加安全,接受程度更高。目前,一些通过诱变获得的水稻突变体材料已经得到推广并用于商业化种植,如ALS突变类型Ser627Asn、Gly628Glu等。这些水稻品种与其他一些商业化非转基因抗除草剂作物是Clearfield生产系统的基础[29]。Clearfield生产系统中通过非转基因抗除草剂作物品种和对应抗性的广谱除草剂相结合应用,可更加高效地控制田间杂草,防止除草剂残留对作物造成的伤害,保证水稻的产量和质量。本研究创制的抗除草剂水稻突变体来源于EMS诱变,无外源基因插入,可以为Clearfield生产系统提供新的抗除草剂水稻材料,为水稻生产和杂草管理提供新的种质资源。
4结论
本研究利用化学诱变华占水稻,创建突变体,并用甲咪唑烟酸筛选获得Pro171Leu 、Ser627Cys、Ser627Asn和Gly628Glu 4种突变类型。除草剂抗性水平试验结果表明,4种突变类型中以Ser627Asn突变类型对IMI类除草剂甲氧咪草烟的抗性最高,而Gly628Glu突变类型对SU类除草剂甲基二磺隆的抗性最高。本研究结果为培育抗除草剂水稻品种提供新的种质资源,对稻田杂草的高效防治,稻田效益提高具有重要作用。
参考文献:
[1]孙学海. 稻田杂草综合防治技术[J]. 现代农业科技,2011 (16):146,148.
[2]ALLARD J L, KEEFUI K, MORISHIMA Y, et al. The crop protection industry’s view on trends in rice crop establishment in Asia and their impact on weed management techniques[C]//TORIYAMA K. Rice is life:scientific perspectives for the 21st century. Proceedings of the World Rice Research Conference 2004. Manila:IRRI,2004:205-208.
[3]伍龙梅,张悦,刘妍,等. 直播稻研究进展及发展对策分析[J]. 中国农学通报,2023,39(6):1-5.
[4]吴云雨,肖宁,余玲,等. 我国抗除草剂水稻种质创制研究进展[J]. 植物遗传资源学报,2021,22(4):890-899.
[5]李香菊. 我国转基因耐除草剂作物研发与应用[J]. 现代农药,2023,22(1):5-10.
[6]任洪雷. 乙酰乳酸合成酶及ALS基因研究概述[J]. 中国农学通报,2016,32(26):37-42.
[7]张红磊,李军玲,张融雪,等. ALS以及ALS抑制剂类除草剂的研究进展[J]. 植物学研究,2021,10(6):11.
[8]陈涛,张善磊,赵凌,等. ALS抑制剂类除草剂抗性水稻功能标记的开发与验证[J]. 中国水稻科学,2018,32(2):137-145.
[9]GALILI G, AMIR R, FERNIE A R. The regulation of essential amino acid synthesis and accumulation in plants[J]. Annual Review of Plant Biology,2016,67(1):153-178.
[10]刘长乐,郭月,李芳芳,等. 抗ALS类除草剂作物种质创制与利用研究进展[J]. 植物遗传资源学报,2022,23(2):333-345.
[11]CROUGHAN T P. Herbicide Resistant Rice:United States:20080167186[P]. 2008-07-10.
[12]POWLES S B, YU Q. Evolution in action:plants resistant to herbicides[J]. Annual Review of Plant Biology,2010,61:317-347.
[13]SHOBA D, RAVEENDRAN M, MANONMANI S, et al. Development and genetic characterization of a novel herbicide (Imazethapyr) tolerant mutant in rice (Oryza sativa L.)[J]. Rice (New York),2017,10(1):10.
[14]GEALY D, MITTEN D, RUTGER J. Gene flow between red rice (Oryza sativa) and herbicide-resistant rice (O. sativa):implications for weed management[J]. Weed Technology,2003,17:627-645.
[15]OKUZAKI A, SHIMIZU T, KAKU K, et al. A novel mutated acetolactate synthase gene conferring specific resistance to pyrimidinyl carboxy herbicides in rice[J]. Plant Molecular Biology,2007,64(1/2):219-224.
[16]CHEN L, GU G, WANG C, et al. Trp548Met mutation of acetolactate synthase in rice confers resistance to a broad spectrum of ALS-inhibiting herbicides[J]. The Crop Journal,2021,9(4):750-758.
[17]PIAO Z, WANG W, WEI Y, et al. Characterization of an acetohydroxy acid synthase mutant conferring tolerance to imidazolinone herbicides in rice (Oryza sativa)[J]. Planta,2018,247(3):693-703.
[18]王芳权,杨杰,范方军,等. 水稻抗咪唑啉酮类除草剂基因ALS功能标记的开发与应用[J]. 作物学报,2018,44(3):324-331.
[19]中华人民共和国农业部. 农药室内生物测定试验准则-(除草剂)第4 部分活性测定试验茎叶喷雾法:NY/T 1155.4-2006[S]. 北京:中国农业出版社,2006.
[20]张丽雅,李奇,史珊珊,等. 稻田稗草对五氟磺草胺的抗性机制及其防治药剂筛选[J]. 中国农业科学,2023,56(14):2713-2723.
[21]亦冰. 豆类作物除草剂——甲氧咪草烟铵盐[J]. 世界农药,2006,28(4):52,46.
[22]赵波. 甲基二磺隆的前世今生[J]. 农药市场信息,2020(22):36.
[23]邓丽琴,祝朋芳,陈长青. 试论常规育种与分子育种的研究应用[J]. 园艺与种苗,2004,24(5):280-281.
[24]YU Q, POWLES S B. Resistance to AHAS inhibitor herbicides:current understanding[J]. Pest Management Science,2014,70(9):1340-1350.
[25]MCCOURT J A, PANG S S, KING-SCOTT J, et al. Herbicide-binding sites revealed in the structure of plant acetohydroxyacid synthase[J]. Proceedings of the National Academy of Sciences of the United States of America,2006,103(3):569-573.
[26]李凌燕,肖冰,张旭冬,等. 转基因耐除草剂玉米MON87419品系特异性定性PCR检测方法的建立[J]. 江苏农业科学,2023,51(1):50-57.
[27]邹拓,杜琪,耿雷跃,等. 抗除草剂水稻耐药性及后代筛选方法的研究[J]. 江苏农业科学,2022,50(13):136-140.
[28]DONG Y, JIN X, TANG Q, et al. Development and event-specific detection of transgenic glyphosate-resistant rice expressing the G2-EPSPS gene[J]. Frontiers in Plant Science,2017,8:885.
[29]TAN S, EVANS R R, DAHMER M L, et al. Imidazolinone-tolerant crops:history,current status and future[J]. Pest Management Science,2005,61(3):246-257.
(责任编辑:石春林)
