
山东高优苹果园土壤细菌多样性特征分析
2024-08-24 00:00:00钱琛范伟国张玮玮杨洪强
山东农业大学学报(自然科学版) 2024年1期



关键词: 苹果; 果实品质; 根际土壤菌群; 高通量测序
中图法分类号: S66 文献标识码: A 文章编号: 1000-2324(2024)01-0054-09
山东苹果适宜区逐步由胶东沿海地区向内地转移,形成以沂蒙山区为主的苹果种植中心。
影响苹果产量和品质的因素有很多,施肥、控水、砧木、整形修剪以及地理环境均会对苹果产量造成影响[1],苹果园根际土壤微生物种群数量及活性也是影响果树产量和品质的重要因素之一。植物根系通过分泌一系列次生代谢产物与土壤微生物之间相互作用,从而形成一个共生的根域微生态环境。根际微生物又通过增加繁殖、加快物质循环等方式与植物根系互相作用,进而促进根际土壤有效养分的活化和分配[2],以加快植物根系对养分的吸收,从而达到提高果园产量和改善果实品质的目的[3]。
果园土壤根际微生物是反映土壤健康的指标之一,是评价果园土壤健康的活性因子[4],其在果树所需土壤养分的活化及循环中起着关键作用[5-6]。土壤微生物的种类及多样性作为评价土壤健康与否的生态指标越来越受到重视[7]。因此,有必要进一步深入探索苹果根际土壤微生物群落结构特征,为果园土壤改善提供重要参考。
土壤是一个极其复杂的生态系统,土壤生物化学反应的主要推动力是土壤微生物以及土壤酶[8]。土壤微生物物种多样性的研究越来越受到重视。研究认为不同土壤质地下的微生物数量特征是不同的,微生物活性与土壤黏粒含量成正相关[9-10],一定比例的土壤沙砾可以提高多种土壤酶活性[11]。此外,地理位置也是影响土壤微生物多样性的因素[12]。土壤酶来源广泛,微生物活性与土壤酶活性密切相关[13],如VA真菌可以分泌磷酸酶从而促使土壤有机磷矿质化,促进植物对磷的吸收[14];根际细菌可以促进脲酶、蔗糖酶及磷酸酶活性[15]。土壤微生物和土壤酶与土壤速效养分密切相关[16],土壤培肥可以活化土壤养分,提高土壤酶活性,促进作物增产[17]。因此,探究高优果园中土壤微生物与酶活性特征差异对果树生产具有重要实践意义。
本文采用高通量测序技术对山东苹果主产区高优/低产苹果园土壤微生物特征进行了调查,以期探索高优苹果园根际土壤微生物主要特征,为土壤环境进一步改良提供理论依据。
1 材料与方法
1.1 苹果园基本情况
于2021年分别在山东沂水、蒙阴和曲阜3 地取样调查,采集当地具有代表性的高产果园和低产果园样本。土壤基本情况见表1。
山东沂水县黄山铺镇,天牧果园家庭农场为高产果园,年均1 975-2 172公斤/亩,占地400多亩。黄棕壤,轻黏土质;盆山村为低产果园,年均1717-1803公斤/亩,占地260多亩。果园其它情况均为:株行距:2.5 m×4 m,7年生红富士/平邑甜茶,生草管理,株间开沟施肥,起垄栽植,树形为自由纺锤形。
山东蒙阴县野店镇伊家圈,高产果园年均3634-3998 公斤/亩,占地100多亩;低产果园年均450-500 公斤/亩,占地30多亩。其它基本情况均为:河滩砂土果园,株行距2.6 m×3.5 m;11年生红富士/平邑甜茶,自然生草,株间开沟施肥,起垄栽植,树形为自由纺锤形。
山东曲阜市吴村镇,高产果园年均2085-2294公斤/亩,占地300多亩,全园毛叶苕子生草管理;低产果园年均450-500公斤/亩,占地30 多亩,自然生草。其它情况均为:轻黏土质,株行距为3.5 m×4 m,7年生红富士/平邑甜茶,株间开沟施肥,起垄栽植,树形为小冠疏层形。
1.2 土壤样品采集与测定
1.2.1 土样采集方法 整个苹果园采用5 点法取样,每个点分别选取有代表性的苹果树,在距苹果树主干50 cm 处挖穴取样。首先铲除植被及表面土壤,取15-40 cm深度的苹果根系及根际土壤。根际土壤取样方法采用“抖根法”,抖落根系的大颗粒土壤,用无菌毛刷沿根系表面刷落附着的土壤,视为根际土壤。每个点取混合土壤样量不少于1 kg,装入无菌密封塑料袋中,贴标签,放入自制冰盒中带回实验室测定。带回的土样平均分成两份,-80 ℃冰箱保存土样供高通量测序用,风干土样测定土壤速效养分和土壤酶活性[18-21]等。根系混合样用于根活力测定,根活力采用TTC法[20]测定。
1.2.2 土壤基本指标测定 土壤pH 用pH 显式酸度计测定。土壤有机质用恒沸水浴水合热法测定,碱解氮用碱解扩散法测定,速效磷用NaHCO3 法测定,速效钾用火焰分光光度法测定[18-19]。
1.2.3 土壤微生物数量、土壤酶活测定 用稀释平板培养计数法统计微生物数量[21]。细菌用牛肉膏蛋白胨培养基,真菌用PDA培养基,放线菌用改良高氏1 号培养基。土壤酸性磷酸酶活性测定用磷酸苯二钠比色法[22],蔗糖酶、过氧化氢酶、脲酶活性测定用关松荫[23]的方法。
1.2.4 土壤细菌群落结构多样性测定 由上海派森诺生物科技有限公司进行细菌的高通量测序。土壤总DNA提取,用E.Z.N.A.®土壤DNA试剂盒提取。并用琼脂糖凝胶电泳检测提取DNA 的纯度和浓度。选择细菌V3-V4 区的16SrRNA 序列进行PCR 扩增,扩增引物为338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)。
1.3 苹果产量与品质调查与测定
随机选取9株生长结果正常的苹果树,统计每株树苹果总个数以及每亩果树栽植数,每株树分4 个方位采摘果实不少于12 个,调查单果重和果实品质。
单果质量采用1/100电子天平进行称重(g);果实纵、横径采用游标卡尺进行测量,果形指数用果实纵径和横径比值表示;数显式糖度计(HT4-TD-45 型)测量可溶性固形物含量;酸度用酸度计(GMK-835F 型)测定;果实硬度用硬度计(GY-4-J 型)测定。
1.4 数据处理与分析
物种丰度图、PcoA主成分分析图和冗余分析图等均通过在线平台(https://www.genescloud.cn/home)进行计算和绘制;Chao1、Shannon 和Pielou-e 指数用SPSS22.0 进行统计分析。其余数据用SPSS2.0 进行随机单因素方差分析,Duncan 氏新复极差法(SSR 法)进行多重比较,不同小写字母表示处理间差异达0.05显著水平。
2 结果与分析
2.1 高优苹果园果实品质分析
三地高产果园的单果质量(除沂水果园外)、可溶性固形物含量及糖酸比均明显高于其低产果园,三地高产果园果形指数等其它指标与其低产果园无显著差异(表2)。三地高产果园果实综合品质和产量均高于其低产果园。
三地高产果园中,曲阜果园单果质量最大,其次为沂水及蒙阴果园,其中曲阜果园单果质量与蒙阴果园相比增加了30.99%;曲阜和沂水果园可溶性固形物含量和糖酸比均显著大于蒙阴果园,其中曲阜果园可溶性固形物含量与蒙阴果园相比增加了18.36%;蒙阴果园果形指数显著大于沂水和曲阜果园。表明,曲阜果园果实综合品质最高,其次为沂水果园和蒙阴果园。
2.2 高优苹果园土壤微生物数量与活性分析
2.2.1 高优苹果园根际土壤微生物数量差异 土壤可培养菌数量结果表明,三地高产果园的细菌数量均明显高于其低产果园,真菌数量(除蒙阴果园外)和放线菌数量则明显低于其低产果园(表3),其中沂水和曲阜高产果园的细菌数量分别是其低产果园的2.25和2.80倍,而蒙阴高产果园真菌数量明显高于其低产果园。
三地高产果园中,沂水和曲阜果园土壤细菌数量差异不显著,与蒙阴果园差异显著,其中沂水果园土壤细菌数量与蒙阴果园相比增加了90.99%;蒙阴果园土壤真菌数量显著高于沂水和曲阜果园,其中蒙阴果园土壤真菌数量与曲阜果园相比增加了82.47%,沂水和曲阜果园土壤真菌数量差异不显著。曲阜果园土壤放线菌数量显著高于沂水和蒙阴果园,曲阜果园土壤放线菌数量与蒙阴果园相比增加了122.17%,这可能与曲阜果园多年全园生草有关。沂水和曲阜果园的细菌与真菌比值显著高于蒙阴果园(表3)。表明,蒙阴河滩沙地菌类数量与曲阜、沂水轻壤土质有明显不同,曲阜、沂水相同土质果园在不同土壤管理模式下土壤菌类构成也有差异。
2.2.2 高优苹果园根际土壤酶活性 土壤酶活性结果表明,三地高产果园的脲酶和蔗糖酶活性明显高于其低产果园(表4)。其中蒙阴和曲阜高产果园脲酶活性分别是其低产果园的1.5和7.41倍。沂水、蒙阴和曲阜三地高产果园蔗糖酶活性分别是其低产果园的1.15、1.37和1.31倍。蒙阴高产果园酸性磷酸酶活性是其低产果园的1.87 倍。曲阜高产果园过氧化氢酶活性是其低产果园的1.56 倍,而沂水高产果园过氧化氢酶活性则低于其低产果园。由表4 还看出,三地高产果园中,不同土壤酶活性变化差异较大。土壤脲酶和土壤蔗糖酶可以作为高产果园的特征酶,不同土质和土壤管理模式对土壤酶活性有较大影响,其中全园生草的曲阜果园脲酶活性最高,轻壤土质的果园蔗糖酶活性明显高于沙壤土质果园。
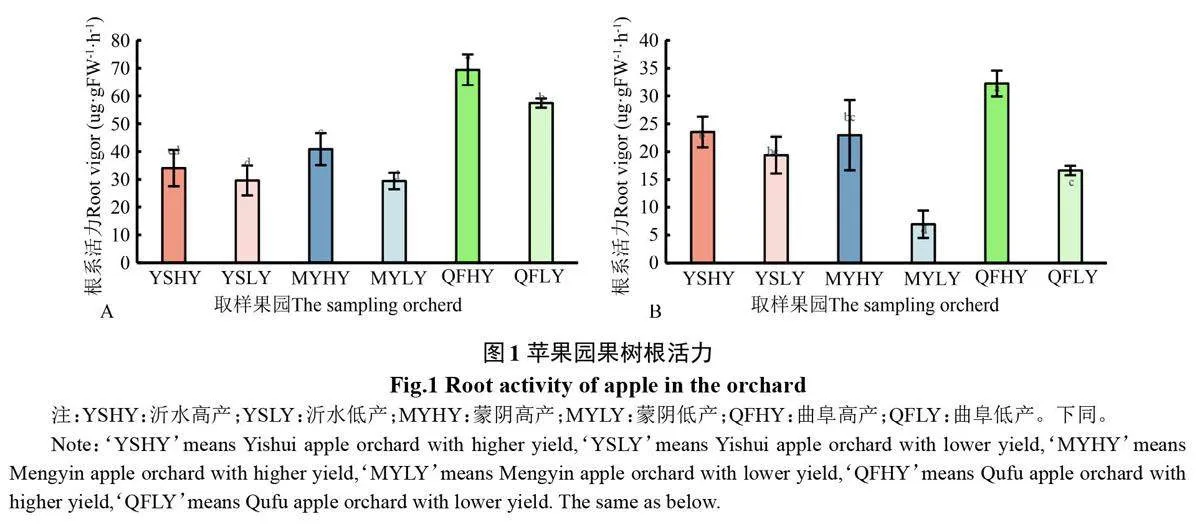
2.2.3 高优苹果园根活力 图1看出,三地高优果园苹果根活力均高于其低产果园。春季(图1-A)沂水、蒙阴和曲阜高产果园苹果根活力分别是其低产果园的1.15、1.39 和1.21 倍。秋季(图1-B)沂水、蒙阴和曲阜高产果园苹果根活力分别是其低产果园的1.22、3.30 和1.94 倍。三地高产果园中,春季和秋季曲阜高产果园苹果根活力均显著高于沂水和蒙阴高产果园,沂水和蒙阴高产果园苹果根活力差异不显著(图1-A、图1-B)。结果表明,根活力也可以作为高产优质果园的一个参考指标,其可能受到土壤综合管理模式的影响较大。
2.3 高优苹果园土壤细菌多样性分析
2.3.1 高优苹果园根际土壤细菌测序多样性差异 三地6 个果园根际土壤细菌高通量测序共获得9 735 799 个高质量细菌序列。序列长度集中在404 bp-431 bp 之间,抽平后生成47 178 个ASVs。六个果园的稀释曲线基本趋于平缓,其覆盖率(Good's coverage)在93.26%-99.07% 之间,说明取样基本合理,细菌群落结构的置信度较高,能够反映取样果园土壤细菌群落的特点。
三地高产果园的ASVs 数量由大到小依次为曲阜、沂水和蒙阴(分别为10 751、9 975 和6 821 个),三地高产果园公有ASVs 数量562 个;三地低产果园的ASVs 数量由大到小依次为沂水、蒙阴和曲阜(分别为9052、8556和7062个),低产果园公有ASVs 数量260 个;曲阜和沂水两地高产果园ASVs 数量显著高于其低产果园。表明,曲阜和沂水高产果园的土壤细菌种群比蒙阴果园更加丰富,可能与曲阜和沂水果园持续多年生草有关,稳定和改善了其土壤生态环境,更有利微生物群落的形成和繁殖。
2.3.2 高优苹果根际土壤细菌分布情况及物种组成分析 三地6 个果园根际土壤细菌门水平的相对丰度如图1-A所示,根际土壤细菌为11 个门,占比前四的优势菌门依次是放线菌门(Actinobacteria)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)和酸杆菌门(Acidobacteria),四者占总细菌数的80%以上,其中放线菌门所占比例(43.06%-68.10%)最高,这四个优势菌门在不同的果园土壤细菌门水平分类组成上所占的比例存在一定差异。三地高产果园的放线菌门(曲阜除外)、绿弯菌门的相对丰度低于其低产果园;三地高产果园的变形菌门、酸杆菌门(蒙阴除外)的相对丰度高于其低产果园。三地高产果园间的放线菌门相对丰度呈现出:蒙阴高产gt;沂水高产gt;曲阜高产;三地高产果园间的变形菌门相对丰度呈现出:沂水高产gt;曲阜高产gt;蒙阴高产。三地高产果园间的酸杆菌门相对丰度呈现出:沂水高产gt;曲阜高产gt;蒙阴高产。
三地6 个果园根际土壤细菌纲水平的相对丰度如图1-B 所示,根际土壤细菌共分为11 个纲,占比前四的优势菌纲依次是放线菌纲(Actinobacteria)、α-变形菌纲(Alphaproteobacteria)、γ-变形菌纲(Gammaproteobacteria)和嗜热菌纲(Thermoleophilia),四者占总细菌数的68% 以上,其中放线菌纲所占比例(33.84%-63.40%)最高,这四个优势菌纲在不同果园土壤细菌分类组成上所占的比例存在一定差异。三地高产果园的放线菌纲(曲阜除外)的相对丰度低于其低产果园;三地高产果园的α-变形菌纲(蒙阴除外)、γ-变形菌纲(曲阜除外)、嗜热菌纲(沂水除外)的相对丰度高于其低产果园。三地高产果园间的放线菌纲相对丰度呈现出:蒙阴高产gt;沂水高产gt;曲阜高产;三地高产果园间的α-变形菌纲相对丰度呈现出:曲阜高产gt;沂水高产gt;蒙阴高产。
三地果园的门、纲水平相对丰度在一定程度上反映了三地果园土壤微生物的种群特征,无论是门水平还是纲水平相对丰度,放线菌门、纲和变形菌门、纲都是三地果园土壤细菌的首要菌群。
2.3.3 高优苹果园根际土壤细菌Alpha 与Beta 多样性差异 由表5 看出,曲阜和沂水两地高产果园土壤细菌的Chao1 指数均分别高于其低产果园,曲阜和沂水高产果园土壤细菌的Shannon指数和Pielou-e 指数显著高于蒙阴果园。三地高产果园土壤细菌的Alpha 多样性指数在一定程度上反应了果园微生物数量及丰富度,结果表明,曲阜和沂水高产果园的土壤细菌多样性比蒙阴果园更加丰富,曲阜和沂水两地果园均为轻壤土质且果园生草情况明显优于蒙阴河滩沙壤土质。这说明,良好的果园土壤及生草覆盖稳定和改善了果园土壤的生态环境,更有利微生物群落的形成和繁殖。
三地6个果园根际土壤细菌的Beta多样性指数显示,任一果园的3个重复样本的聚集性较好,显示出较好的重复性,不同地区果园土壤细菌群落差异显著(P=0.001,R=1)。三地高产果园细菌样本群落内的组成差异均低于其低产果园,三地高产果园间细菌样本群落内的组成差异不显著。说明,高产果园细菌群落结构更为稳定。
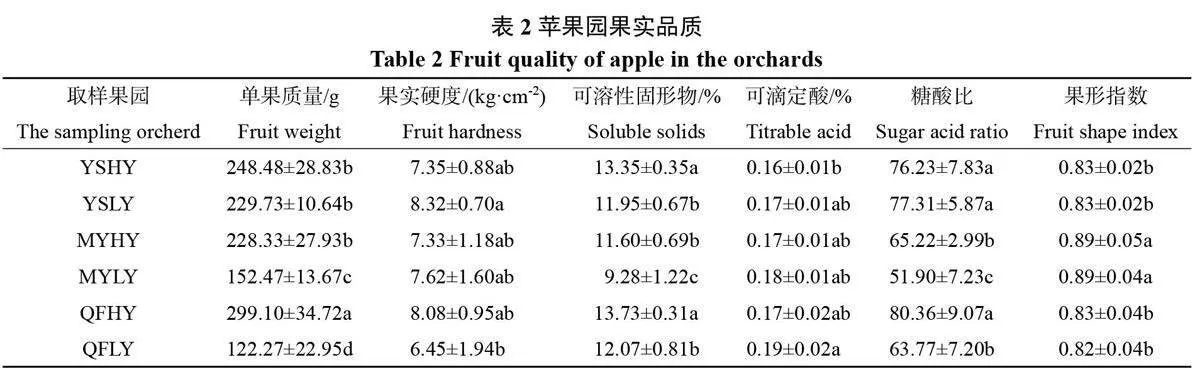
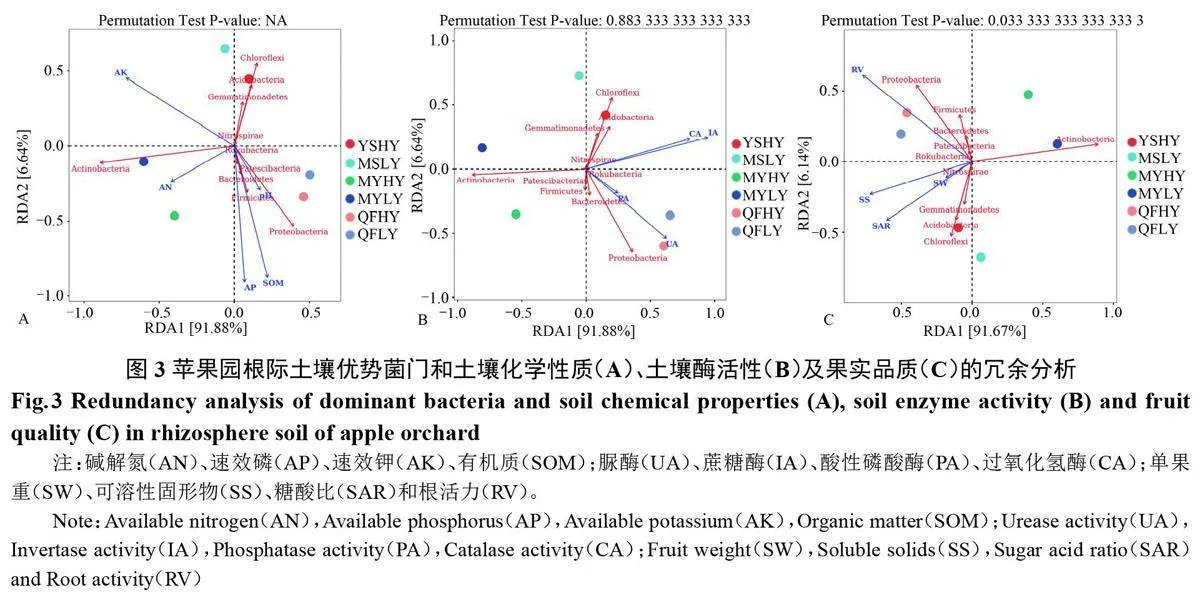
2.4 高优苹果园土壤细菌组成与土壤化学性质、酶活性及果实品质的冗余分析
三地6个果园根际土壤细菌群落平均丰度前10位的门水平的丰度数据与土壤pH和养分含量(图3-A)、土壤酶(图3-B)和果实品质(图3-C)的RDA 冗余分析结果(图3)显示,三地高产果园中,沂水高产果园根际土壤细菌群落相对丰度与其土壤速效钾含量、过氧化氢酶活性、蔗糖酶活性、糖酸比、可溶性固形物含量呈正相关;蒙阴高产果园根际土壤细菌群落相对丰度与其土壤pH和各养分含量呈正相关,与土壤酶活性及果实品质指标呈负相关;曲阜高产果园根际土壤细菌群落相对丰度与土壤pH、有机质及速效磷及脲酶活性、根活力等呈正相关性。蒙阴高产果园还应继续进行土壤微环境改良,以优化微生物群落结构,提升脲酶、蔗糖酶等土壤酶活性,进而利于果实品质的提高;曲阜和沂水高产果园良好的土壤微环境有利于有益微生物群落的生长发育。
3讨论
根际微生物数量通常在数千到数百万之间波动,微生物作为土壤生物中最活跃的存在,参与众多的生物化学反应[24]。同时土壤微生物也不是单独存在的,根际微生物种群类型、数量及活性是随环境改变动态变化的,在特定条件下,土壤微生物与宿主植物形成复杂的缔合网络,维系宿主-微生物间的稳态[25]。高优果园土壤中菌落种群和构成更为优化稳定,细菌种群更加丰富,土壤酶活性增高,使得土壤有效养分供给稳定和充足。如高优果园根际土壤变形菌门中的α-变形菌纲、γ-变形菌纲等丰度明显提高,这些菌纲包含了有利于氮素固氮的固氮细菌[33]、假单胞菌等有益菌属,有利于氮素的吸收同化。土壤微生物数量与土壤酶活性密切相关,细菌对脲酶和蔗糖酶活性有显著的促进作用[25],高优苹果园根际土壤脲酶、蔗糖酶等活性增强,这些均利于维持土壤有效养分的稳定供应。土壤环境的改善也利于根吸收养分能力增强,从而促进果实品质和产量提高。连续多年的苹果种植,根系分泌物的数量增多促进低产果园土壤真菌数量的增加[26],土壤放线菌在代谢过程中所产生的抗菌素如链霉素、尼达霉素、武夷菌素等可以有效抑制细菌、真菌、病毒等的生长,低产果园放线菌数量更高可能是应对真菌、病毒、线虫较多生态的自我保护方式[27]。高产果园土壤管理更精细,其根际土壤的细菌、真菌和放线菌占比比低产果园更合理,微生物群落结构也更加稳定[28]。
影响果园根际土壤微生物生命活动的因素很多,如植物种类、生命生长发育阶段、年周期变化及植株长势、土壤类型和土壤管理方式等[29],根际土壤微生物与土壤质地密切相关[30]。不同土质对微生物种群和多样性有很大影响,曲阜和沂水轻黏壤土与蒙阴河滩沙地果园对其土壤中菌类种群数量和构成有明显影响,相同土质不同土壤管理模式也对其土壤菌类构成有一定影响。研究认为,微生物数量和活性与土壤黏粒的含量成正相关[15-16],黏土和壤土中根际细菌数量明显高于砂土中的[31-32]。高产苹果园根际土壤的脲酶、蔗糖酶活性更大,细菌对脲酶和蔗糖酶活性有显著的促进作用[34],这维系了土壤有效养分的稳定供应,促进碳、氮素等的分解释放,土壤环境得以改善,根系吸收同化能力增强,果实品质和产量也得到提高。值得注意的是,蒙阴果园土壤酸化较重,土壤pH 通过改变土壤营养元素的形态而影响微生物群落[35],土壤酸化还会降低绿弯菌门和酸杆菌门的相对丰度[36]。因此沙土果园土壤管理中还要注意土壤酸碱度的变化,合理施肥和管理,维持适宜酸碱平衡,保证根际土壤微生物种群的有益发展。根际土壤微生物与植物生长环境密切相关[37],对低产果园的改造中,果园间作绿肥、合理生草和覆盖均能提高养分分解和循环速率,促进促生菌和共生微生物生长,并抑制部分有害真菌的繁殖[38],此外配施生物菌肥也能明显改变土壤细菌相对丰度[39]等,生产者可根据果园实际情况选择适合方式促进生产。
本文从土壤细菌群落多样性角度进行了分析,以此探究其在苹果园根际土壤中的作用以及对苹果产量品质的影响。由于土壤微生物包括细菌和真菌等很多种群,因此有待对土壤微生物其它群落多样性的影响做进一步研究。
4结论
高优果园土壤中菌落种群和构成更为优化稳定,细菌种群更加丰富,土壤酶活性增高,促进根吸收养分能力增强,果实品质和产量提高,但不同土质和管理水平对苹果果实品质也有较大影响。
猜你喜欢
中国中药杂志(2016年24期)2017-04-18 17:42:52
中国中药杂志(2017年4期)2017-03-28 17:06:08
中国中药杂志(2017年3期)2017-03-20 21:11:11
科技创新导报(2016年28期)2017-03-14 11:32:29
作文与考试·小学高年级版(2016年17期)2016-12-23 20:21:17
农家科技下旬刊(2016年9期)2016-12-15 10:37:39
农业与技术(2016年20期)2016-12-08 21:48:31
小朋友·快乐手工(2016年10期)2016-12-08 06:15:06
中国科技博览(2016年22期)2016-11-01 14:13:10
现代园艺(2016年17期)2016-10-17 06:35:23
