
烟草糖苷水解酶GH3基因家族鉴定与表达分析
2024-06-28 15:30张贞宇罗正飞杨帅顾江南尹协韦银龙聂琼
山东农业科学 2024年4期
张贞宇 罗正飞 杨帅 顾江南 尹协 韦银龙 聂琼
摘要:糖苷水解酶CH3基因家族在植物的糖代谢和生长发育中起着重要作用,而烟草CH3基因家族目前尚未见相关鉴定和分析研究报道。本研究采用生物信息学方法在烟草基因组中鉴定CH3家族基因,并对其在不同组织中及逆境条件下的表达模式进行了分析。结果表明,在普通烟草K326的基因组中共鉴定出35个CH3基因,其中23个被定位在14条染色体上,存在6对共线性基因;CH3家族蛋白均为热稳定性蛋白,其基因上游启动子区均包含多个激素和胁迫响应元件。转录组数据表明,CH3家族基因大多数在烟苗茎尖高表达,少部分在根和茎中高表达,表达量受光照时间影响;烟苗受到干旱胁迫后CH3家族成员都有不同程度的响应。挑选4个NtCH3基因进行qRT-PCR分析,结果表明,NtCH3-5、NtCH3-19、NtCH3-22、NtCH3-26可能参与响应低温以及ABA处理。这可为深入了解烟草CH3基因家族及其功能提供依据。
关键词:烟草;CH3基因家族;糖苷水解酶;生物信息学分析;表达模式
中图分类号:S572:Q781 文献标识号:A 文章编号:1001-4942(2024)04-0009-11
烟草(Nicotiana tabacum L.)是我国重要的经济作物,作为嗜好类用品,香气是评判其质量和商业价值的重要指标。研究表明,烟草中的致香成分主要以游离态和结合态两种形式存在,结合态呈香物质没有香气,必须经过分解释放出游离态呈香物质才能产生香气。结合态潜香物质大部分是以糖苷形式存在,如己醇、苯甲醇、苯乙醇和4-羟基-a-紫罗兰醇等。糖苷态潜香物质在常温常压下虽为非挥发性或半挥发性,但在陈化、加工或燃烧过程中却能被糖苷水解酶水解释放挥发性香气物质,从而影响烟草产品的风味和香气。
糖苷水解酶(glycoside hydrolases,GH)又名糖苷酶,是指能够水解两个或两个以上碳水化合物及碳水化合物和非碳水化合物之间糖苷键的一类酶。国际生物化学和分子生物学联合会(IUBMB)基于底物特异性对糖苷水解酶进行命名,如a-葡萄糖苷酶、B-葡萄糖苷酶、B-木糖苷酶、a-甘露糖苷酶等。碳水化合物活性酶数据库(Carbohydrate-Active-enZYmes Database,http://www.CAZy.org)基于糖苷水解酶的氨基酸序列相似性及三维结构性质,将其分为135个家族。其中,糖苷水解酶3家族(GH3)是一类广泛存在于真核生物和原核生物中的酶,具有B-葡萄糖苷酶和B-木糖苷酶等多种催化活性,能够催化多种糖苷类化合物的水解反应,参与生物体内的多种代谢过程,如糖类代谢、细胞壁合成、激素合成、植物生长发育及香气形成等。在拟南芥中,B-葡萄糖苷酶会通过影响ABA含量来改变植株耐旱性;水稻中的Os4BGlu31被证明通过平衡苯丙烯类化合物、类黄酮和植物激素糖缀合物在胁迫适应中发挥作用;将B-葡萄糖苷酶在烟草中过量表达,提高了烟株生物产量,缩短了生育周期。添加B-葡萄糖苷酶有助于茶、果汁、葡萄酒中存在的糖苷类香气前体水解,进而增加产品的香气,改善产品的品质;B-木糖苷酶可促进葡萄酒酿造过程中糖基化的萜烯类物质水解,增加香气。
目前尚未见烟草GH3基因及其相关功能的报道。因此,本研究利用生物信息学方法在普通烟草K326的基因组中鉴定GH3基因家族成员,系统分析其进化关系、理化性质及基因结构,并利用转录组数据和qRT-PCR技术分析其时空表达模式和胁迫响应模式,为探究该基因家族在烟草糖代谢和香气形成方面的功能奠定基础。
1材料与方法
1.1材料与处理
烟草K326种子由贵州大学烟草品质重点实验室提供。2023年4月10日播种,采用烤烟漂浮育苗方式于28℃光照培养箱(16 h光照/8 h黑暗)中育苗培养45天至五叶一心。
1.2烟草GH3基因鉴定及其编码蛋白理化性质分析
普通烟草基因组序列以及注释文件通过茄科基因组数据库(https://solgenomics.net/)获得,由PFAM(http://pfam-legacy.xfam.org/)下载GH3隐马尔科夫模型(profile hidden markov models,profile HMMs)序列谱(PF00933)。利用HMMER 3.0软件在K326蛋白组文件中检索,将得到的GH3候选成员利用SMART(http://smart.embl-heidel-berg.de/)进行进一步筛选,最终获得烟草GH3家族成员。利用在线网站ExPasy(https://web.expasy.org/protparam/)对烟草GH3家族成员进行理化性质分析,包括氨基酸数目、分子量、蛋白质疏水性值等指标。
1.3烟草GH3家族成员系统进化分析
通过NCBI下载辣椒(Calxucum annum)和拟南芥(Arabidopsis thatiana)蛋白完整数据,同样使用HMMER 3.0鉴定和筛选GH3家族蛋白序列,随后利用MEGA 11对3个物种GH3蛋白进行多序列对比和构建系统进化树。采用邻接法,1 000次重复,利用ITOL(https://itol.embl.de/)进行进化树美化。
1.4烟草GH3蛋白保守结构域和基因结构分析
将烟草GH3蛋白上传至MEME(https://meme-stute.org/meme/tools/meme)网站,保守基序设定为10个,其他采用默认设定。利用软件TBtools进行保守结构域作图并使用GFF注释文件绘制基因结构图。
1.5烟草GH3基因定位、共线性分析及Ka/Ks计算
烟草CH3基因染色体定位利用TBtools软件中的Gene Location Visualize功能进行作图,并显示各个染色体基因密度。利用TBtools软件对烟草GH3基因进行复制分析以及与拟南芥、辣椒的共线性分析。拟南芥基因组数据从NCBI上获得,辣椒基因组数据在Ensemble上获得。利用KaKs_Calculator v2.0软件计算复制基因的Ks和Ka。
1.6烟草GH3基因顺式作用元件分析
从烟草基因组中获取每个CH3基因CDS序列上游2 000 bp序列,使用Plant-CARE数据库(http://bioinformatics.peb.ugent.be/webtools/plant-care/html/)获得GH3基因启动子区的顺式作用元件,并通过TBtools进行可视化。
1.7烟草CH3基因转录组测序分析
在GEO(https://www.ncbi.nlm.ruh.gov/geo/)网站下载GSE95717和GSE214048的表达量数据。GSE95717测序背景:烟草在长日照(18h光照/6 h黑暗)条件下生长至8周,收集光照0、6、12、18 h的根、茎、茎尖样本进行转录组测序。GSE214048测序背景:使用200 mmol/L甘露醇模拟干旱处理五叶一心烟草幼苗,收集处理0、1、2、4、8h的烟叶样本进行转录组测序。采用TBtools软件归一化处理后进行可视化作图,分析烟草GH3基因家族成员的差异表达特征。
1.8烟草CH3基因表达分析
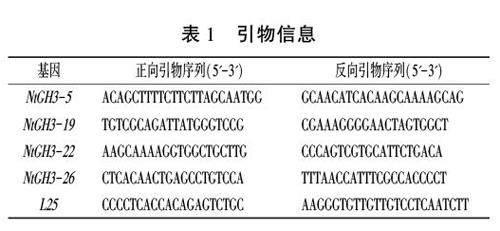
为进一步验证烟草CH3家族成员是否对低温和ABA激素进行响应,挑选4个NtGH3基因(NtGH3-5、NtGH3-19、NtGH3-22和NtGH3-26)进行qRT-PCR分析。选择茁壮且长势相似的五叶一心幼苗进行处理。ABA处理使用100 umol/LABA溶液:冷胁迫则是将幼苗放置在清水培养盘中,然后放入4℃光照培养箱中进行培养。每个处理20株,3次重复,分别在处理0、2、12、24 h取植株叶片,及时存放于-80℃超低温冰箱中保存,用于RNA提取。用Plant RNA Kit纯植物试剂盒(GenStar,R6827)提取总RNA,使用StarScriptⅡRT Mix with gDNA Remover试剂盒(GenStar,A224)获得cDNA。使用Primer-Blast(https://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi)进行特异引物设计,引物信息见表1。用L25作为内参基因,每个反应3次重复,采用2-△△ct法计算基因表达量。
2结果与分析
2.1烟草GH3家族成员鉴定及编码蛋白的理化性质分析
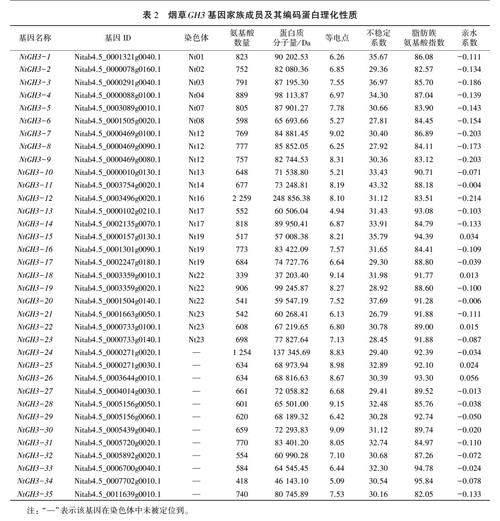
通过HMMER 3.0软件在烟草基因组数据库中检索得到CH3家族成员,去除重复和冗余序列,经在线数据库SMART复筛,共鉴定得到35个烟草GH3家族成员,将其依次命名为NtGH3-I-NtCH3-35(表2)。其中23个成员分别被定位在14条染色体上。
蛋白理化性质分析结果(表2)表明,烟草GH3家族成员的蛋白序列长度在339-2259个氨基酸之间,分子量为37.2-248.9 kDa,其中Nt-GH3-18的氨基酸数量最少,分子量最小:NtGH3-12的氨基酸数量最多,分子量最大。烟草GH3成员中有20个属于碱性蛋白,15个属于酸性蛋白,其中NtGH3-18的等电点最高(9.14),NtGH3-13的等电点最低(4.94)。烟草GH3蛋白的不稳定系数为26.79-43.32,仅NtGH3-11的不稳定系数大于40,较为不稳定,其余均为稳定蛋白。烟草GH3蛋白的脂肪族氨基酸指数为82.05-95.84,表明该类蛋白的热稳定性较好。同时烟草GH3蛋白亲水系数为-0.214-0.056,其中NtGH3-15、NtGH3-18、NtGH3-22、NtGH3-25、Nt-GH3-26为疏水性蛋白,其余30个蛋白均为亲水性蛋白。
2.2烟草GH3家族成员的系统进化分析
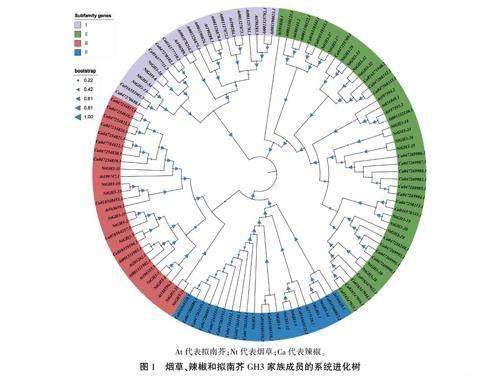
使用HMMER 3.0软件分别在拟南芥和辣椒中鉴定出30、51个GH3基因,使用MEGA 11软件采用最大似然(ML)法构建其与烟草GH3蛋白的邻接(NJ)系统进化树。结果(图1)显示,116个GH3蛋白可分为4类,其中,I类包括4个烟草成员、13个拟南芥成员和5个辣椒成员:Ⅱ类成员最多,包括15个烟草成员、8个拟南芥成员和22个辣椒成员:Ⅲ类成员最少,包括6个烟草成员、2个拟南芥成员和13个辣椒成员:Ⅳ类包括10个烟草成员、7个拟南芥成员11个辣椒成员。同时又可将I和II类分为编码B-葡萄糖苷酶类,Ⅲ、Ⅳ类分为编码B一木糖苷酶类。每一类都包含了3个物种的GH3家族成员,说明GH3家族成员的分化早于3个物种的分化。辣椒家族成员数量较多,说明GH3家族成员在该物种形成后可能经历了多次复制事件。
2.3烟草GH3基因家族成员序列分析及启动子分析
序列相似的基因一般具有相似的生物学功能,为此本研究采用在线网站MEME对烟草GH3家族成员的保守基序进行了分析,结果(图2A)表明NtGH3家族成员均包含4-10个基序,除Nt-GH3-2/和NtGH3-32外均含有Motifl和Motif2,表明这两个基序是GH3家族成员的保守序列。CD Search分析(图2B)显示,NtGH3成员含有糖苷水解酶家族3的保守结构域(Glyco_hydro_3_C),其中16个NtGH3蛋白还含有与木聚糖降解相关的PLN03080保守结构域,19个NtGH3蛋白含有参与葡萄糖降解的PRK15098保守结构域,可据此将其分为B-葡萄糖苷酶和B-木糖苷酶两类,前者的保守基序中多出现Motif5和Motif6,推测这两种保守基序可能决定了糖苷水解酶的功能。NtGH3基因结构分析(图2C)表明,除NtCH3-12外显子为18个以外,其余成员均含有4-12个外显子。
利用Plant-CARE在线软件对NtGH3基因家族成员上游2 000 bp序列进行分析,发现上游序列中有多个与激素响应、光响应和胁迫响应相关的顺式作用元件(图3)。其中,与光反应相关的元件有ACE、G-box、MRE、Spl和GTl-motif;与激素反应相关的元件有:CGTCA-motif、TGACG-mo-tif(MejA)、ABRE(ABA)和P-box(GA3)等;还有与非生物胁迫相关的TC-rich repeats、LTR和MBS等元件。与茉莉酸甲酯(MejA)相关的CGT-CA-motif基序和TGACG-motif以及与脱落酸(ABA)相关的ABRE存在于大多数家族成员中,这意味着大多数NtGH3成员可能参与这两类激素信号传导。
2.4烟草GH3基因定位与共线性分析
染色体定位结果(图4)表明,有23个NtGH3基因分布在14条染色体上,其中12、19、22、23号染色体上各有3个成员,17号染色体上有两个成员,1、2、3、4、7、8、13、14、16号染色体上各有1个成员,12个NtGH3基因未能定位到染色体上。
共线性分析结果表明:23个NtGH3成员中共有6对共线性基因(图5A),主要为大片段复制,没有串联复制的基因对:通过计算,6个复制基因对的Ka/Ks都远小于1(表3),主要为同义替换。烟草分别与拟南芥、辣椒GH3成员的共线性分析发现:9个CaGH3基因与13个NtGH3基因共组成13个基因对,2个AtGH3基因与3个NtGH3基因共组成3个基因对(图5B),说明基因复制可能是烟草基因组GH3基因扩增和进化的动力。
2.5烟草GH3家族基因表达分析
生长8周的烟草幼苗光照0、6、12、18 h的根、茎、茎尖的转录组数据表明,NtGH3基因在不同部位和不同光照时间差异表达(图6A)。NtGH3-2、NtGH3-16、NtGH3-31主要在根中高表达,光照后表达量降低:NtGH3-6、NtGH3-10和NtGH3-11主要在茎中表达,表达高峰分别出现在光照12、12、0h:其余NtGH3基因在茎尖中高表达,表达量随光照时间延长呈先上升后下降的趋势。
使用200 mmol/L甘露醇模拟干旱处理后,烟草幼苗的转录组数据中共检测到23个NtGH3成员(图6B)。干旱胁迫后,NtGH3-2、NtGH3-3、Nt-GH3-5、NtGH3-11、NtGH3-16的表达量下调,Nt-GH3-1、NtGH3-4、NtGH3-6、NtGH3-7、NtGH3-10、NtGH3-12、NtGH3-13、NtGH3-18、NtGH3-19的表达量呈现增加趋势,NtGH3-14、NtGH3-17、NtGH3-20、NtGH3-22的表达趋势为先增加后下降,NtGH3-15、NtGH3-8的表达量表现为先降低后升高又降低的趋势,NtGH3-9、NtGH3-21、Nt-GH3-23表达量不受干旱胁迫的影响。
2.6低温和ABA对部分NtGH3基因表达的影响
对NtGH3家族基因的启动子元件分析发现,大多数基因上游含有低温和ABA响应元件。因此,本试验对烟草幼苗进行24 h低温(4℃)和ABA处理后,分析NtGH3-5、NtGH3-19、NtGH3-22、NtGH3-26四个基因的表达变化。结果(图7)显示,低温胁迫下,4个基因表现出了不同的变化,较长时间的低温处理(24h)使NtGH3-5的表达显著下调,而使NtGH3-22、NtGH3-26的表达显著上调:NtGH3-19的表达量则随处理时间延长呈现先升高后降低的趋势。ABA处理后,NtGH3-5和NtGH3-19的表达量表现出先下降后升高的趋势,而NtGH3-22表现出先升高后降低的趋势,在12h时表达量达到峰值。NtGH3-26对ABA非常敏感,ABA处理后其表达明显受到抑制,仅为处理前的30%。表明这4个基因可能参与了低温和ABA的响应。
3讨论与结论
本研究在普通烟草K326的全基因组范围内鉴定到35个NtGH3基因,根据保守结构域类型可将其分为B-木糖苷酶和B-葡萄糖苷酶两类。NtGH3成员均为热稳定性蛋白,这与在棘孢木霉中的预测结果相似。在拟南芥和辣椒基因组中也分别鉴定到30个和51个GH3家族成员。系统发育树和共线性分析表明,NtGH3成员中有6对共线性基因,复制方式主要为大片段复制。Ka/Ks表示的是异义替换(Ka)和同义替换(Ks)之间的比例,共线性基因对中Ka/Ks都小于0.5,表明NtGH3基因家族进化过程受到了纯化作用。GH3家族的分化可能早于3个物种的形成,但后期辣椒GH3家族成员的分化与烟草的亲缘关系更近。辣椒GH3成员个数明显多于烟草,从染色体数目来推测辣椒CH3家族的复制事件多于烟草。
启动子的作用对于基因表达至关重要,决定了特定蛋白质在特定细胞或组织中何时产生,因此,对启动子序列进行分析,可以揭示基因表达调控的分子机制。NtGH3家族成员上游含有较多的激素响应位点、逆境胁迫应答元件和光反应元件识别位点,推测NtGH3可通过激素调控对外界环境变化做出响应。前人研究发现,植物产生脱落酸和茉莉酸甲酯可作为激活防御机制的信号,而且脱落酸和茉莉酸甲酯与植物香气的形成密切相关。NtGH3家族的大多数成员都含有脱落酸和茉莉酸甲酯响应元件,表明NtGH3可能通过产生茉莉酸甲酯和ABA来响应外界环境变化和促进香气物质的形成。
长日照下不同部位的转录组数据分析发现,大部分基因在烟草幼苗各部位均有表达,但部分成员表达量极低:各部位基因表达量存在差异,大部分成员在茎尖中高表达,少部分在茎与根中高表达。表明NtCH3成员的表达受到日照时间的调控。而NtGH3-21和NtGH3-23可能为烟草GH3基因家族扩张中出现的不进行表达的“假基因”。
渗透调节是植物应对逆境胁迫的重要途径。研究表明糖苷水解酶可提高耐辐射球菌的渗透耐受性。本研究发现烟草GH3基因家族成员包含许多逆境胁迫应答元件,因此探究了NtGH3家族基因在非生物胁迫下的表达模式,结果表明,大多数基因对干旱存在应答,呈现随着胁迫时间延长而上调、下调及先上调后下调表达3种情况:在低温和ABA处理下,NtGH3家族4个基因均有不同程度的响应,变化趋势存在差异,其中NtGH3-22在低温下随处理时间延长表达量逐渐升高,NtGH3-26对外源ABA敏感,短时间内表达量就显著降低。
综上,本研究从烟草基因组中鉴定出GH3基因家族的35个成员,并对其进行了生物信息学分析,确定了烟草GH3家族成员分属于B-葡萄糖苷酶和B-木糖苷酶,在干旱、低温等逆境胁迫响应方面发挥作用。因此,NtGH3基因家族可以作为研究烟草糖代谢和逆境胁迫响应机制的候选基因。本研究结果可为烟草的遗传改良及优良品种选育提供理论基础。
作者贡献:张贞宇是实验的设计和执行人:罗正飞、扬帅及顾江南完成数据分析和论文初稿的写作:尹协参与实验设计和实验结果分析:聂琼是项目的构思者及负责人,指导实验设计、数据分析和论文写作与修改。全体作者都已阅读并同意最终的文本。
猜你喜欢
奥秘(创新大赛)(2023年3期)2023-05-06
湖北农业科学(2017年3期)2017-03-21
中国中药杂志(2017年1期)2017-03-06
浙江中西医结合杂志(2017年2期)2017-01-12
江苏农业科学(2016年5期)2016-07-23
江苏农业科学(2016年4期)2016-06-14
当代化工研究(2016年9期)2016-03-20
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年1期)2015-04-17
声屏世界(2014年6期)2014-02-28
