
激素对圆齿野鸦椿种子萌发的影响
2024-08-22 00:00:00阳先波冉芳周宇曦涂淑萍蔡军火姜雪茹钟鑫宏
南方农业学报 2024年1期
摘要:【目的】探究激素在圆齿野鸦椿种子萌发中的作用及其与活性氧的关系,为圆齿野鸦椿种子破眠技术和种苗高效繁育技术提供理论依据。【方法】以自然层积210d的圆齿野鸦椿种子为试验材料,去除外部硬质种壳,进行为期12d的萌发试验,测定纯水、不同浓度的赤霉素(GA₃)(20、40和60μmol/L)和脱落酸(ABA)(10、20和30μmol/L)处理下萌发率及活性氧、抗氧化物质、植物激素、可溶性糖和可溶性蛋白含量的变化,并观察记录种子的萌发形态。【结果】纯水处理萌发率最终达77.8%,GA,处理萌发率最高可达82.2%,ABA处理萌发率最终均未达50.0%,而保留硬质种皮纯水处理的种子萌发率仅17.8%。3种浓度的GA,处理均可有效缩短圆齿野鸦椿种子萌发率达50.0%的时间(T。),促进根茎生长,但其最终萌发率均与纯水处理无显著差异(Pgt;0.05);ABA处理浓度越高,对圆齿野鸦椿种子抑制萌发作用越强。相关分析显示,在萌发的第8和12d,萌发率与GA,、油菜素内酯(BR)和茉莉酸(JA)呈显著正相关(Plt;0.05,下同),且超氧阴离子(O2-)与BR和JA含量呈显著负相关,与过氧化氢酶(CAT)、ABA、ABA/GA₂均呈显著正相关。【结论】添加GA₃可有效缩短圆齿野鸦椿种子萌发率达50%的时间,促进根茎生长,O2-与ABA的积累对种子萌发存在抑制作用,BR和JA对种子萌发的促进作用可能与对O2-的清除有关。
关键词:圆齿野鸦椿;种子萌发;植物激素;活性氧
中图分类号:S792.99文献标志码:A文章编号:2095-1191(2024)01-0160-09
Effects of hormones on seed germination of Euscaphis konishii
YANG Xian-bo,RAN Fang,ZHOU Yu-xi,TU Shu-ping,CAI Jun-huo,JIANG Xue-ru*,ZHONG Xin-hong
(College of Landscape and Art,JiangxiAgricultural University,Nanchang,Jiangxi330045,China)
Abstract:[Objective]The role of hormones in seed germination of Euscaphis konishi and its relationship with reac-tive oxygen species were explored to provide an important theoretical basis for its seed dormancy breaking technology and efficient seedling breeding technology.【Method]E.konishii seeds with210d natural stratified were used as materials,outer hardseed coat was removed,a12d germination experiment was conducted.The changes of germination rate,active oxygen,antioxidant substances,plant hormones,soluble sugar andsoluble protein contens underpure water,different con-centrations of gibberellin(GA₃)(20,40and60μmolL)and abscisic acid(ABA)(10,20and30μmol/L)were measured and germination morphology ofseeds was observed.【Result]The germination rate of pure water treatment was77.8%,the germination rate of GA₃treatment was up to82.2%,the germination rate of ABAtreatment did not reach50.0%,and the germination rate of pure water treatment with hard seed coat was only17.8%.The three concentrations of GA;treat-ments could effectively shorten the time when germination rate reached50.0%(Tso)and promote root and stem growth,but the final germination rate was not significantly different from that of pure water treatment(Pgt;0.05).ABA treatment could effectively inhibit the germination of E.konishii seeds.The higher the concentration ofABA,the stronger the inhibi-tion.Correlation analysis showed that the germination rate was significantly positively correlated with GA,brassinolide(BR)and jasmonic(JA)at8and12d(Plt;0.05,the same below),superoxide anion(Oz·)was negatively correlated with BR and JA contents,and significantly positively correlated with catalase(CAT),ABA,ABA/GA₃.【Conclusion]Adding GA₃can effectively shorten the time of the seed germination rate of E.konishii to reach50%and promote the growth of root and stem.The accumulation of Oz·and ABA inhibit seed germination and the promotion of BR andJA on seed germi-nation in E.konishiimaybe related to the removal ofO₂
Keywords:Euscaphiskonishii;seed germination;phytohormone;reactive oxygen
Foundation items:National Natural Science Foundation of China(31960327);Jiangxi Natural Science Foundation(20212BAB215014)
0引言
【研究意义】圆齿野鸦椿(Euscaphis konishii)为省沽油科(Staphyleaccae)野鸦椿属(Euscaphis)常绿小乔木,是我国南方特有的药赏两用珍贵本土树种,其果实成熟裂开后如红色蝴蝶挂满枝头,红果期长,跨越秋、冬、春三季。此外,圆齿野鸦椿还富含黄酮类、三萜类、酚酸类和倍半萜类等多种生理活性物质,具有抗炎、抗肿瘤、抗肝纤维化及抑菌等功效(梁文贤等,2018)。圆齿野鸦椿种子具有深休眠特性(游双红,2013),发芽率低、出苗不整齐是制约其苗木培育和规模化生产的主要原因。有报道表明脱落酸(ABA)和赤霉素类(GAs)可通过影响种子的活性氧(ROS)与植物激素间的相互关系以调控种子萌发(李敏和何勇,2023),但其在圆齿野鸦椿种子萌发过程中的相互作用尚不清楚。因此,揭示ABA和GAs诱导圆齿野鸦椿种子萌发中的生理机制,对探究激素在圆齿野鸦椿种子休眠的调控作用及圆齿野鸦椿种苗高效繁育具有重要意义。【前人研究进展】在大部分物种中,环境信号对种子休眠的影响主要是由ABA和GAs介导(Finch-Savage and Leubner-Metzger,2006)。根据激素信号模型,ABA(发芽抑制作用)和GAs(发芽促进作用)的相对效果是休眠水平和种子发芽的主要决定要素。例如,在云南红豆杉(Taxus yummanensis)种子湿沙层积期间,种胚继续生长,赤霉素(GA₃)和玉米素核苷(ZR)含量相对稳定,ABA含量下降(Bian et al.,2018)。银杏(Ginkgo biloba)种子休眠受ABA/GA3平衡的调控,GAs的生物合成和信号转导对休眠解除起着重要作用(Jia et al.
2021)。研究发现除ABA和GAs外,ROS作为信号分子,在植物种子休眠打破和萌发过程中也具有重要作用。此外,ROS还可能与激素信号相互作用,共同参与种子休眠和萌发的环境条件感知和传导(李敏和何勇,2023)。低温层积下的黑孜然(Bunium persicum)种子深层复杂生理休眠的释放与ROS和ABA/GA₃相互作用有关(Amooaghaie and Ahmadi,2017)。在大麦的研究中发现,ROS可通过ABA8°OH1基因影响ABA水平进而影响种子休眠和萌发(Ishibashi et al.,2017)。可见,ABA和GAs及ROS在种子休眠及萌发调控机制中具有重要作用。【本研究切入点】目前,ABA、GAs及ROS关于调控和影响圆齿野鸦椿种子休眠和萌发的机制尚不清楚。【拟解决的关键问题]以层积210d的圆齿野鸦椿种子为试验材料,测定外源添加ABA和GA₃对其萌发、ROS和植物激素的影响;比较ABA和GA₃在圆齿野鸦椿种子萌发过程中的作用,并将萌发率与各项生理指标进行相关分析,探究激素与ROS在圆齿野鸦椿种子萌发中的作用,以期为缩短圆齿野鸦椿育苗周期和促进产业市场化发展提供理论基础。
1材料与方法
1.1试验材料
供试种子来源于江西农业大学花卉基地(28°4534\"N,115°4948\"E)自然生长的20年树龄的圆齿野鸦椿母树。采集时间为2021年12月初,挑选颗粒饱满、无病虫害的种子于常温水浸泡2d后装入细孔尼龙袋中,置于树荫处自然沙藏层积。
1.2种子萌发试验与萌发形态观察
于2022年6月初取出层积时间为210d的种子进行萌发试验。种子经消毒和润洗后,采用50℃水浸种(阳先波等,2022)。经过上述预处理后,去除坚硬外种皮(如无特殊说明,以下提到的种子均为去除坚硬外种皮的种子),外源施加不同浓度的ABA(10、20和30μmol/L)和GA₃(20、40和60μmol/L)处理,以纯水处理的种子为对照1(CK1),以保留硬质种皮的种子为对照2(CK2),共8个处理,每个处理重复3次,每个重复30粒种子。将种子置于垫有1层滤纸、直径为9cm的培养皿中,在人工气候箱内发芽,由于脱落酸水溶液对光敏感,属强光分解化合物,因此整个萌发试验在黑暗环境中进行,温度为25℃,萌发时间为12d,每2d更换1次滤纸,每次加2mL的处理溶液。根据萌发结果,分别取去壳种子中CK1、20μmol/LABA及40μmol/L GA,处理下萌发时间第0、4、8和12d的圆齿野鸦椿种子,经纯水润洗、吸水纸擦干后,液氮冷冻并储存于-80℃超低温冰箱备用。
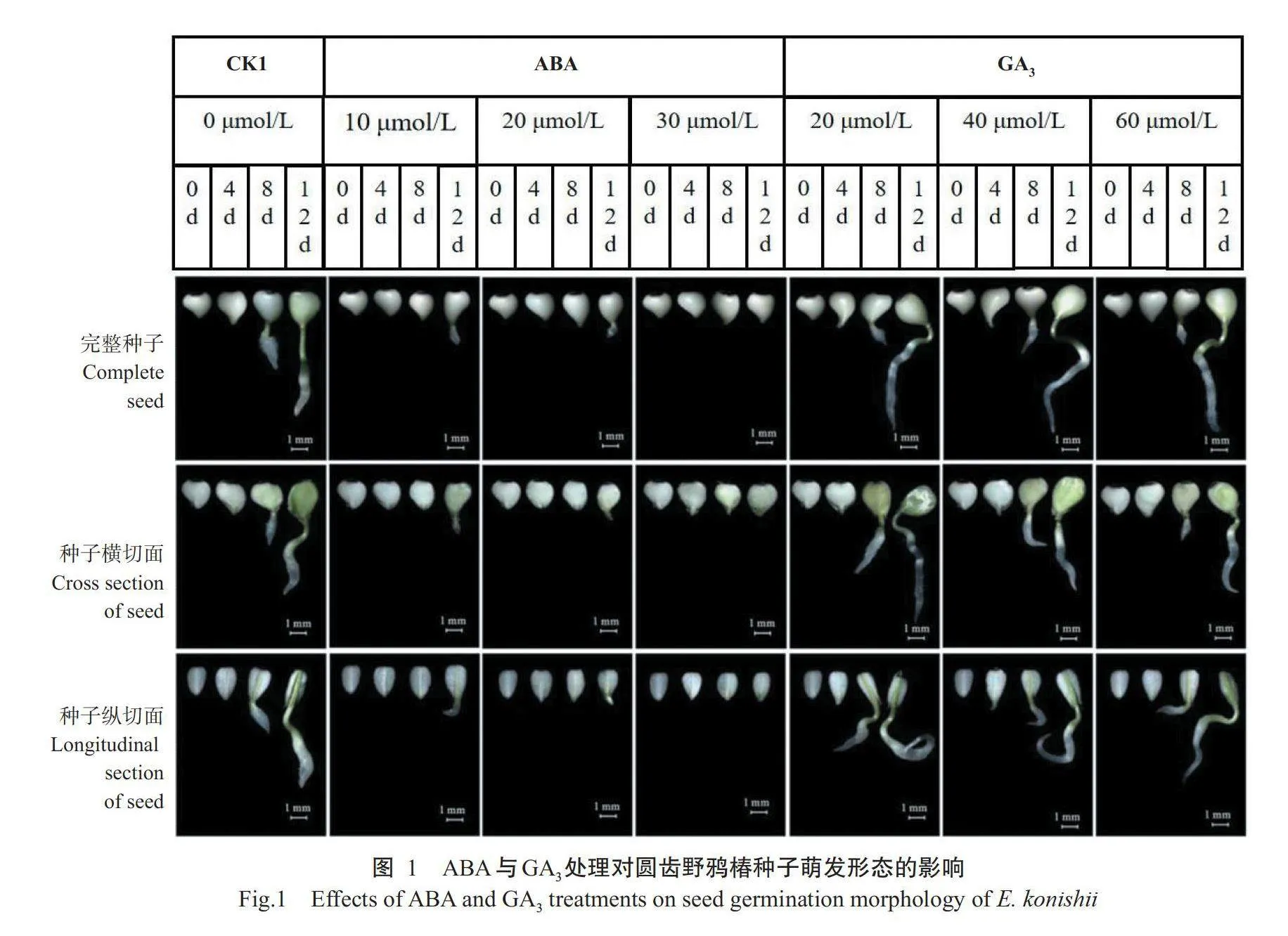
前期预试验表明圆齿野鸦椿去壳种子萌发12d后其萌发率即趋于稳定,因此,本研究中萌发率的统计持续至第12d,每天下午记录发芽种子数,萌发试验结束后计算萌发率及萌发率达50%所需的时间(To)。在萌发试验的第0、4、8和12d用体式显微镜观察CK1、ABA处理和GA₃处理的完整种子、种子横切面(平行于种脐的切开面)、种子纵切面(垂直于种脐的切开面),记录种子的生长形态图。
萌发率(%)=正常发芽的种子数/30×100
1.3项目测定及方法
1.3.1ROS测定过氧化氢(H₂O₂)含量测定采用试剂盒(苏州科铭生物技术有限公司)进行测定,超氧阴离子(O₂·)含量测定采用羟胺氧化法(高俊凤,2006)。
1.3.2抗氧化物质测定参照王学奎(2006)的方法采用愈创木酚法测定过氧化物酶(POD)活性、紫外吸收法测定过氧化氢酶(CAT)活性。超氧化物歧化酶(SOD)活性和抗坏血酸(AsA)含量采用试剂盒(苏州科铭生物技术有限公司)进行测定,测定方法参考说明书。
1.3.3植物激素含量检测采用酶联免疫法(ELISA)测定圆齿野鸦椿种子内ABA、GA₃、生长素(IAA)、ZR、油菜素甾醇(BR)和茉莉酸(JA)含量(Yang et al.,2001)。
1.3.4可溶性糖(SS)与可溶性蛋白(SP)含量测定SS含量采用试剂盒(苏州科铭生物技术有限公司)进行测定,SP含量采用考马斯亮蓝G-250法(王学奎,2006)进行测定。
1.4统计分析
试验数据采用Excel2010进行统计,采用SPSS22.0进行Duncan's多重比较和Spearman相关分析。使用多镜头图像分析系统(Mshot image analysissys-tem)、Origin2018及Adobe Photoshop2020制图。
2结果与分析
2.1ABA与GA₃处理下圆齿野鸦椿种子萌发的差异
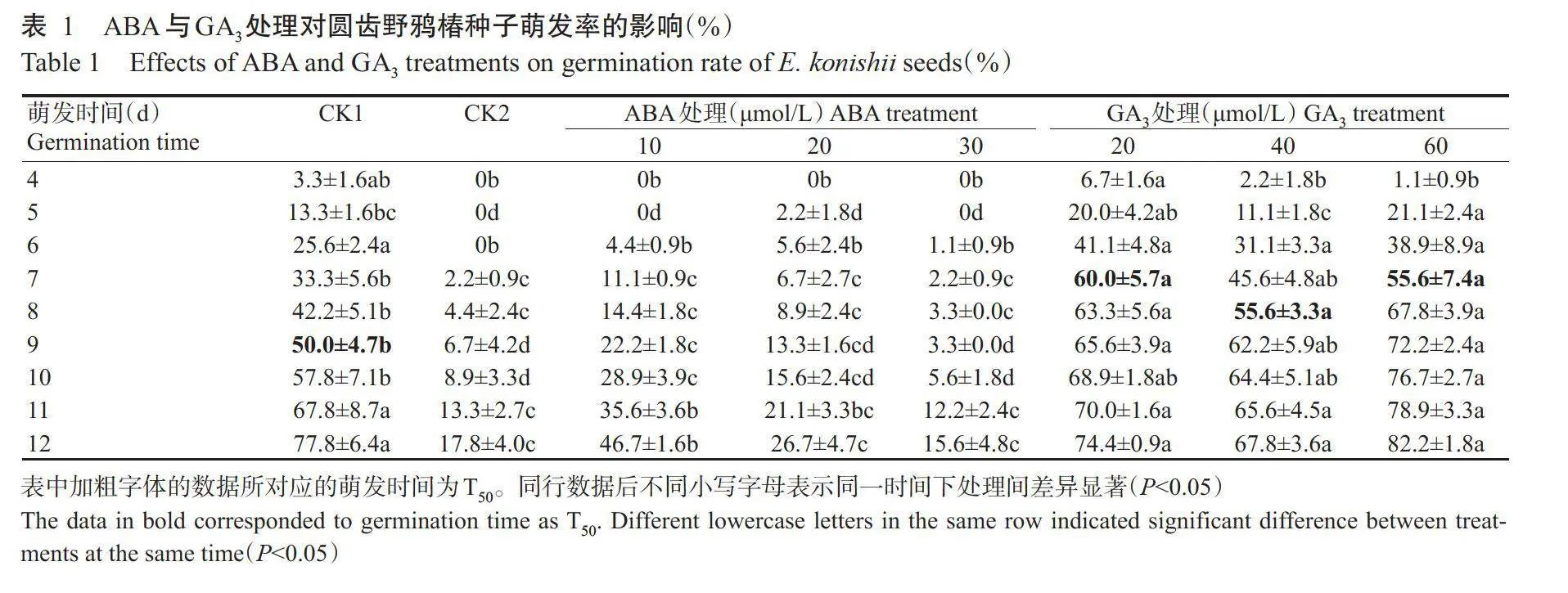
2.1.1种子萌发率的差异本研究发现,在萌发前3d所有处理的种子萌发率均为0。在第6~10d,3个不同浓度GA₃处理的种子萌发率均高于其他处理;而在第11~12d时,3个不同浓度GA₃处理的种子萌发率与CK1无显著差异(Pgt;0.05,下同),但CK1和CA₃处理的种子萌发率均显著高于3个ABA处理和CK2(表1)(Plt;0.05,下同)。ABA处理在第5d时才开始有少量种子萌发,其萌发率始终低于CK1和GA₃处理,且浓度越高萌发率越低。CK2的种子在第7d时才有少量种子萌发,其萌发率变化与30μmol/LABA处理相似,始终低于其他处理。CK1的种子的T₃₀为9d,萌发率最高达77.8%;所有浓度的GA₃处理均可缩短T₅₀,其最高萌发率可达82.2%(第12d);而CK2和ABA处理的种子在整个试验中种子萌发率均未能达50.0%,其中,CK2仅17.8%。
2.1.2种子萌发形态的差异GA₃处理的种子根茎生长纤长,不同浓度GA₃处理间种子的生长形态无明显差异,不同的是,CK1的种子胚根生长粗壮(图1)。在ABA处理中,多数种子的胚根在第8d时胚根仍未突破种皮,直至第12d时,10和20μmol/L ABA处理的部分种子才开始萌发,但胚根和胚轴生长短小,30μmol/LABA处理的种子生长最为缓慢,直至第12d时大部分种子胚根仍未突破种皮。
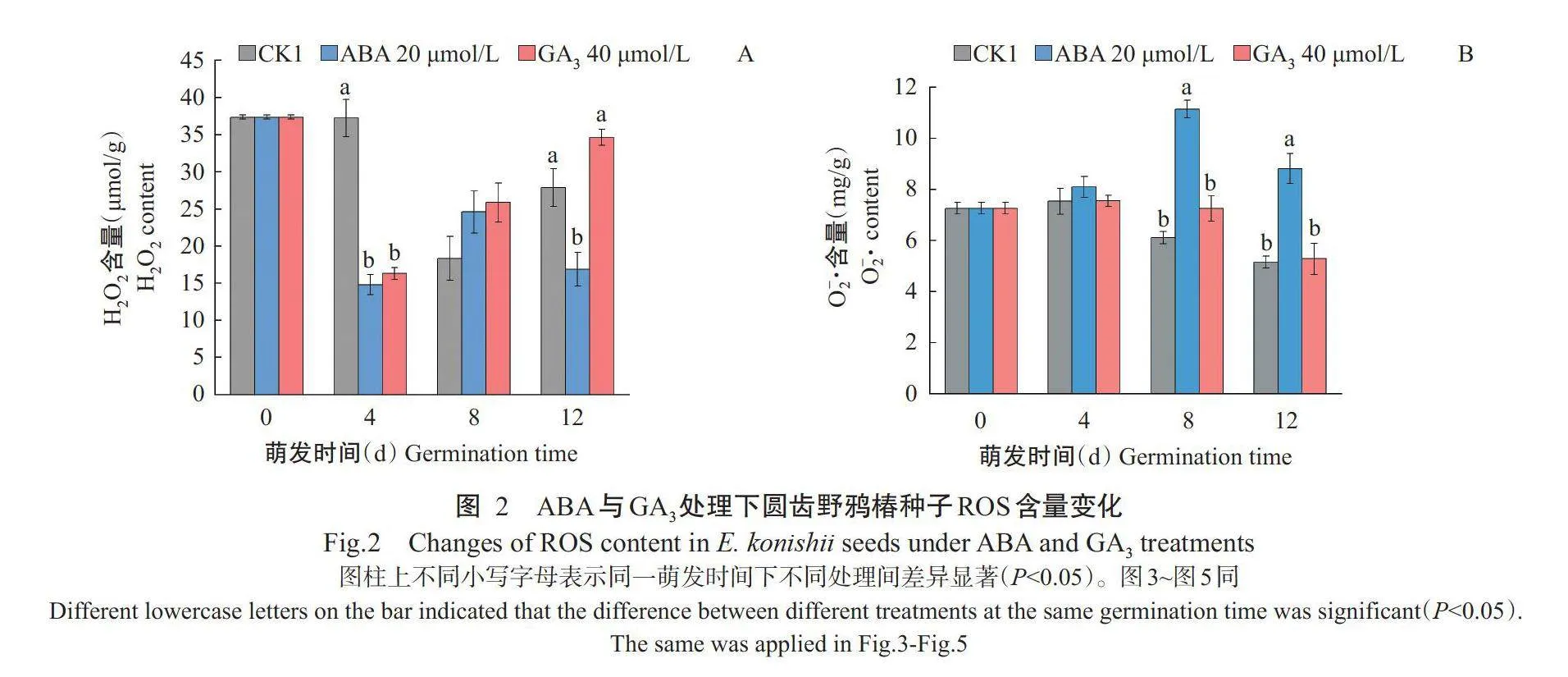
2.2ABA与GA₃处理下圆齿野鸦椿种子ROS含量的差异
如图2-A所示,在萌发第4d,GA₃和ABA处理的H₂O₂含量均显著低于CK1;萌发第12d,ABA处理的H₂O₂含量显著低于CK1,而GA₃的H₂O₂含量高于CK1,但与CK1间无显著差异;随萌发时间进行,CK1和GA₃处理的H₂O₂含量总体上均呈先降低后升高的变化趋势,不同的是GA₃处理的变化启动更早。如图2-B所示,在萌发第8和12d,ABA处理O₂·含量均显著高于CK1和GA₃处理,而CK1与GA₃处理间的O2-含量差异不显著。
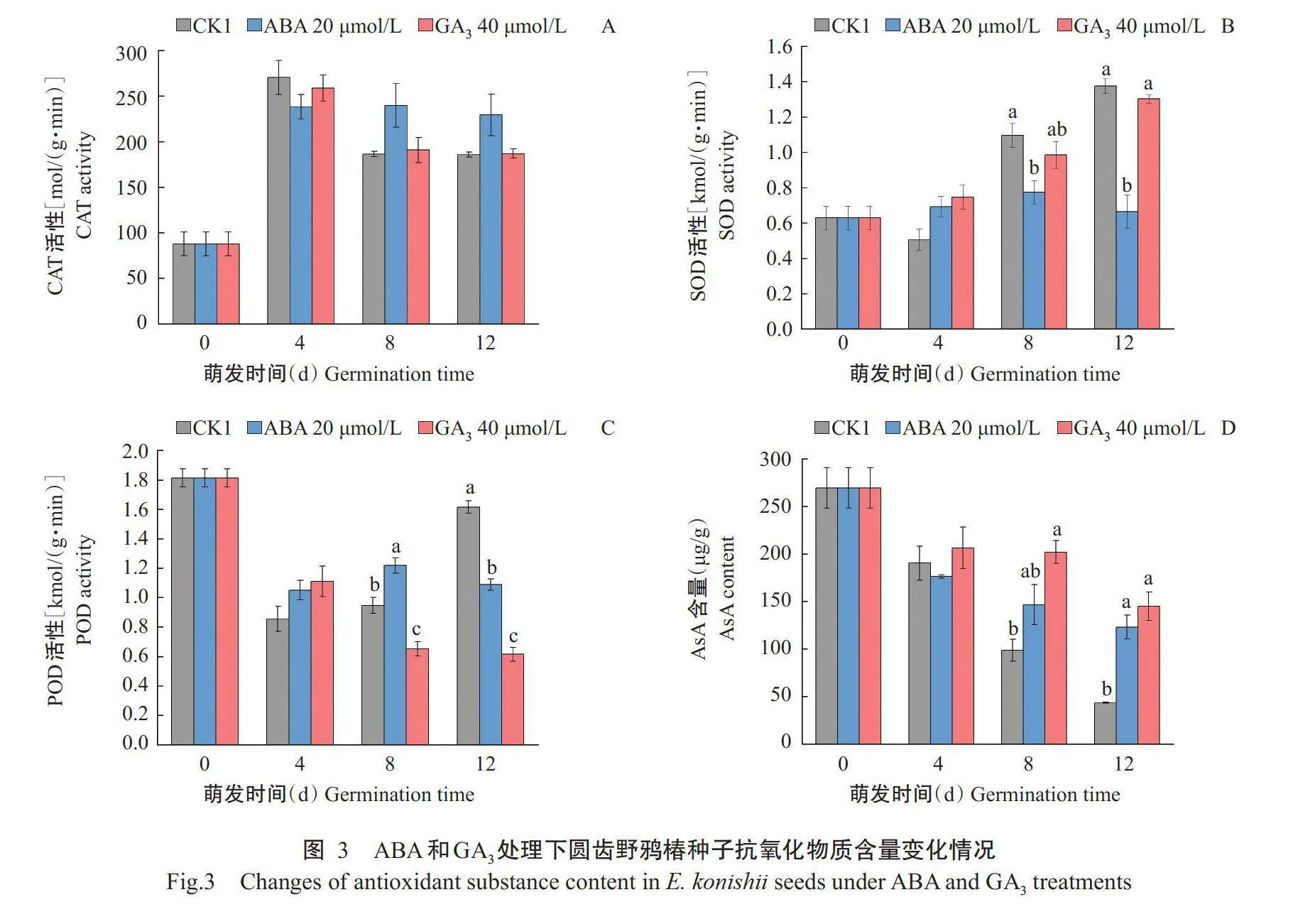
2.3ABA与GA₃处理下圆齿野鸦椿种子抗氧化物质含量的差异
如图3-A所示,3个处理间的CAT活性在整个萌发过程中均无显著差异;不同的是,ABA处理的CAT活性在整个萌发过程均较高,而CK1和GA₃处理则在第8d下降。在萌发第8d,ABA处理的SOD活性显著低于CK1,萌发第12d显著低于CK1和GA₃处理,而2个萌发时间CK1与GA₃处理间则差异不显著;另外,与ABA处理不同,CK1和GA₃处理的SOD活性均随萌发时间的推进而逐渐升高(图3-B)。在萌发第0~4d,3个处理间的POD活性无显著差异,在萌发第8和12d时,GA₃处理显著小于其他处理;与萌发0d相比,在萌发第4d所有处理的POD活性均大幅度下降,在第4~12d,CK1呈逐步上升趋势,GA₃处理呈逐步下降趋势,ABA处理的POD活性变化相对较平缓(图3-C)。与POD活性变化相似,3个处理间的AsA含量在第0~4d时无显著差异,但在萌发第8和12d,ABA和GA₃处理的AsA含量均高于CK1,且3个处理的AsA含量均随萌发时间推进而持续降低,其中,CK1下降幅度最大,ABA处理次之,GA₃处理下降幅度最小(图3-D)。
2.4ABA与GA₃处理下圆齿野鸦椿种子植物激素含量的差异
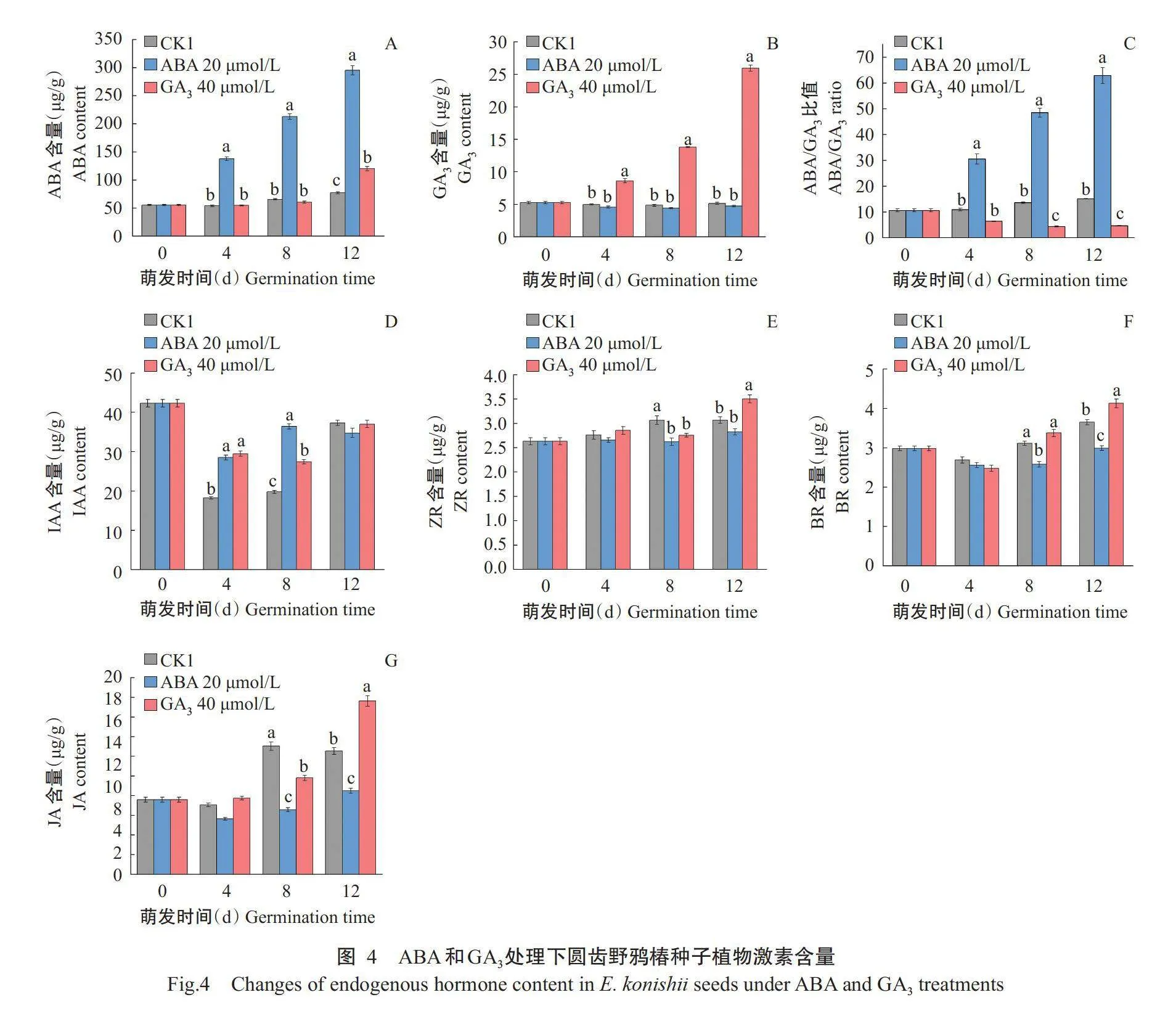
在外施ABA和GA₃处理下,圆齿野鸦椿种子萌发过程中植物激素发生不同的变化(图4)。外施ABA可显著增加圆齿野鸦椿种子内源ABA含量,同样地,外施GA₃可显著增加圆齿野鸦椿种子内源GA₃含量。外施ABA处理的ABA/GA₃比值在萌发过程中均显著高于CK1和GA₃处理;GA₃处理在萌发时间第8和12d均显著低于CK1和ABA处理。在萌发第4d,外施ABA和GA₃处理的种子内源IAA含量均显著高于CK1;而在萌发第8d,IAA含量则表现为ABA处理gt;GA₃处理gt;CK1;在萌发第12d,3个处理间IAA含量无显著差异。在萌发第8和12d,外施ABA处理的BR和JA含量显著低于CK1,同时ABA处理的BR和JA含量也显著低于外施GA₃处理;外施GA₃处理在萌发第8d时ZR和JA含量显著低于CK1,在萌发第12d,外施GA₃处理ZR、BR和JA含量显著高于CK1。
2.5ABA与GA₃处理下圆齿野鸦椿SS和SP含量的差异
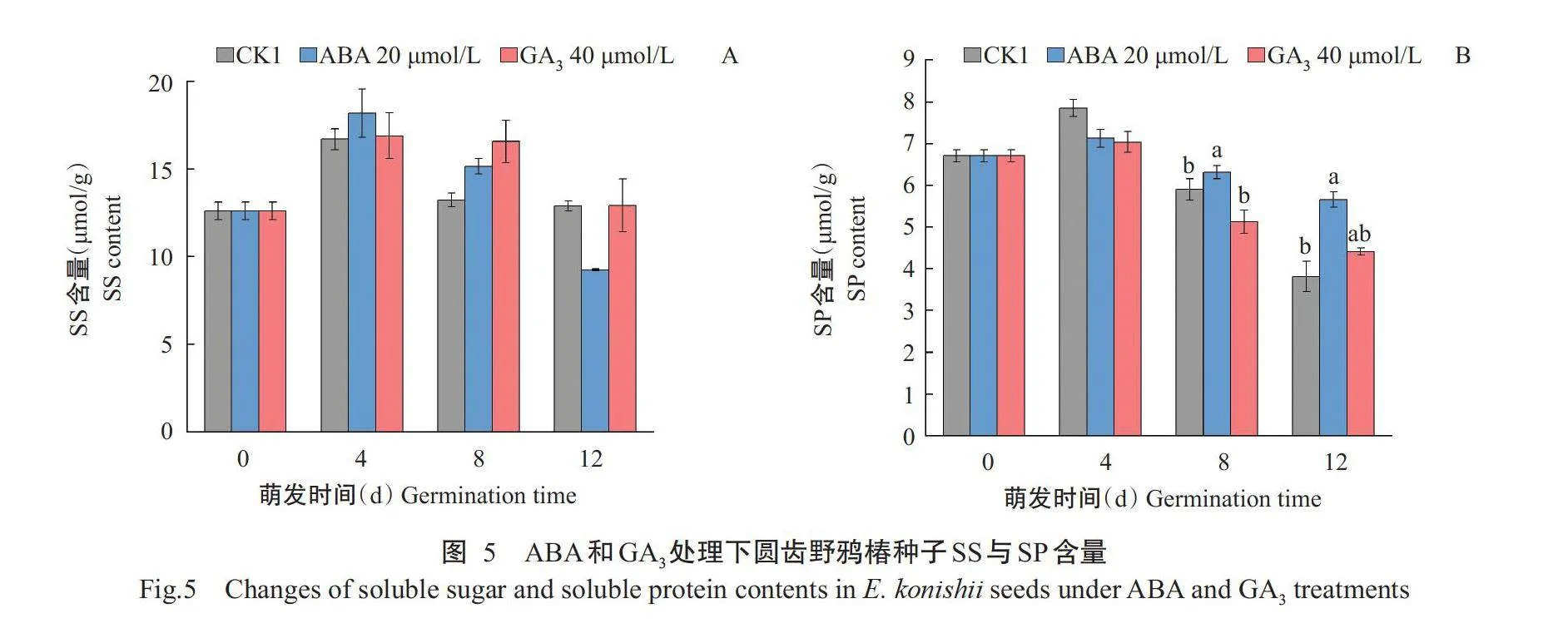
随萌发时间的延长,CK1和ABA处理的SS含量均呈先上升后持续下降的变化趋势,GA₃处理的SS含量呈上升—平稳—下降的变化趋势,在整个萌发过程中3个处理间的SS含量无显著差异(图5-A)。3个处理的SP含量在萌发过程中均呈先上升后下降的变化趋势,不同的是,CK1和GA₃处理的SP含量下降幅度较ABA处理大(图5-B)。
2.6ABA与GA₃处理下圆齿野鸦椿种子萌发过程中各指标的相关分析
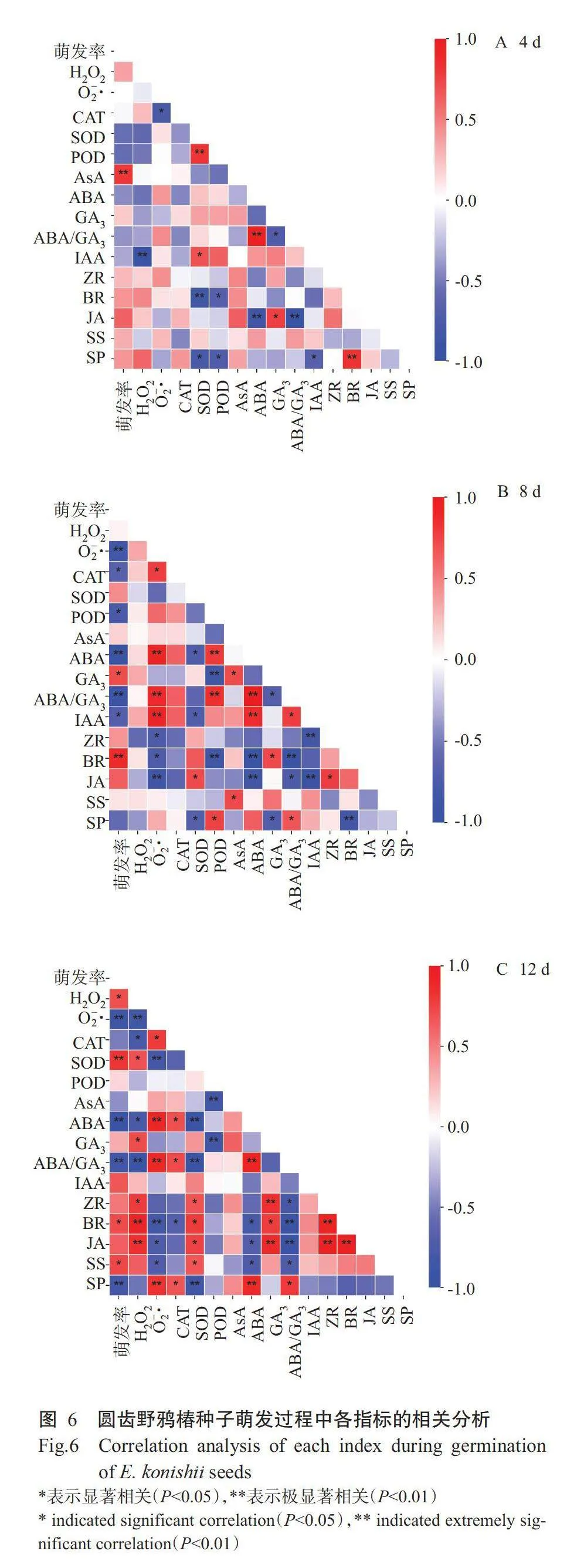
为进一步探究外源ABA与GA₃处理下圆齿野鸦椿种子的生理活动与萌发特性的关系,将种子的萌发率与各指标进行相关分析。如图6所示,方块颜色越来越鲜艳,说明指标间的相关性越强。在萌发第4d,萌发率仅与AsA呈极显著正相关(Plt;0.01,下同),而与其他指标无显著相关性(图6-A)。在萌发第8d,萌发率与GA₃呈显著正相关,与BR呈极显著正相关,与CAT、POD和IAA呈显著负相关,与O₂·、ABA、ABA/GA₃呈极显著负相关(图6-B)。在萌发第12d,萌发率与H₂O₂、BR和SS呈显著正相关,与SOD呈极显著正相关,与O₂·、ABA、ABA/GA₃和SP呈极显著负相关(图6-C)。同时,在整个萌发过程中JA与ABA、ABA/GA₃呈显著负相关;在萌发第8和12d,BR与GA₃均呈显著正相关,与O2-、ABA、ABA/GA₃呈显著负相关;O·与CAT、ABA、ABA/GA₃均呈显著正相关,与JA呈显著负相关;ABA与ABA/GA₃呈极显著正相关,与SOD呈显著负相关。
3讨论
3.1去除外种皮对圆齿野鸦椿种子萌发的影响
造成圆齿野鸦椿种子休眠的原因主要有3种:自然成熟后种胚尚未发育完全;种子内含活性抑制物质;种皮的机械障碍(游双红,2013)。采用湿沙冷层积对圆齿野鸦椿种子进行处理后,萌发率峰值发生在层积120d,但仅48.0%,层积时间超过120d,则萌发率下降(张莉梅等,2015)。另外,将圆齿野鸦椿种子经浓硫酸和赤霉素浸泡后,再进行9个月的变温(4℃90d→25℃90d→4℃90d)层积处理仍未彻底解除种子的休眠,发芽率仅27.50%(游双红,2013)。目前,普遍采用的方法是将采集的种子进行沙藏自然层积450d左右,至第3年取出播种,萌发率可达75%左右,且萌发快而整齐(邹双全,2018)本研究采用层积210d种子进行试验,结果发现在萌发前如未去掉坚硬外种皮,在第12d时萌发率仅17.8%,去除坚硬外种皮的种子在第12d时萌发率可达77.8%,与自然层积450d后的萌发率相近。由此说明,层积至210d的大部分圆齿野鸦椿种子种胚已发育完全,可能是由于硬种皮的机械障碍或种皮内活性抑制物的影响而未能萌发。
3.2外源添加ABA与GA₃对圆齿野鸦椿种子萌发的影响
ABA和GA₃是拮抗调节种子发育的主要激素,通过其活性水平及信号传导等影响种子胚胎的发生、成熟、休眠及萌发(宋松泉等,2023)。ABA具有诱导和维持种子初生休眠的作用,GA₃则具有打破种子休眠、促进种子生长的作用;通过激活ABA合成相关基因NCED2、NCED5、NCED6和NCED9ABA,以及失活GA₃合成相关基因AtGA33oxl和AtGA320ox1可诱导拟南芥种子的休眠,反之则促进种子休眠的解除和萌发,本研究中也发现,ABA浓度越高,对圆齿野鸦椿种子萌发的抑制作用越强,与CK1相比,添加GA₃能有效加快圆齿野鸦椿种子萌发,缩短Ts₀,促进根茎生长,与四川红杉(Larix mas-tersiana)种子试验得出结果(莫丹等,2021)一致。本研究中纯水、20μmol/LABA、40μmol/LGA₃处理的SP和SS含量均在萌发第4d上升之后持续下降,可能与种子在萌发后启动各类生理活动的酶种类增多,需要合成大量的SP和SS为种子萌发做准备(李韦瑶等,2022),在萌发第4d后SP水解产生氨基酸,用于新蛋白质的合成和呼吸氧化提供碳架,SS分解产生能量以供种子萌发(王恒等,2022)。外源ABA处理的圆齿野鸦椿种子SP含量下降缓慢,且在萌发形态上表现为根茎生长短小,可能与施加ABA抑制种子萌发而降低蛋白酶对SP的催化分解有关(周述波等,2005)。
3.3外源添加ABA和GA₃对圆齿野鸦椿种子内源激素与ROS的影响
本研究相关分析结果中,随着萌发时间的延长,呈显著性相关的指标逐渐增多,尤其在萌发试验中后期,即萌发第8和12d,说明种子内激素及ROS相关生理活动随着萌发的进行越来越活跃。植物激素包括GA₃、ABA、JA、ZR、BR、乙烯和水杨酸等既相互独立又协同调控植物的生长发育及对环境的适应过程。已有大量报道表明,种子萌发在很大程度上取决于ABA和GA₃的合成与分解代谢之间的动态平衡(Seo et al.,2006)。在萌发试验中后期,圆齿野鸦椿种子的萌发率和内源JA均与ABA、ABA/GA₃呈显著负相关;已有研究发现紫荆(Cercis chinensis)"" 种子休眠解除过程中ABA含量降低,JA含量上升,JA/ABA呈上升趋势(高云鹏,2020),可见,JA可能对圆齿种子的萌发具有一定的促进作用,而外源ABA的施加则显著降低了种子萌发过程中JA含量。同时,种子萌发率与GA₃和BR均存在显著正相关关系。BR是植物体内甾醇类化合物的总称,在植物光形态建成、逆境胁迫、生殖发育、细胞分裂和分化等方面具有重要作用(黎家和李传友,2019),推测BR具有促进圆齿野鸦椿种子萌发的作用,可能还与GA₃存在协同关系,关于BR在圆齿野鸦椿种子休眠和萌发中的作用机制还需进一步研究。
ROS一方面是有氧代谢的有毒副产物,过量产生会引起细胞死亡;另一方面,当ROS含量控制在一定范围时又可作为信号分子参与生物对环境的感知过程(Mittler,2002),如参与种子的休眠和萌发(Ishibashi et al.,2017;Bally,2019)。本研究发现,圆齿野鸦椿种子内的O₂·在萌发中后期均与萌发率呈极显著负相关,说明此时O₂·过量积累反而会抑制圆齿野鸦椿种子萌发(杨雲雲等,2021)。同时,植物激素BR和JA均与O₂·呈显著负相关,在黑麦草(Loliumperenne)(鲜靖苹等,2019)及番茄(Solanum lycopersicum)(范翠枝等,2021)种子研究中发现外源添加茉莉酸甲酯(MeJA)和BR可降低种子内O₂的积累,而圆齿野鸦椿种子萌发过程中BR和JA含量与萌发率呈显著正相关,与O₂·含量呈显著负相关,说明BR和JA对种子萌发的促进作用可能与对O₂·的清除有关。SOD是存在于植物细胞中最重要的清理自由基的酶,可减轻O₂·对植物体的毒害作用,在圆齿野鸦椿种子萌发试验中后期SOD与ABA呈显著负相关,间接说明SOD活性的增加可减少抑制物质的合成,从而提高萌发率(夏军等,2019)。关于圆齿野鸦椿种子萌发过程中O₂·与ABA的作用关系有待进一步研究。
4结论
添加GA₂能有效缩短圆齿野鸦椿种子萌发率达50%的时间,促进根茎生长,添加ABA能有效抑制圆齿野鸦椿种子萌发。O₂·与ABA的积累对圆齿野鸦椿种子萌发存在抑制作用,BR和JA对种子萌发的促进作用可能与对O₂·的清除有关。
参考文献:
范翠枝,吴馨怡,关欣,郑春芳,赵海燕,顾志壮,刘伟成,陈军,郑青松.2021.油菜素内酯浸种对盐胁迫番茄种子萌发的影响及其生理机制[J].生态学报,41(5):1857-1867.[Fan CZ,WuXY,Guan X,Zheng CF,ZhaoHY,GuZ Z,Liu WC,Chen J,Zheng QS.2021.Concentration effects and its physiological mechanism of soaking seeds with brassinolide on tomato seed germination under salt stress[J].Acta Ecologica Sinica,41(5):1857-1867.]doi:10.5846/stxb201905090946.
高俊凤.2006.植物生理学实验指导[M].北京:高等教育出版社:167-169.[Gao JF.2006.Experimental instruction in plant physiology[M].Beijing:Higher Education Press:167-169.]
高云鹏.2020.紫荆种子休眠解除过程中生理生化变化及分子机理研究[D].南京:南京林业大学.[Gao YP.2020.Study on physiological and biochemical changes and molecular mechanism in Cercis chinensis seeds during dor-mancy releasing[D].Nanjing:Nanjing Forestry University.]
黎家,李传友.2019.新中国成立70年来植物激素研究进展[J].中国科学:生命科学,49(10):1227-1281.[LiJ,LiC"Y.2019.Seventy-year major research progressin planthor-mones by Chinese scholars[J].Scientia Sinica(Vitae),49(10):1227-1281.]doi:10.1360/SSV-2019-0197.
李敏,何勇.2023.活性氧在种子萌发过程中的作用研究进展[J].浙江农林大学学报,40(2):254-264.[LiM,He Y.2023.Research progress on the role of reactive oxygen spe-cies in seed germination[J].Journal of Zhejang Aamp;FUni-versity,40(2):254-264.]doi:10.11833j.issn.2095-0756.20220681.
李韦瑶,肖汇川,张碧莹,李杰,苏滢森,崔国文,秦立刚.2022.不同激素对山韭种子萌发特性的影响[J].种子,41(6):76-83.[LiWY,XiaoHC,Zhang BY,LiJ,Su YM,CuiG W.Qin LG.2022.Effects of different hormones on seed germination characteristics of Allimn senescens L.[J].Seed,41(6):76-83.]doi:10.16590jcnki.1001-4705.2022.06.076.
梁文贤,倪林,邹小兴,黄维,邹双全.2018.野鸦椿属植物化学成分和药理活性研究进展[J].中草药,49(5):1220-1226.[Liang WX,Ni L,Zou XX,Huang W,Zou SQ.2018.Research progress on chemical constituents ofEus-caphis and their pharmacological effects[J].Chinese Tra-ditional and Herbal Drugs,49(5):1220-1226.]doi:10.7501/j.issn.0253-2670.2018.05.034.
莫丹,齐芳,锁才序,蒲光兰,向双.2021.四川红杉(Larix mastersiana)种子快速萌发与幼苗生长技术[J].应用与环境生物学报,27(3):677-685.[Mo D,QiF,Suo CX,Pu GL,Xiang S.2021.Studieson seed rapid germination"and seedling growth technology of Larix mastersiana[J]Chinese Journal of Applied and Environmental Biology,27"(3):677-685.]doi:10.19675/j.cnki.1006-687x.2021.01007.
宋松泉,唐翠芳,雷华平,费思恬,陈海波.2023.ABA调控种子发育的研究进展[J].广西植物,43(9):1553-1567.[Song SQ,Tang CF,Lei HP,FeiS T,Chen HB.2023.Research progress on seed development regulated by ABA[J].Guihaia,43(9):1553-1567.]doi:10.11931/guihaia."gxzw202303053.
王恒,彭嵩敏,孙吉康,韩文军,杜哲.2022.外源蔗糖调节萌发早期蚬壳花椒种子氧化—还原状态的相关机制研究[J].植物生理学报,58(8):1587-1597.[Wang H,Peng sM,Sun JK,Han WJ,Du Z.2022.Study on redox state regulated by exogenous sucrose during the early germina-tion of Zanthoxylum dissitum[J].Plant Physiology Jour-nal,58(8):1587-1597.]doi:10.13592/j.cnki.ppj.100014
王学奎.2006.植物生理生化实验原理和技术[M].第2版."北京:高等教育出版社:186-188.[Wang XK.2006.Prin-ciple and technology of plantphysiological and biochemi-cal experiments[M].The20dEdition.Beijing:Higher Edu-cation Press:186-188.]
夏军,时晓娟,郝先哲,李楠楠,田雨,李军宏,罗宏海.2019.低温对不同基因型棉种萌发过程中酶活性及激素含量的影响[J].植物生理学报,55(9):1291-1305.[Xia J,Shi XJ,Hao XZ,LiNN,TianY,LiJH,LuoH H.2019.Effects of low temperature on enzyme activity and hor-mone conent in germination of different genotypes of co-tton seeds[J].Plant Physiology Journal,55(9):1291-1305.]doi:10.13592/j.cnki.ppj.2019.0322.
鲜靖苹,王勇,张平.2019.外源茉莉酸甲酯对镉胁迫下多年生黑麦草种子萌发及幼苗生理的影响[J].西北师范大学学报(自然科学版),55(6):74-82.[Xian JP,Wang YZhang P.2019.Effects of exogenous methyl jasmonate on seed germination physiological characteristics of Loliumpe-renne under cadmium stress[J].Journal of Northwest Nor-mal University(Natural Science),55(6):74-82.]doi:10."16783/j.cnki.nwnuz.2019.06.014
阳先波,姜雪茹,吴慧平,再芳,涂淑萍,蔡军火,张隆.2022.不同始温浸种对圆齿野鸦椿休眠种子萌发特性的影响研究[J].江西农业大学学报,44(4):929-937.[Yang XB,Jiang XR,Wu HP,Ran F,Tu SP,Cai JH,Zhang L.2022.Effects of soaking seeds at different initial tempera tures on germination characteristics of Euscaphis konishii dormant seeds[J].Acta Agricultura Universitatis Jiangxien-sis,44(4):929-937.]doi:10.13836jiau.2022092
杨雲雲,陈鑫,陈启洲,卢芳,徐晨,杨洪涛,苏佩佩,刘晓龙2021.脱落酸对水稻种子萌发期耐高温胁迫的诱抗效应[J].华北农学报,36(3):185-194.[Yang YY,Chen X,Chen QZ,Lu F,Xu C,Yang HT,Su PP,Liu XL.2021.Pri-ming effects of abscisic acid on high temperature stress to-lerance in rice at seed germination stage[J].Acta Agricul-tura Boreali-Sinica,36(3):185-194.]doi:10.7668/hbnxb."20191968.
游双红.2013.圆齿野鸦椿种子休眠机理及催芽技术研究[D].南昌:江西农业大学.[You SH.2013.The dormancy mechanism and stratification technique of Euscaphis koni-shiiseeds[D].Nanchang:JiangxiAgricultural University.]
张莉梅,张子晗,刘源,赵青,喻方圆.2015.低温层积过程中圆齿野鸦椿种子的生理生化变化[J].种子,34(7):37-40.[Zhang LM,Zhang ZH,LiuY,Zhao Q,YuFY.2015.The biochemical and physiological changes of Euscaphis konishii Hayata seeds during the period of stratification[J].Seed,34(7):37-40.]doi:10.16590/J.cnki.1001-4705.2015.07.037
周述波,林伟,萧浪涛,李再军,童建华.2005.外源GA₃和ABA对杂交水稻种子萌发的影响[J].湖南农业大学学报(自然科学版),31(3):269-271.[Zhou SB,Lin W,Xiao LT,LiZJ,Tong JH.2005.The effect of exogenous GA₃and ABA on germination of hybrid rice seeds[J].Journal of Hunan Agricultural University(Natural Sciences),31(3):269-271.]doi:10.13331/j.cnkijhau.2005.03.011.
邹双全.2018.圆齿野鸦椿研究[M].北京:科学出版社:31-37.[ZouSQ.2018.Study on Euscaphis konishi[M].Bei-jing:Science Press:31-37.]
Amooaghaie R,Ahmadi F.2017.Triangular interplay between ROS,ABA and GA3in dormancy alleviation of Bumium persicum seeds by cold stratification[J].Russian Journal of Plant Physiology,64(4):588-599.doi:10.1134/S1021443717040021.
Bailly C.2019.The signalling role of ROS in the regulation of seed germination and dormancy[J].The Biochemical Jour-nal,476(20):3019-3032.doi:10.1042/BCJ20190159.
Bian FY,Su JR,Liu WD,LiSF.2018.Dormancy release and germination of Taxus yummanensis seeds during wet sand storage[J].Scientific Reports,8(1):3205.doi:10.1038/s41598-018-21469-9.
Finch-Savage WE,Leubner-Metzger G.2006.Seed dormancy and the control of germination[J].New Phytologist,171(3).doi:10.1111/j.1469-8137.2006.01787.x.
Ishibashi Y,Aoki N,Kasa S,Sakamoto M,Kai K,Tomokiyo R,Watabe G,Yuasa T,Iwaya-Inoue M.2017.The interre-lationship between abscisic acid and reactive oxygen spe-cies plays akey role in barley seeddormancy and germina-tion[J].Frontiers in Plant Science,8(3):275.doi:10.3389/fpls.2017.00275.
Jia ZC,Zhao BB,Liu SA,Lu ZG,Chang B,Jiang HR,Cui H,He QS,Li WX,Jin B,Wang L.2021.Embryo tran-scriptome and miRNA analyses reveal the regulatory net-work of seed dormancy in Ginkgo biloba[J].Tree Physio-logy,41(4):571-588.doi:10.1093/treephys/tpaa023.
Mittler R.2002.Oxidative stress,antioxidants and stress tole-rance[J].Trends in Plant Science,7(9):405-410"Seo JH,Choi YH,Yoo MY,Hong KS,Lee BH,Yon GH.,Kim YS,Kim YK,Ryu SY.2006.Isolaion of dihydro-phaseic acid from seed extract of Nelumbo mucifera[J]"Korean Journal ofPharmacognosy,37(4):290-293
Yang JC,Zhang JH,Wang ZQ,ZhuQS,Wang W.2001.Hor-monal changes in the grains of rice subjected to water stress during grain filling[J].Plant Physiology,127(1):315-323.doi:10.1104/pp.127.1.315.
(责任编辑 邓慧灵)
猜你喜欢
江苏农业科学(2016年8期)2017-02-15 20:45:02
江苏农业科学(2016年8期)2017-02-15 20:12:29
天津农业科学(2016年12期)2017-01-11 19:50:58
现代农业科技(2016年20期)2016-12-20 09:14:59
现代检验医学杂志(2016年1期)2016-11-12 13:19:38
现代园艺(2016年17期)2016-10-17 06:52:18
中国科技博览(2016年14期)2016-04-25 02:04:42
山东医药(2015年38期)2015-12-07 09:12:28
食品工业科技(2014年13期)2014-03-11 18:17:06
食品科学(2013年24期)2013-03-11 18:30:53
