
青钱柳二倍体和四倍体叶特征比较研究
2024-09-12 00:00:00刘夏岚宋子琪胡凤荣尚旭岚
南京林业大学学报(自然科学版) 2024年4期



摘要:【目的】比较青钱柳(Cyclocarya paliurus)二倍体和四倍体叶表型特征、解剖结构和气孔特征等方面的差异,旨在获得不同倍性青钱柳叶片特征的基础数据。【方法】以青钱柳二倍体和四倍体苗木为材料,对32项叶片特征指标进行测定,并进行独立样本t检验、相关性分析和主成分分析。【结果】除中部小叶的最大叶宽位置和锯齿宽度在倍性间无显著差异外,其余30项叶特征在倍性间均达到显著或极显著差异。其中,四倍体小叶数量极显著少于二倍体,而复叶面积和比叶质量极显著大于二倍体;四倍体顶端小叶和中部小叶的叶长、叶宽和叶面积极显著大于二倍体,而叶形指数极显著小于二倍体;四倍体锯齿密度极显著小于二倍体;四倍体叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度均极显著大于二倍体,而栅海比和紧密度极显著小于二倍体;四倍体气孔长度和气孔宽度极显著大于二倍体,而气孔密度极显著小于二倍体。相关性分析结果表明,青钱柳表型特征与解剖结构和气孔特征各指标间存在广泛而显著的相关关系。主成分分析提取了反映两种倍性青钱柳叶特征差异的指标,分别为小叶叶宽、小叶面积、叶形指数、海绵组织厚度、气孔长度、栅栏组织厚度和紧密度。【结论】青钱柳二倍体和四倍体的叶特征存在显著差异,小叶叶宽、小叶面积和叶形指数等叶表型特征可用于青钱柳二倍体和四倍体初筛。
关键词:青钱柳;多倍体;表型特征;解剖结构;气孔特征
中图分类号:S722"""""" 文献标志码:A开放科学(资源服务)标识码(OSID):
文章编号:1000-2006(2024)04-0076-09
A comparative study on leaf characters between diploid and tetraploid" of Cyclocarya paliurus
LIU Xialan1,SONG Ziqi1,HU Fengrong2,SHANG Xulan1*
(1. Co-Innovation Center for Sustainable Forestry in Southern China," College of Forestry and Grassland, Nanjing Forestry University, Nanjing 210037, China; 2. College of Landscape Architecture, Nanjing Forestry University, Nanjing 210037, China)
Abstract: 【Objective】 To compare the differences in leaf phenotypes characters, anatomical structures" and stomatal characteristics between diploid and tetraploid Cyclocarya paliurus, we obtained"" data on the leaf characteristics of C. paliurus with different ploidy. 【Method】 Diploid and tetraploid seedlings of C. paliurus were used as study materials, and 32 leaf traits were measured. Independent samples t-tests, a correlation analysis, and principal component analysis (PCA) were conducted. 【Result】 There were no significant differences between diploid and tetraploid seedlings for sawtooth width and the position of the" maximum leaf width of the middle leaflet. For the other 30 leaf traits, there were significant or extremely significant differences between diploid and tetraploid seedlings. Among them, the number of leaflets of tetraploids was significantly less than that of diploids, while the area and specific leaf weight of compound leaves of tetraploids were significantly greater than those of diploids. The length, width" and area of the terminal and middle leaflets of tetraploids were significantly greater than those of diploids, while the leaf shape index was significantly smaller than that of diploids. The sawtooth density of tetraploids was significantly smaller than that of diploids. The leaflet thickness, the" upper epidermis thickness, lower epidermis thickness, palisade tissue thickness, and sponge tissue thickness of tetraploids were all significantly higher than those of diploids, while the ratio of palisade tissue to sponge tissue and cell tense ratio were significantly smaller than those of diploids. The stomatal length and width of tetraploids were significantly greater than those of diploids, while the stomatal density was significantly smaller than that of diploids. The correlation analysis results indicated that there were broad and significant correlations between the phenotypic characteristics, anatomical structure, and stomatal characteristics of C. paliurus. The indicators reflecting the differences of leaf characteristics between diploid and tetraploid C. paliurus were extracted by a PCA, including leaflet width, leaflet area, leaf shape index, sponge tissue thickness, stomatal length, palisade tissue thickness" and cell tense ratio. 【Conclusion】 There were significant differences in leaf traits between diploid and tetraploid C. paliurus, and leaf phenotypic characteristics, such as leaflet width, leaflet area, and leaf shape index, can be used for the preliminary screening of diploid and tetraploid C. paliurus.
Keywords:Cyclocarya paliurus; polyploid; phenotypic characters; anatomical structure; stomatal characteristics
多倍体普遍存在于植物界,多倍化是推动植物进化的重要因素[1]。多倍化常伴随着植物细胞、形态和生理特性的改变[2]。而叶片作为植物进行光合作用和蒸腾作用的重要器官,其形态对于植物的生长发育至关重要[3]。大多数多倍体植株的叶片在横向上常表现为叶大小、叶基部、叶尖形态和叶长宽比等方面异于二倍体;在纵向上,叶片厚度因角质层、表皮层或栅栏组织等组织的细胞大小、层数和细胞间隙的改变也不同于二倍体[4]。
青钱柳(Cyclocarya paliurus)是胡桃科青钱柳属植物,是中国特有的单种属植物[5],集药用、材用和观赏价值于一身[6]。青钱柳叶中多糖类[7]、黄酮类[8]和三萜类[9]等活性物质的含量较高,具有降血糖、降血脂、降血压等多种药用价值[10]。目前对青钱柳的研究多集中在叶生物活性物质的调控[11]、优良品种的选育[12-13]以及无性快繁[14]等方面。南京林业大学青钱柳课题组前期研究发现,青钱柳的自然种群中同时存在二倍体和四倍体两种倍性的植株[15],但有关青钱柳倍性的研究甚少[16-17],鉴于染色体加倍会导致青钱柳叶片性状产生何种变化鲜见报道。本研究以二倍体和四倍体苗木为材料,比较叶表型特征、解剖结构和气孔特征等方面的差异,旨在获得不同倍性青钱柳叶片特征的基础数据,为进一步的研究和开发利用提供参考。
1 材料与方法
1.1 供试材料
试验地位于南京林业大学白马教学科研基地(119°09′E,31°35′N)。2019年11月从湖北省五峰县购买1年生青钱柳实生苗,栽植于无纺布容器中,2020年3月平茬,5月定干,采用染色体计数法确定所有植株的倍性。2020年9月末,选无病虫害的健康二倍体(2n = 2x = 32)和四倍体(2n = 4x = 64)平均株各23株进行相关指标测定。
1.2 性状观测方法
1.2.1 表型特征观测
采集两种倍性所有植株从上往下数第4片完全展开的功能复叶,放入自封袋并用冰盒保鲜带回扫描,60 ℃烘干至质量恒定,测定复叶干质量,比叶质量(specific leaf weight,SLW)为单位叶面积的叶干质量。
采用Adobe Photoshop CC 2019软件对所有复叶的扫描图像(图1)进行指标的记录和测定。
具体指标为小叶数量(number of leaflets,LN)和复叶面积(compound leaf area,CLA),以及顶端小叶和中部小叶(奇数对最中间左侧小叶,偶数对取除去顶端小叶最中间小叶)的叶长(length of leaflets,LL)(叶片基部到顶端的长度)、叶宽(width of leaflets,LW)(叶片最宽处的宽度)、小叶面积(area of leaflets,LA)和叶最宽处至叶尖的距离,叶形指数(leaf shape index,LSI)为叶长与叶宽之比,最大叶宽位置(position of maximum leaf width,MLWP)为叶最宽处至叶尖的距离与叶长之比。按复叶长度将叶片等分为3部分,参考熊中人[18]和郭燕等[19]的方法,测定顶端小叶和中部小叶的叶缘周长和锯齿数量,计算锯齿密度(sawtooth density,SD),另从每片复叶的顶端小叶和中部小叶中部随机选取10个锯齿,测定锯齿宽度(sawtooth width,SWW)、锯齿深度(sawtooth depth,SWD)和锯齿与叶缘间的角度(the angle between sawtooth and leaf margin,LSA),并通过锯齿数量与叶周长之比计算锯齿密度。
1.2.2 解剖结构观测
采集两种倍性所有植株从上往下数第3片完全展开的功能复叶中部的1片小叶,避开叶片主脉切取约5 mm×5 mm的叶块,快速放入70% FAA固定液(体积分数70% 乙醇、冰醋酸、甲醛按照18∶1∶1的体积比混合)中固定保存,采用常规石蜡切片法制片(切片厚度8 μm),番红-固绿对染法染色,每个植株制作3个切片,利用光学显微镜(Olympus" BX41)镜检,DP72 系统采集图像,每个切片随机选取3个图像,采用Adobe Photoshop CC 2019软件测量上表皮厚度(upper epidermis thickness,UE)、栅栏组织厚度(palisade tissue thickness,PT)、海绵组织厚度(spongy tissue thickness,ST)和下表皮厚度(lower epidermis thickness,LE),叶片厚度(leaf thickness,LT)为上表皮厚度、栅栏组织厚度、海绵组织厚度与下表皮厚度的总和,栅海比(ratio of thickness of palisade tissue to sponge tissue,P/S)为栅栏组织厚度与海绵组织厚度之比,紧密度(cell tense ratio,CTR)为栅栏组织厚度与叶片厚度之比,疏松度(sponge ratio,SR)为海绵组织厚度与叶片总厚度之比。
1.2.3 气孔特征观测
上午9:00左右,采集两种倍性所有植株从上往下数第3片完全展开的功能复叶中部的1片小叶,采用指甲油印迹法[20],每个植株制作3个气孔样本,利用光学显微镜(Olympus" BX41,日本)镜检,DP72 系统采集图像,每个样本随机选取3个视野记录气孔数,气孔密度(stomata density,STD)为单位视野面积中的气孔数。另从每个视野随机选取10个气孔,采用Adobe Photoshop CC 2019软件测量气孔长度(stomata length,SL)和气孔宽度(stomata width,SW)。
1.3 数据处理与分析
运用Excel 2019对数据进行统计,计算平均值(mean)、标准差(SD)和变异系数(coefficient of variation,CV)。运用SPSS 26.0对倍性间各性状的显著性进行独立样本t检验,对32项倍性间具有显著差异的性状进行相关性(Pearson)分析和主成分(PCA)分析,采用OriginPro 2021 软件绘制主成分(PCA)排序图和相关性(Pearson)热图。
2 结果与分析
2.1 两种倍性青钱柳叶表型特征的差异
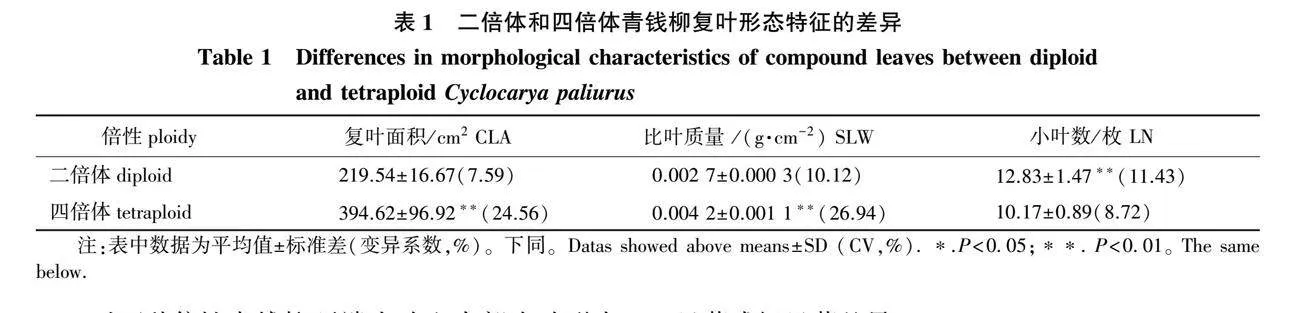
对二倍体和四倍体青钱柳的复叶面积、比叶质量和小叶数量3个指标进行比较可知(表1),倍性间存在极显著差异。其中,四倍体青钱柳的复叶面积和比叶质量极显著大于二倍体(Plt;0.01),四倍体分别为二倍体的1.80倍和1.56倍。小叶数量则表现为四倍体极显著小于二倍体(Plt;0.01),四倍体小叶多为9枚,其次为11枚,少见8或10枚,平均为10.17枚;二倍体小叶多为13枚,其次为11或15枚,少见12或14枚,平均为12.83枚。
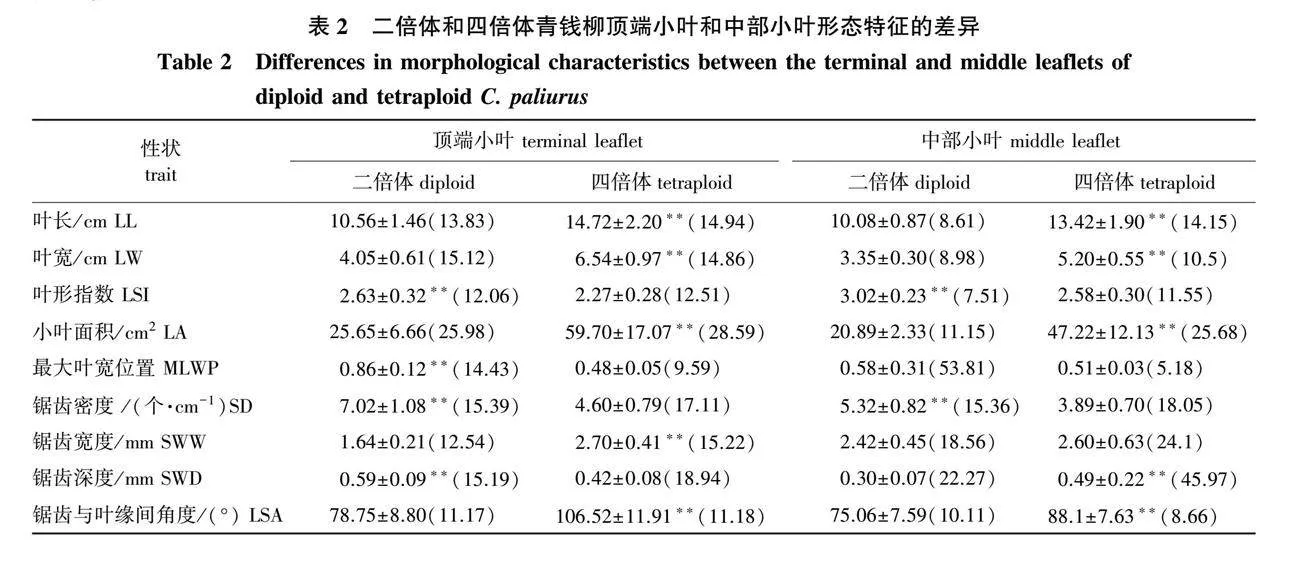
对两种倍性青钱柳顶端小叶和中部小叶形态指标进行比较,结果(表2)可知,顶端小叶各指标在倍性间均存在极显著差异(Plt;0.01);中部小叶除最大叶宽位置和锯齿宽度外,其余各指标均存在显著或极显著差异。
四倍体青钱柳顶端小叶和中部小叶的叶长、叶宽和叶面积极显著大于二倍体(Plt;0.01),而叶形指数和锯齿密度极显著小于二倍体(Plt;0.01)。其中,四倍体顶端小叶的叶长、叶宽和叶面积分别为二倍体的1.39倍、1.61倍和2.33倍;四倍体中部小叶的叶长、叶宽和叶面积分别为二倍体的1.33倍、1.55倍和2.26倍。二倍体顶端小叶和中部小叶的叶形指数分别为2.63和3.02,最大叶宽位置分别为0.86和0.58,说明二倍体青钱柳顶端小叶叶形为倒长椭圆形,中部小叶为长椭圆状披针形。四倍体顶端小叶和中部小叶的叶形指数分别为2.27和2.58,最大叶宽位置分别为0.48和0.51,说明四倍体青钱柳顶端小叶为椭圆形,中部小叶为长椭圆形。
四倍体顶端小叶和中部小叶的锯齿密度分别为4.60和3.89,分别仅为二倍体的65.53%和73.12%。四倍体顶端小叶锯齿宽度和锯齿与叶缘间角度分别为二倍体的1.65和1.35倍,而四倍体顶端小叶的锯齿深度仅是二倍体的71.19%;四倍体中部小叶锯齿宽度、锯齿深度和锯齿与叶缘间角度分别为二倍体的1.07、1.63和1.17倍。由此可以看出,二倍体叶缘锯齿细小且致密,四倍体叶缘锯齿较大且排列较为宽松,而且两个倍性在顶端小叶叶缘锯齿方面的差异大于中部小叶。
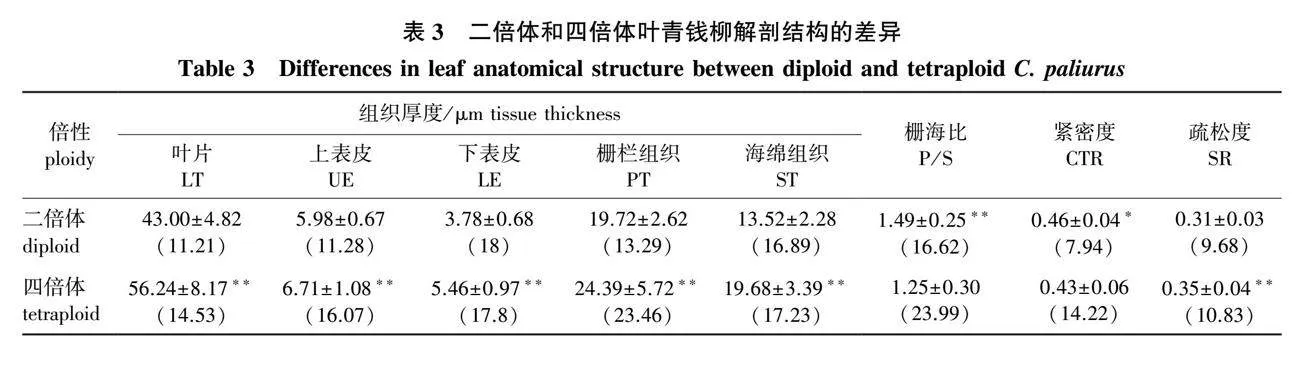
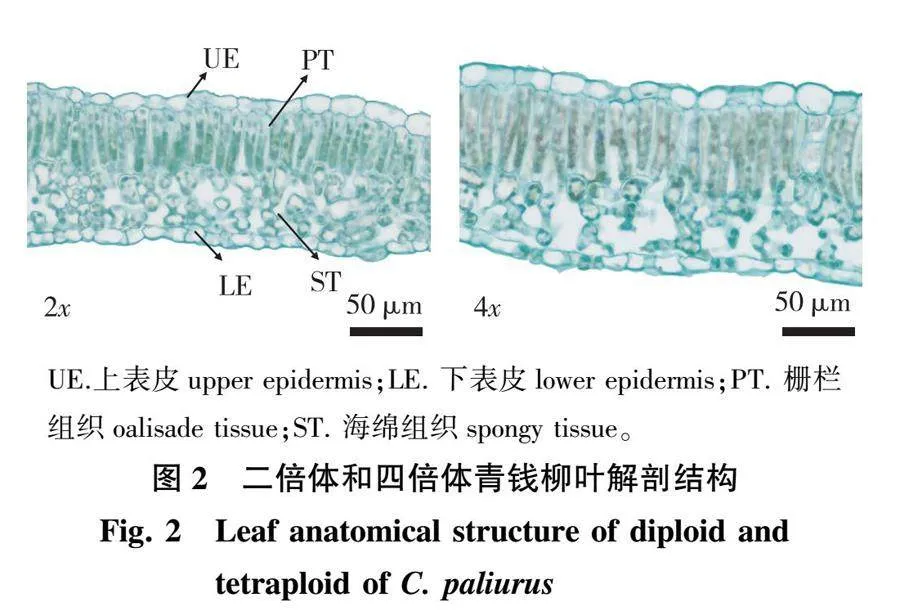
2.2 两种倍性青钱柳叶解剖结构的差异
对青钱柳叶解剖结构进行观测(图2)发现,二倍体和四倍体青钱柳叶均为背腹型叶,二倍体和四倍体的上、下表皮细胞排列均较紧密,形状较规则,叶肉皆分化为栅栏组织和海绵组织。栅栏组织呈长柱形整齐排成1列,而海绵组织排列较疏且不规则。
比较青钱柳二倍体和四倍体叶的解剖结构见表3。
由表3可知,除紧密度呈显著性差异(Plt;0.05),其余指标均为极显著差异(Plt;0.01)。其中四倍体的叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度和疏松度极显著大于二倍体(Plt;0.01),栅海比极显著小于二倍体(Plt;0.01),紧密度显著小于二倍体(Plt;0.05)。具体表现为四倍体的叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度和疏松度分别是二倍体的1.31、1.12、1.44、1.24、1.46和1.12倍,四倍体的栅海比和紧密度是二倍体的83.89%和93.48%。由此可知,四倍体叶片各组织均比二倍体厚,但栅海比和紧密度显著小于二倍体。
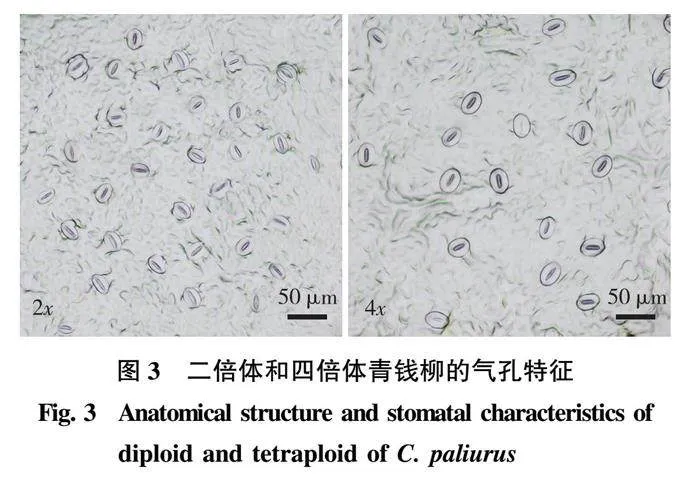
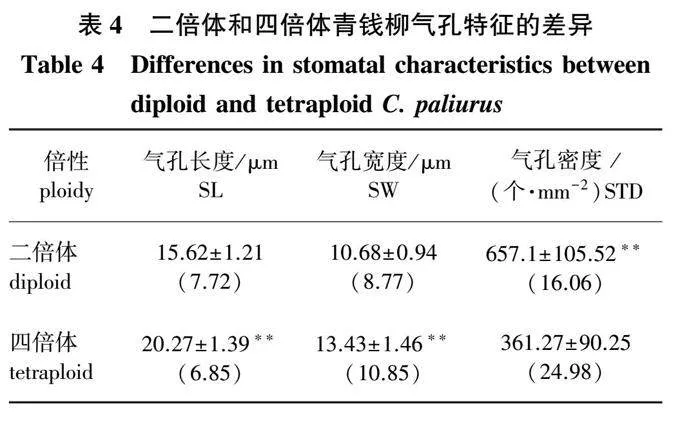
2.3 两种倍性青钱柳气孔特征的差异
对青钱柳二倍体和四倍体的气孔特征进行比较(图3、表4)可知,气孔长度、气孔宽度和气孔密度在两种倍性间均存在极显著差异。其中,四倍体的气孔长度和气孔宽度极显著大于二倍体(Plt;0.01),四倍体的气孔长度和气孔宽度是二倍体的1.30和1.26倍,而气孔密度极显著小于二倍体(Plt;0.01),四倍体的气孔密度仅是二倍体的54.98%。可见,与二倍体相比,四倍体的气孔更大、气孔密度更小。
2.4 两种倍性青钱柳叶特征相关性分析
由青钱柳32项叶特征的相关性分析结果(图4)可知,青钱柳叶片特征的各指标间呈极显著相
关的有327对,其中极显著正相关184对,极显著负相关143对;性状间呈显著相关有76对,其中正相关和负相关各38对。
小叶数量(LN)与大部分性状间具有显著或极显著相关性,其中小叶数量(LN)与复叶面积(CLA)、比叶质量(SLW)、顶端小叶叶长(TLL)、中部小叶叶长(MLL)、顶端小叶叶宽(TLW)、中部小叶叶宽(MLW)、顶端小叶面积(TLA)、中部小叶面积(MLA)、顶端小叶锯齿宽度(TSWW)和中部小叶锯齿深度(MSWD)呈显著或极显著负相关;与顶端小叶最大叶宽位置(TMLWP)、顶端小叶锯齿密度(TSD)、中部小叶锯齿密度(MSD)、顶端小叶叶形指数(TLSI)和中部小叶叶形指数(MLSI)呈显著或极显著正相关。
顶端小叶的叶长(TLL)、叶宽(TLW)和叶面积(TLA)3个叶形指标与中部小叶各叶形指标呈极显著正相关,说明顶端小叶大的,其中部小叶也相应较大。顶端小叶的叶长(TLL)、叶宽(TLW)和叶面积(TLA)3个叶形指标与顶端小叶锯齿密度(TSD)均呈极显著负相关,中部小叶叶形指标与其锯齿密度也呈极显著负相关。
解剖结构和气孔特征与表型特征间大多呈显著相关。其中,叶片厚度(LT)、下表皮厚度(LE)、栅栏组织厚度(PT)、海绵组织厚度(ST)、气孔长度(SL)和气孔宽度(SW)与顶端小叶和中部小叶的叶长、叶宽和叶面积等叶形指标均呈极显著正相关;而紧密度(CTR)和气孔密度(STD)与顶端小叶和中部小叶的叶长、叶宽和叶面积等叶形指标均呈极显著负相关。
以上相关性分析表明,青钱柳叶片形态与解剖结构和气孔特征各指标间存在广泛而显著的相关关系,主要表现为:小叶数量少的,其复叶面积大,顶端小叶和中部小叶的叶长、叶宽和叶面积均较大,而叶形指数小,叶缘锯齿较稀较圆钝,叶片较厚,气孔大但密度小。
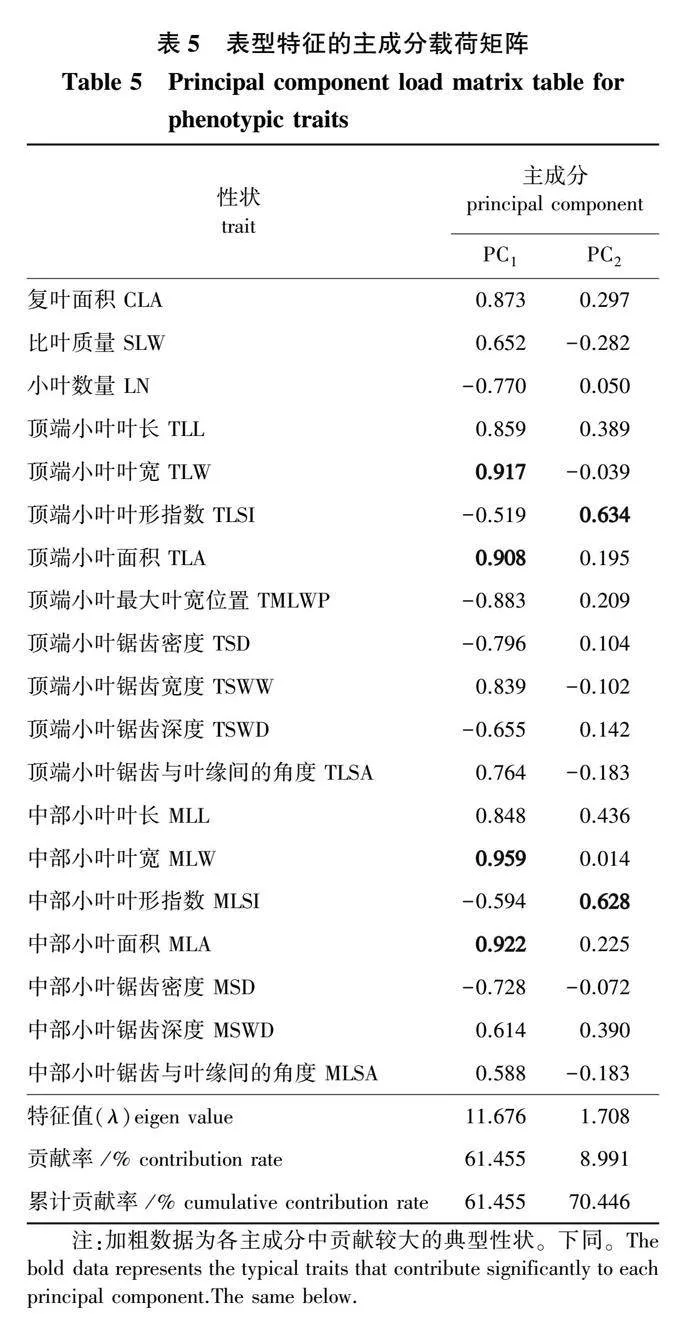
2.5 两种倍性青钱柳叶特征主成分分析
本研究的32项叶特征由于信息大量重叠,进行数据分析较为复杂,借助主成分分析对倍性间差异显著的30项叶特征指标进一步筛选,根据各主成分中每个指标载荷量的绝对值大小筛选出贡献较大的典型指标[21],并对二倍体和四倍体共46个样本进行PCA排序。
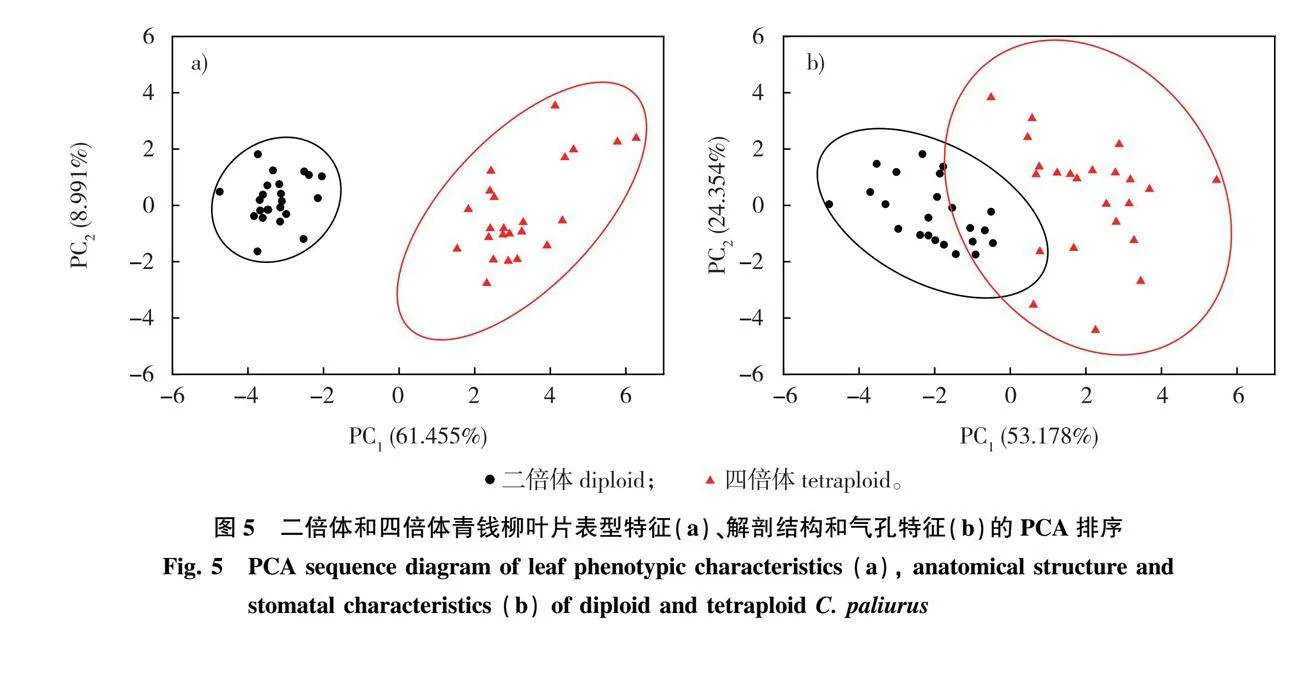
对19项表型特征的指标作主成分分析并绘制PCA排序图,由图5a可知所有样本明显地归在两个独立的置信椭圆内,一类是二倍体,另一类是四倍体,说明根据表型特征可以直接区分青钱柳二倍体和四倍体。由表5可知,前2个主成分的特征值大于1,第1主成分的贡献率为61.455%,第2主成分的贡献率为8.991%,累计贡献率达到70.446%,前2个主成分能较好地概括叶片表型特征的大部分信息。其中第1主成分按贡献值从大到小主要包括中部小叶叶宽(MLW)、中部小叶面积(MLA)、顶端小叶叶宽(TLW)和顶端小叶面积(TLA),第2主成分主要包括顶端小叶叶形指数(TLSI)和中部小叶叶形指数(MLSI),共提取出6个叶表型的典型性状。
对11项解剖结构和气孔特征作主成分分析(表6)发现,前2个主成分的特征值大于1,第1主成分贡献率为53.178%,第2主成分贡献率为24.354%,累计贡献率达到77.532%,说明前2个主成分已经能够代表样本的大部分信息。其中第1主成分按贡献值从大到小主要包括海绵组织厚度(ST)和气孔长度(SL),第2主成分主要包括栅栏组织厚度(PT)和紧密度(CTR),共提取出4个典型性状。从叶片解剖结构和气孔特征的PCA排序图(图5b)发现,二倍体和四倍体的置信椭圆具有部分重叠,说明二、四倍体的叶片解剖结构和气孔特征存在交叉重叠。
3 讨 论
多倍体植物由于多个染色体组的累积,细胞和细胞核的增大,形态上常表现出植株更高、叶片更大更厚、气孔数量增多等特征[22-23],但一些植物在高倍性时也可能表现为发育畸形且生长缓慢[10]。本项研究表明,四倍体青钱柳的复叶面积极显著大于二倍体,而小叶数量则极显著小于二倍体,四倍体青钱柳小叶数量多为9枚,其次为11枚,而二倍体小叶数量多为13枚,其次为11枚。与二倍体相比,四倍体小叶还表现为锯齿较疏松,锯齿较圆钝。唐军荣等[24]的研究也发现,人工诱导的四倍体滇杨(Populus yunnanensis)的叶缘锯齿巨大,二倍体滇杨的叶缘锯齿则细小致密,青钱柳二倍体和四倍体的叶缘锯齿特征与此相似。本研究结果还显示,青钱柳四倍体顶端小叶和中部小叶的叶长、叶宽和叶面积均显著大于二倍体,而叶形指数极显著小于二倍体,这与枣(Ziziphus jujuba)[25]和枇杷(Eriobotrya japonica)[26]多倍体的表现一致。
大量研究表明,多倍体的叶片厚度较二倍体有所增加,如四倍体和三倍体枇杷[26]叶片比二倍体更大更厚;四倍体及嵌合体滇杨的叶片厚度和栅栏组织厚度均大于二倍体[24]。本研究中青钱柳四倍体的叶片厚度和不同组织的厚度都极显著高于二倍体,这与大青杨(P. ussuriensis)[27]和滇杨[24]的研究结果相似。
气孔特征是稳定遗传的性状,在倍性间具有重要的研究价值。气孔大小和气孔密度是鉴别多倍体的典型特征,该特征已普遍应用于橡胶(Hevea brasiliensis)[28]、桂中报春苣苔(Primulina guizhongensis)[29]和黄连木(Pistacia chinensis)[30]等多倍体的鉴别。张源源等[28]对二倍体、三倍体和四倍体橡胶树的研究发现,随着倍性的提高,气孔长度和气孔宽度增大,而气孔密度减小。本研究中,青钱柳二倍体的气孔密度极显著高于四倍体,但气孔长度和宽度极显著低于四倍体,说明青钱柳气孔大小和倍性呈正比,气孔密度和倍性呈反比,这与滇杨[24]、团花(Neolamarckia cadamba)[31]和毛竹(Phyllostachys edulis)[32]多倍体的表现一致。但也有研究表明倍性和气孔没有相关性,郗连连等[33]对花楸属(Sorbus)植物的基因组大小与气孔特征的研究发现,花楸属植物的气孔大小和密度与倍性的相关性不大。
青钱柳表型特征间具有较高程度的相关性,如叶长、叶宽、叶形指数与叶面积间的相关性较高,这与杜鹃叶山茶(Camellia azalea)[34]和荔枝(Litchi chinensis)[35]的研究结果一致。通过主成分分析提取了青钱柳两种倍性叶特征的典型性状,主要包括顶端小叶和中部小叶的叶宽、叶面积和叶形指数,以及海绵组织厚度、气孔长度、栅栏组织厚度和紧密度,叶表型特征的PCA排序图中两种倍性样本归在两个独立的置信椭圆内,说明根据叶表型可直接区分青钱柳二倍体和四倍体。
综上所述,青钱柳二倍体和四倍体的叶片特征存在显著差异,四倍体小叶多为9枚,而二倍体多为13枚;四倍体顶端小叶和中部小叶均较二倍体长,也较二倍体宽;四倍体顶端小叶和中部小叶的叶形指数均较二倍体小,二倍体顶端小叶为倒长椭圆形,中部小叶为长椭圆状披针形,四倍体顶端小叶为椭圆形,中部小叶为长椭圆形;四倍体锯齿较大,较稀疏,二倍体叶缘锯齿较小,较密。在对二、四倍体青钱柳大规模混合群体进行鉴定时,为提高效率,可以通过小叶数量、小叶长度、小叶宽度和叶形指数等表型特征进行初筛,再结合海绵组织厚度、栅栏组织厚度、紧密度和气孔长度等指标进行鉴定,最后采用染色体计数或流式细胞分析法予以验证。由于本研究测定所用叶片均采自盆栽苗,今后还需在不同种源、不同树龄的人工林和天然林植株中开展大量的观测,以进一步验证其可靠性。
参考文献(reference):
[1]WOOD T E,TAKEBAYASHI N,BARKER M S,et al.The frequency of polyploid speciation in vascular plants[J].Proc Natl Acad Sci USA,2009,106(33):13875-13879.DOI: 10.1073/pnas.0811575106.
[2]GUO W,YANG J,SUN X D,et al.Divergence in eco-physiological responses to drought mirrors the distinct distribution of Chamerion angustifolium cytotypes in the Himalaya-Hengduan mountains region[J].Front Plant Sci,2016,7:1329.DOI: 10.3389/fpls.2016.01329.
[3]袁静,周冰莹.植物叶缘锯齿发育调控的研究进展[J].中国农学通报,2021,37(26):24-31.YUAN J,ZHOU B Y.The regulation of leaf margin serration development in plants:a review[J].Chin Agric Sci Bull,2021,37(26):24-31.
[4]晁亚琮,武荣花,蒋卉,等.同源多倍体化对植物侧生器官形态建成影响的研究进展[J].河南农业科学,2021,50(5):1-6.CHAO Y C,WU R H,JIANG H,et al.Research progress on effect of autopolyploid on plant lateral organ morphogenesis[J].J Henan Agric Sci,2021,50(5):1-6.DOI: 10.15933/j.cnki.1004-3268.2021.05.001.
[5]方升佐.青钱柳产业发展历程及资源培育研究进展[J].南京林业大学学报(自然科学版),2022,46(6):115-126.FANG S Z.A review on the development history and the resource silviculture of Cyclocarya paliurus industry[J].J Nanjing For Univ (Nat Sci Ed),2022,46(6):115-126.DOI: 10.12302/j.issn.1000-2006.202206019.
[6]方升佐,尚旭岚,洑香香.青钱柳种子生物学研究[M].北京:中国林业出版社,2017.FANG S Z,SHANG X L,FU X X.Seed biology of Cyclocarya paliurus[M].Beijing:China Forestry Publishing House,2017.
[7]XIA X C,MAO D X,DAI H M,et al.Effect of Cyclocarya paliurus polysaccharides on streptozotocin-induced diabetic nephropathy in rats[J].J Tradit Chin Med,2020,40(6):956-964.DOI: 10.19852/j.cnki.jtcm.2020.06.007.
[8]肖岩,马博稷,李冰涛,等.青钱柳醇提物中化学成分的UHPLC-Q-TOF/MS分析[J].中国实验方剂学杂志,2022,28(16):196-204.XIAO Y,MA B J,LI B T,et al.Analysis of chemical constituents in ethanol extract of Cyclocarya paliurus dried leaves by UHPLC-Q-TOF/MS[J].Chin J Exp Tradit Med Formulae,2022,28(16):196-204.DOI: 10.13422/j.cnki.syfjx.20212051.
[9]WANG Y R,CUI B S,HAN S W,et al.New dammarane triterpenoid saponins from the leaves of Cyclocarya paliurus[J].J Asian Nat Prod Res,2018,20(11):1019-1027.DOI: 10.1080/10286020.2018.1457653.
[10]陈絮蒙,王雅靖,陈靓,等.青钱柳活性物质及其代谢调节作用的研究与应用进展[J].食品与发酵工业,2023,49(12):336-344.CHEN X M,WANG Y J,CHEN L,et al.Research and application progress of active substances and their metabolic regulatory effects of Cyclocarya paliurus[J].Food Ferment Ind,2023,49(12):336-344.DOI: 10.13995/j.cnki.11-1802/ts.035077.
[11]聂小华,吴聪聪,林胜利,等.青钱柳中活性物质及其功能特性研究进展[J].浙江工业大学学报,2022,50(2):222-227.NIE X H,WU C C,LIN S L,et al.Advance on active compounds and functional properties of Cyclocarya paliurus (Batal.) Iljinskaja[J].J Zhejiang Univ Technol,2022,50(2):222-227.DOI: 10.3969/j.issn.1006-4303.2022.02.017.
[12]田力,徐骋炜,尚旭岚,等.青钱柳药用优良单株评价与选择[J].南京林业大学学报(自然科学版),2021,45(1):21-28.TIAN L,XU C W,SHANG X L,et al.Evaluation and selection on superior individuals for medicinal use of Cyclocarya paliurus[J].J Nanjing For Univ (Nat Sci Ed),2021,45(1):21-28.DOI: 10.12302/j.issn.1000-2006.202002018.
[13]徐展宏,朱莹,金慧颖,等.不同叶色青钱柳叶片色素、多酚含量及光合特性的差异[J].南京林业大学学报(自然科学版),2022,46(2):103-110.XU Z H,ZHU Y,JIN H Y,et al.Variations in the contents of leaf pigments and polyphenols and photosynthesis traits in Cyclocarya paliurus with different leaf colors[J].J Nanjing For Univ (Nat Sci Ed),2022,46(2):103-110.DOI: 10.12302/j.issn.1000-2006.202105048.
[14]吴玲利,韩航,曾艳玲,等.青钱柳组织培养及快速繁殖[J].植物生理学报,2019,55(1):61-68.WU L L,HAN H,ZENG Y L,et al.Tissue culture and rapid propagation of Cyclocarya paliurus[J].Plant Physiol J,2019,55(1):61-68.DOI: 10.13592/j.cnki.ppj.2018.0360.
[15]QU Y Q,SHANG X L,ZENG Z Y,et al.Whole-genome duplication reshaped adaptive evolution in a relict plant species,Cyclocarya paliurus[J].Genomics Proteomics Bioinformatics,2023:S1672-S0229(23)00033-5.DOI: 10.1016/j.gpb.2023.02.001.
[16]MAO X,FU X X,HUANG P,et al.Heterodichogamy,pollen viability,and seed set in a population of polyploidy Cyclocarya paliurus (Batal) Iljinskaja (Juglandaceae)[J].Forests,2019,10(4):347.DOI: 10.3390/f10040347.
[17]QU Y Q,WANG Q,YU Y H,et al.Comparative analysis of the complete chloroplast genome between tetraploidy and diploidy of Cyclocarya paliurus (Batal.) Iljinskaja[J].Mitochondrial DNA B Resour,2021,6(9):2669-2671.DOI: 10.1080/23802359.2021.1964402.
[18]熊中人.中国花楸属复叶组植物叶特征及其分类学意义[D].南京:南京林业大学,2019.XIONG Z R.Leaf characters of Sorbus section Sorbus from China and their taxonomic significance[D].Nanjing:Nanjing Forestry University,2019.DOI: 10.27242/d.cnki.gnjlu.2019.000029.
[19]郭燕,张树航,李颖,等.燕山板栗种质资源叶片表型性状多样性研究[J].园艺学报,2022,49(8):1673-1688.GUO Y,ZHANG S H,LI Y,et al.Diversity analysis of leaves phenotypic traits of Yanshan chestnut[J].Acta Hortic Sin,2022,49(8):1673-1688.DOI: 10.16420/j.issn.0513-353x.2021-0760.
[20]曹林青,钟秋平,邹玉玲,等.油桐种质资源叶片结构变异及与环境因子的关系[J].南京林业大学学报(自然科学版),2023,47(4):95-102.CAO L Q,ZHONG Q P,ZOU Y L,et al.Leaf structure variations and relationship with environmental fators among germplasm resources of Vernicia fordii[J].J Nanjing For Univ (Nat Sci Ed),2023,47(4):95-102.DOI: 10.12302/j.issn.1000-2006.202108020.
[21]郭素娟,武燕奇.板栗叶片解剖结构特征及其与抗旱性的关系[J].西北农林科技大学学报(自然科学版),2018,46(9):51-59.GUO S J,WU Y Q.Leaf anatomical structure characteristics and drought resistance of Chinese chestnut[J].J Northwest A F Univ (Nat Sci Ed),2018,46(9):51-59.DOI: 10.13207/j.cnki.jnwafu.2018.09.007.
[22]孙清荣,孙美娟,孙洪雁,等.梨(Pyrus communis L.)多倍体新种质表型变异多样性研究[J].果树学报,2016,33(1):1-7.SUN Q R,SUN M J,SUN H Y,et al.Diversity of phenotypic variation of polyploid plants new germplasm in pear(Pyrus communis L.)[J].J Fruit Sci,2016,33(1):1-7.DOI: 10.13925/j.cnki.gsxb.20150042.
[23]陶抵辉.植物多倍体的研究与应用[J].生物技术通报,2010(7):22-27.TAO D H.Studies and application on plant polyploids[J].Biotechnol Bull,2010(7):22-27.DOI: 10.13560/j.cnki.biotech.bull.1985.2010.07.015.
[24]唐军荣,李斌,朱丽娜,等.滇杨多倍体苗期叶片形态及光合生理比较分析[J].林业科学研究,2016,29(1):103-109.TANG J R,LI B,ZHU L N,et al.Analysis on leaf morphology and photosynthesis physiology of polyploidized Populus yunnanensis seedlings[J].For Res,2016,29(1):103-109.DOI: 10.13275/j.cnki.lykxyj.2016.01.015.
[25]刘金霞,刘盼,米娜瓦尔·亚森,等.南疆地区枣二倍体及其同源四倍体表型性状比较[J].西北农业学报,2022,31(5):595-602.LIU J X,LIU P,Minavar Y S,et al.Comparison of phenotypic characters between diploid and autotetraploid of Chinese jujube in southern Xinjiang[J].Acta Agric Boreali Occidentalis Sin,2022,31(5):595-602.DOI:10.7606/j.issn.1004-1389.2022.05.008.
[26]温国,孙皓浦,党江波,等.多倍体与二倍体枇杷叶片特征及抗旱性初步分析[J].果树学报,2019,36(8):968-979.WEN G,SUN H P,DANG J B,et al.A preliminary study on leaf characteristics and drought resistance of polyploid and diploid loquat[J].J Fruit Sci,2019,36(8):968-979.DOI: 10.13925/j.cnki.gsxb.20190082.
[27]祁传磊,靳春莲,李开隆,等.不同倍性大青杨的光合特性及叶片解剖结构比较[J].植物生理学通讯,2010,46(9):917-922.QI C L,JIN C L,LI K L,et al.Comparison of photosynthetic characteristics and leaf anatomy structure of different ploidy Populus ussuriensis Kom[J].Plant Physiol Commun,2010,46(9):917-922.DOI: 10.13592/j.cnki.ppj.2010.09.022.
[28]张源源,郭涵,袁红章,等.橡胶树不同倍性的气孔性状差异研究[J].热带作物学报,2017,38(3):389-394.ZHANG Y Y,GUO H,YUAN H Z,et al.Stomatal differences among diploid,triploid and tetraploid in rubber tree[J].Chin J Trop Crops,2017,38(3):389-394.DOI: 10.3969/j.issn.1000-2561.2017.03.001.
[29]余文迪,刘娟旭,余义勋,等.桂中报春苣苔四倍体诱导及鉴定[J/OL].分子植物育种,2023:1-12.(2023-02-10).https://kns.cnki.net/kcms/detail/46.1068.S.20230210.1133.004.html.YU W D,LIU J X,YU Y X,et al.Induction and identification of tetraploid in Primulina guizhongensis[J/OL].Mol Plant Breed,2023:1-12.(2023-02-10).https://kns.cnki.net/kcms/detail/46.1068.S.20230210.1133.004.html.
[30]李娟娟,邓伟,许泽楠,等.黄连木多倍体诱导及鉴定[J].东北林业大学学报,2022,50(10):18-22.LI J J,DENG W,XU Z N,et al.Induction and identification of polyploid from Pistacia chinensis[J].J Northeast For Univ,2022,50(10):18-22.DOI: 10.13759/j.cnki.dlxb.2022.10.004.
[31]张石虎,古敏,周玮,等.黄梁木多倍体离体诱导及其性状变异[J].分子植物育种,2022,20(7):2372-2383.ZHANG S H,GU M,ZHOU W,et al.In vitro induction of polyploid and its traits variation of Neolamarckia cadamba[J].Mol Plant Breed,2022,20(7):2372-2383.DOI: 10.13271/j.mpb.020.002372.
[32]徐鹏飞,杨艳红,张毓婷,等.毛竹四倍体诱导及初步鉴定[J].林业科学,2020,56(8):55-62.XU P F,YANG Y H,ZHANG Y T,et al.Induction and preliminary identification of tetraploid in Phyllostachys edulis[J].Sci Silvae Sin,2020,56(8):55-62.DOI: 10.11707/j.1001-7488.20200807.
[33]郗连连,李嘉宝,朱凯琳,等.花楸属3种植物的基因组大小与叶气孔特征分析[J].植物科学学报,2020,38(1):32-38.XI L L,LI J B,ZHU K L,et al.Variation in genome size and stomatal traits among three Sorbus species[J].Plant Sci J,2020,38(1):32-38.DOI: 10.11913/PSJ.2095-0837.2020.10032.
[34]徐斌,彭莉霞,杨会肖,等.杜鹃红山茶叶片主要性状的遗传多样性分析[J].植物研究,2015,35(5):730-734.XU B,PENG L X,YANG H X,et al.Genetic diversity analysis for leaf main traits of Camellia azalea[J].Bull Bot Res,2015,35(5):730-734.DOI: 10.7525/j.issn.1673-5102.2015.05.015.
[35]林祺英,李芳,蔡汝鹏,等.海南荔枝资源叶片性状多样性分析[J].热带生物学报,2023,14(6):628-635.LIN Q Y,LI F,CAI R P,et al.Diversity analysis of leaf traits of litchi genetic resources in Hainan[J].J Trop Biol,2023,14(6):628-635.DOI: 10.15886/j.cnki.rdswxb.20220126.
(责任编辑 吴祝华)
收稿日期Received:2023-05-23""" 修回日期Accepted:2023-06-19
基金项目:国家自然科学基金项目(31971642,32271959);江苏省重点研发计划(现代农业)重点项目(BE2019388)。
第一作者:刘夏岚(68465983@qq.com)。
*通信作者:尚旭岚(shangxulan@njfu.edu.cn),副教授。
引文格式:刘夏岚,宋子琪,胡凤荣,等. 青钱柳二倍体和四倍体叶特征比较研究[J]. 南京林业大学学报(自然科学版),2024,48(4):76-84.
LIU X L,SONG Z Q,HU F R, et al. A comparative study on leaf characters between" diploid" and tetraploid of Cyclocarya paliurus[J]. Journal of Nanjing Forestry University (Natural Sciences Edition),2024,48(4):76-84.
DOI:10.12302/j.issn.1000-2006.202305024.
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09 06:16:12
山西农业科学(2021年3期)2021-12-06 17:36:14
今日农业(2020年14期)2020-12-14 19:47:34
诊断学(理论与实践)(2020年6期)2020-12-09 04:07:52
大众科学(2020年7期)2020-10-26 16:46:40
农业技术与装备(2020年7期)2020-08-18 12:16:00
中成药(2017年6期)2017-06-13 07:30:35
中成药(2017年4期)2017-05-17 06:09:31
临床与实验病理学杂志(2017年3期)2017-03-07 09:33:00
江苏农业科学(2016年9期)2016-11-28 01:35:57
