
蕹菜开花特性和繁育系统初探
2024-05-29 13:56邹琴谭舒婷黄亚郭笑影王新月彭富星陈雅维艾辛
中国瓜菜 2024年5期
邹琴 谭舒婷 黄亚 郭笑影 王新月 彭富星 陈雅维 艾辛

摘 要:為解决蕹菜品种间花期不遇、有性杂交困难的问题,通过对渔场蕹菜、泰国大叶蕹菜、汝城蕹菜和长沙藤蕹4个蕹菜亲本的开花特性和繁育系统观测和11 h?d-1短日照调控蕹菜品种的花期,并开展了蕹菜品种间有性杂交。结果表明,蕹菜自然光周期条件下现蕾期为8—11月份,刈割发芽后短日照诱导11~13 d现蕾,现蕾至开花11 d左右,单花开放时间约12 h;开花当天09:00—13:00花粉活力最高,11:00—13:00柱头可授性最强;渔场蕹菜和长沙藤蕹花粉/胚珠比(P/O值)、杂交指数(OCI值)基本一致,但长沙藤蕹在控制授粉试验中表现为自交不结实;花粉粒为球状或帽状,表面布满尖刺;以籽蕹品种为母本的杂交结实率最高可达75.00%,结籽率高;长沙藤蕹为母本的杂交结实率最高可达6.00%,结籽率为0。综合分析表明,蕹菜繁育系统倾向于异交混合交配型,11 h·d-1短日照处理可以实现蕹菜品种间花期相遇,蕹菜品种间杂交亲和,但长沙藤蕹有可能丧失了结籽能力。
关键词:蕹菜;光周期;花部特征;繁育系统;花粉活力;柱头可授性;杂交育种
中图分类号:S636.9 文献标志码:A 文章编号:1673-2871(2024)05-133-09
Preliminary study on flowering characteristics and breeding system of water spinach
ZOU Qin, TAN Shuting, HUANG Ya, GUO Xiaoying, WANG Xinyue, PENG Fuxing, CHEN Yawei, AI Xin
(Engineering Research Center for Horticultural Crop Germplasm Creation and New Variety Breeding, Ministry of Education/Key Laboratory for Vegetable Biology of Hunan Province/College of Horticulture, Hunan Agricultural University, Changsha 410128, Hunan, China)
Abstract: In order to solve the problem of infertility during flowering period and difficulty in sexual hybridization among water spinach varieties, the flowering characteristics and breeding system of four water spinach parents, namely Yuchang water spinach, Thai water spinach with large leaves, Rucheng water spinach, Changsha vine water spinach were observed, and the flowering period between water spinach varieties was regulated by 11 h·d-1 short day light. Sexual hybridization research was also conducted among water spinach varieties. The results showed that under natural photoperiod conditions, the budding period of water spinach was from August to November. After cutting and sprouting, short day light induced budding for 11 to 13 days, and the budding period lasted for about 11 days until flowering, with a single flower opening time of about 12 hours. On the day of flowering, from 09:00 to 13:00, the pollen vitality was highest, and from 11:00 to 13:00, the stigma had the strongest receptivity. The P/O value and OCI value of Yuchang water spinach are basically the same as those of Changsha vine water spinach, but Changsha vine water spinach showed poor self pollination in the controlled pollination experiment. The pollen grains are spherical or cap shaped, with a surface covered with spikes. The hybrid seed setting rate using the seed water spinach variety as the maternal parent can reach to 75.00% , with a high seed setting rate. The hybrid fruiting rate of Changsha vine water spinach as the female parent can reach to 6.00%, and the seed setting rate is 0. Based on comprehensive analysis, it can be concluded that the water spinach breeding system tends to be a hybrid mating type, and the 11 h·d-1 short day treatment can achieve the encounter of flowering periods among water spinach varieties. The hybridization between water spinach varieties is compatible, but Changsha vine water spinach may have lost its ability to bear seeds.
Key words: Water spinach; Photoperiod; Floral characteristics; Breeding system; Pollen vitality; Stigma acceptability; Hybrid breeding
蕹菜为旋花科番薯属植物,原产中国,主要分布于我国南方各地,是夏秋季主要绿叶蔬菜[1]。蕹菜分为籽蕹和藤蕹,消费者认为藤蕹的口感好,但藤蕹属于专性短日照植物[2],因花期太晚不能结实,无法开展籽蕹与藤蕹的有性杂交育种,严重限制了蕹菜种质创新和新品种选育工作。深入了解植物的开花特性,准确判别植物的繁育系统类型,既是植物杂交选育的前提条件,也直接关系到授粉结实的成功率[3]。植物繁殖生物学研究主要涉及植物的开花特性、花粉活力和柱头可授性测定、繁育系统的确定以及杂交育种体系的构建等,可为植物的有性生殖和种质资源的创新研究奠定基础[4-6]。繁育系统是进化生物学研究中最活跃的领域,它代表了影响下一代有关性表达的特征总和[7]。蕹菜的开花生物学和繁育系统的研究目前还缺乏系统性的报道,笔者通过开展蕹菜开花特性和繁育系统的研究,人工调控光周期诱导蕹菜现蕾开花,调整藤蕹和籽蕹花期相遇,在盛花期了解蕹菜的开花特性和繁育系统,以期为有效开展蕹菜品种间杂交育种奠定理论基础。
1 材料与方法
1.1 试验地点
试验于2023年7—9月在湖南农业大学金山实验基地玻璃温室和露地进行。
1.2 试验材料及其栽培管理
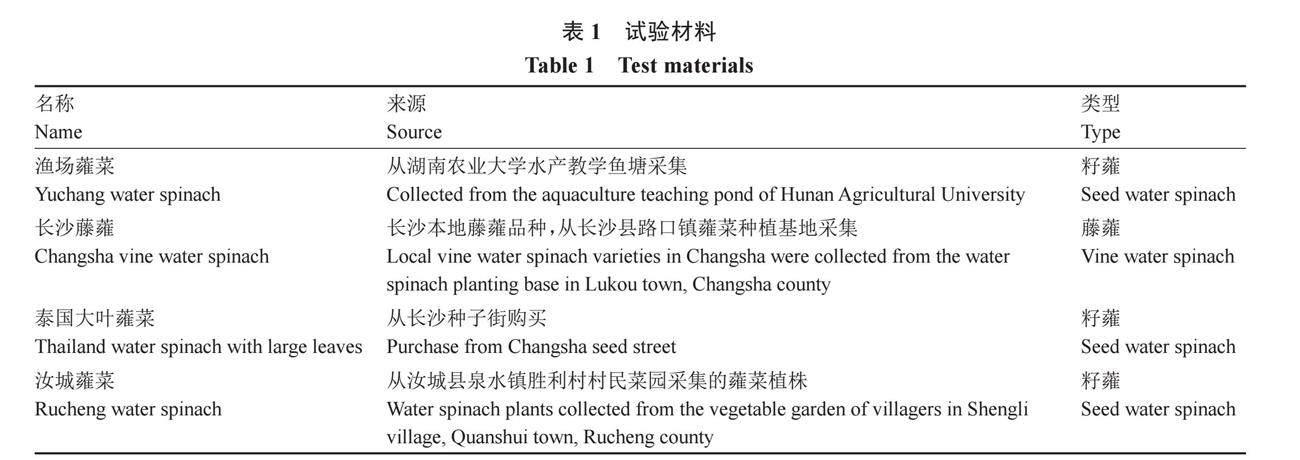
试验材料如表1所示。籽蕹品种播种后选择1株健壮苗,待幼苗长大后用茎段进行扦插繁殖各40株,长沙藤蕹同样选择1株进行茎段扦插繁殖40株,生根后移栽,分露地盆栽和玻璃温室内无土栽培槽种植,两种种植方式每个品种各种植20株,每株挂上标明品种名称的吊牌;露地盆栽的容器为35 cm的塑料盆,栽培槽宽90 cm,双行栽培,行距30 cm,株距45 cm,栽培基质为购买的商品育苗基质。盆栽蕹菜放置在温室外的空地上栽培,每隔3 d施1次0.6%水溶性大量元素肥(N、P、K质量比15∶15∶15),栽培槽内每隔15 d埋施15 g复合肥(N、P、K质量比15∶15∶15),水分管理根据天气、基质水分含量和植株的生长情况及时补充。露地盆栽主要进行蕹菜开花生物学和繁育生物学试验,玻璃温室内采用光周期调控蕹菜的花期,主要开展蕹菜有性杂交工作。露地盆栽的蕹菜每隔20 d左右刈割1次。
1.3 开花生物学特性和花部结构特征
盆栽蕹菜在自然光周期条件下选择10株进行花期观测、花器官性状的测定和繁育系统测定。玻璃温室内种植的蕹菜于2022年7月19日统一刈割后进行11 h·d-1短日照处理(遮光),每个品种选择10株,每天观察并记录蕹菜在人工调控短日照11 h·d-1条件下始蕾和始花所需天数。在盛花期时观察并记录植株花序结构特征、花蕾发育进程、开花动态和开花后不同时期的形态特征、单花花部结构特征、花柱类型,并拍照。随机选择开花前一天的花蕾,从上午9:00开始,每隔2 h观测花药状态和散粉时间,持续观测10 d。
观测并记录每花序的花数,每个品种随机选取10朵完全开放的花朵,用直尺测量花序梗长、花柄长、花萼长、花冠筒长、花瓣长、花冠直径、雌蕊长、雄蕊长,使用游标卡尺测量花序梗粗、花柄粗。观测记录柱头颜色、柱头形态、花柱类型和冠喉颜色。持续10 d观测每个品种当天开放花的花柱类型,每个品种设置3个重复组,最后计算每品种每花柱类型的比率。
1.4 花粉/胚珠比(P/O)和杂交指数(OCI)估算
以温室内种植的渔场蕹菜和长沙藤蕹作为试验材料,各选取3朵花,取将要散粉的花药,置于洁净离心管内,每朵花分别制成2 mL花粉粒悬浮液。用移液枪吸取1 ?L于载玻片上,在光学显微镜下统计花粉量,每个视野中的花粉数不少于40粒,每朵花3次重复,观察花粉数乘以2×103,取平均值计算单花花粉总量。在体视显微镜下用解剖针划开心皮,观察统计胚珠数。以单花的花粉总量除以胚珠数,得到花粉/胚珠比(P/O值)。依據Cruden[8]的标准,对蕹菜繁育系统进行判定。
以温室内种植的渔场蕹菜和长沙藤蕹作为试验材料,按照Dafni[9]的标准,对其进行杂交指数估算,以此判定蕹菜的繁育系统。
1.5 控制性授粉试验
在自然光照条件下,选取长势良好的渔场蕹菜15株,分别做以下处理:(1)不授粉,不套袋,检测自然传粉条件下的结实率;(2)不授粉,不去雄,套袋,检测自花授粉能力;(3)去雄,人工异株授粉,套袋,检测异交结实率;(4)去雄,人工同株授粉,套袋,检测自交结实率;(5)去雄,不授粉,套袋,检测有无融合生殖现象。每处理3株,统计每处理的平均处理花朵数、平均结实数和平均结实率,根据平均结实率判定蕹菜的繁育系统类型。
1.6 花粉粒形态和花粉活力检测
取渔场蕹菜、泰国大叶蕹菜、汝城蕹菜和长沙藤蕹的花,采用荷兰飞纳台式扫描电镜进行蕹菜花粉粒结构的观测。选取温室内种植的渔场蕹菜和长沙藤蕹的花粉,用碘-碘化钾染色法[10]、0.5%TTC染色法[11]、联苯胺-甲萘酚染色法[12]、0.5%MTT法[13]在OLYMPUS金相显微镜下进行花粉活力的测定。采集开花前一天和开花当天09:00、11:00、13:00、15:00、17:00以及开花后一天17:00的花粉,取少许放在干燥洁净的载玻片上,滴加反应液,待充分反应后盖上盖玻片后镜检,观察花粉染色情况。每个品种选择3朵即将开放的花蕾、开花当天的花和开花后一天的花,分别制作3张片子,每片取5个视野进行观测,最后统计3朵花的平均染色率,代表花粉活力高低。
1.7 柱头可授性测定
选取温室内种植的盛花期的渔场蕹菜和长沙藤蕹花朵,将开花前一天和开花当天09:00、11:00、13:00、15:00、17:00以及开花后1 d 15:00的花蕾和花朵,用镊子摘取柱头浸入凹面载玻片中,将载玻片放入OLYMPUS金相显微镜下进行观测。凹面载玻片含有联苯胺-过氧化氢反应液[14](V1%联苯胺∶V3%过氧化氢∶V水=4∶11∶22)溶液,柱头周围的反应液呈现蓝色并有大量气泡出现,气泡越多说明柱头的可授性越强,反之,则越弱。每个品种、每次测定选取5朵花蕾或已开放的花朵,3次重复;观测并记录每个样品的柱头冒泡情况。
1.8 杂交授粉试验
在11 h·d-1光照条件下诱导蕹菜开花并实现花期相遇,以渔场蕹菜、泰国大叶蕹菜、汝城蕹菜作为母本,长沙藤蕹作为父本进行人工去雄杂交。随后又以渔场蕹菜和长沙藤蕹的杂交子一代作为母本,渔场蕹菜和长沙藤蕹分别作为父本进行回交。在开花前一天下午15:00-17:00对花药未裂开的花蕾去雄后进行套袋处理,在开花当天09:00-10:00授粉,授粉后将授粉花的花冠轻捻闭合,使之不受其他花粉干扰。每杂交组合设置3次重复,授粉后记录授粉数、授粉日期,记录结实数,计算3次重复的平均授粉数、平均结实数和平均结实率。
1.9 数据处理
利用Excel 2019进行原始数据的统计和平均值的计算以及相关图表制作,利用SPSS 26.0数据分析软件中的单因素ANOVA进行显著性差异分析。
2 结果与分析
2.1 蕹菜花部结构的差异
蕹菜花为聚伞花序,每花序具1~6朵花,苞片小鳞片状,花梗无毛;萼片近于等长,卵形,顶端钝,具小短尖头,外面无毛;花瓣为连瓣花,呈漏斗状;花冠为白色,冠喉颜色有白色、紫色,每花中有5个雄蕊,不等长,3短2长;花丝基部被毛,子房圆锥状,无毛(图1)。
根据花柱与花药的位置分为长花柱花、短花柱花、等长花柱花3种类型(表2,图1),在观测的蕹菜品种中,发现只有泰国大叶蕹菜和汝城蕹菜有3种花柱类型共存的现象,而渔场蕹菜和长沙藤蕹只有长花柱类型的花。品种间花朵的大小有明显差异(表3),泰国大叶蕹菜和渔场蕹菜花朵大,花冠直径达6.01 cm,长沙藤蕹花朵小,花冠直径3.35 cm,而汝城蕹菜花朵大小居于两者之间,花冠直径4.87 cm。蕹菜冠喉颜色除长沙藤蕹为紫色外,其他3个品种都是白色。柱头颜色在长沙藤蕹中出现紫色和白色,而其他3个品种都是白色。
2.2 不同光周期条件下蕹菜的开花特性比较
在自然光周期条件下,蕹菜现蕾日期在品种间有很大差异(表4),渔场蕹菜和汝城蕹菜现蕾期在9月份,泰国大叶蕹菜10月份现蕾,而长沙藤蕹11月份现蕾,品种间现蕾期最近的相隔11 d,最远的相隔64 d。蕹菜现蕾至开花历经11 d左右。天气晴朗时,在09:00后蕹菜花处于完全开放状态。在人工11 h·d-1短日照调控下,蕹菜始蕾和始花所需天数如表4所示,不同品种蕹菜幼苗从接受短日照开始至现蕾所需时间差异不大,多数植株短日照处理后10~13 d,蕹菜现蕾至开花历经12 d(图2)。蕹菜开花12 h后花冠便全部向内皱缩,在开花2 d花冠皱缩处会出现局部枯萎现象,在开花3 d花瓣呈完全枯萎皱缩状态。在玻璃温室内,天气晴朗时,白天温度32~42 ℃,蕹菜05:00~06:00开花,07:00后蕹菜花全部处于完全開放状态;天气阴冷时,白天温度21~25 ℃,蕹菜09:00左右开花,09:30左右花完全开放。汝城蕹菜花无论晴天还是阴天都比其他品种开花较晚,即便天气晴朗时,也要11:00左右才处于完全开放状态。蕹菜在开花前一天13:00有少许花药微微裂开,不散粉。在15:00花药全部裂开,有少许湿润花粉暴露在花药裂口处。在17:00有轻微散粉,花粉处于湿润状态。
2.3 花粉形态、花粉活力和柱头可授性测定
蕹菜花粉粒表观呈球形状、帽子状,均布满尖刺。长宽度均匀,长/宽度99.2~117 μm,高度50.1~86.7 μm(图3)。
渔场蕹菜和长沙藤蕹的花粉和柱头显微观察结果如图4所示。采用的碘-碘化钾染色法处理花粉均没有染色,而后采用TTC染色法效果也不理想,只有少量花粉被染色(未在图中显示)。使用联苯胺-甲萘酚染色法和MTT法染色效果明显,染色快且两种方法染色结果一致。前者花粉被染成红色或粉红色,后者花粉被染成深紫色或紫色。花粉活力和柱头可授性检测结果表明,蕹菜在开花前一天至开花当天,花粉活力都特别高,在开花当天17:00花粉活力显著下降。开花当天11:00-13:00柱头表面冒泡最多,可授性最强;而开花后一天柱头出现褐化萎蔫并有破损现象,不具可授性(表5)。
2.4 P/O值和OCI值估算
以渔场蕹菜和长沙藤蕹为观测对象,对花粉量和胚珠数进行显微观察,求得渔场蕹菜花粉量的平均值为5.08×105,胚珠数为4,P/O值为1.27×105。长沙藤蕹花粉量的平均值为3.94×105,胚珠数为4,P/O值为9.9×104。根据Crunden关于繁育系统的判断标准,渔场蕹菜和长沙藤蕹的繁育系统为专性异交型。
根据Dafni的方法对2种蕹菜的杂交指数进行测量。渔场蕹菜花朵直径平均值为6.01 cm,大于6 mm,记为3,长沙藤蕹花朵直径平均值为3.35 cm,大于6 mm,记为3。在花药还未开裂时,通过联苯胺-过氧化氢溶液检测该时期的柱头具有可授性,因此渔场蕹菜和长沙藤蕹都是雌蕊先熟,记为0。渔场蕹菜和长沙藤蕹都为长柱花,柱头和花药不在同一位置,记为1。3者之和所得OCI值为4,由此判定渔场蕹菜和长沙藤蕹的繁育系统为部分自交亲和,异交为主,需要传粉者。
2.5 繁育系统的检验
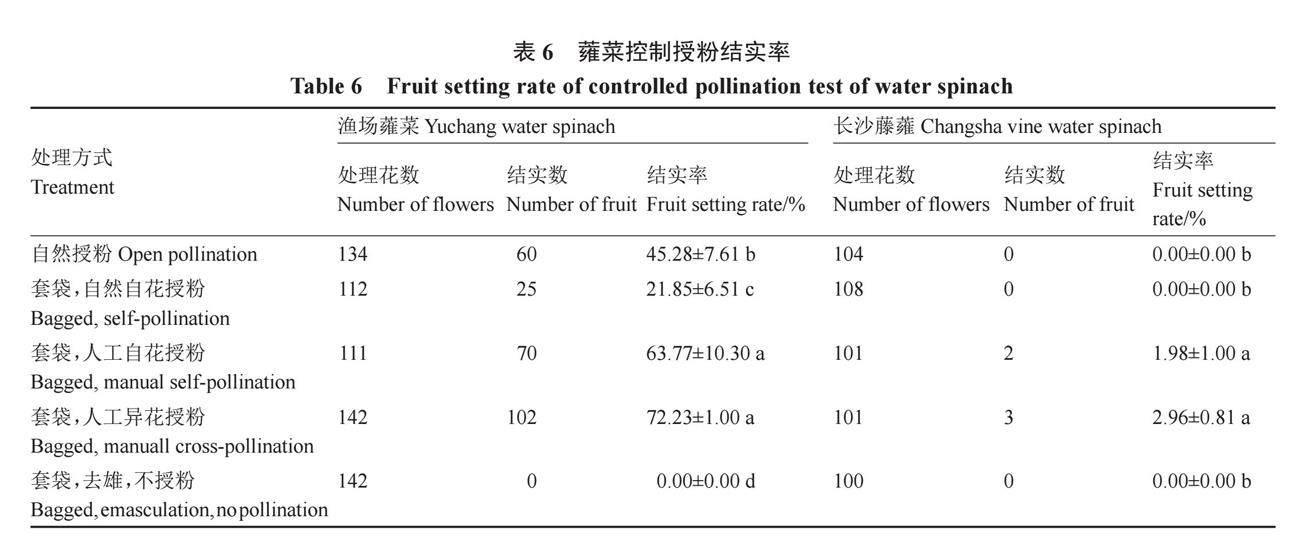
通过自然和人工辅助授粉后,如表6所示,根据控制试验可得出,渔场蕹菜无隔离自然开放授粉能够结实,但结实率只有45.28%,可能是因为还处在高温期且周边缺少植被,访花昆虫少,因此授粉率低。套袋隔离人工异花授粉结实率为72.23%,显著高于套袋隔离自然自花传粉。去雄套袋处理的结实率为0,表明渔场蕹菜不存在无融合生殖现象。人工自花授粉的结实率为63.77%,说明渔场蕹菜不存在杂交不亲和现象。综合判断渔场蕹菜的繁育系统为部分自交亲和,异交需要传粉者。长沙藤蕹通过人工调控光周期诱导开花后,在开放的玻璃温室内进行授粉试验,开放授粉不能结实,且隔离人工自花或异花授粉结实率极低,说明长沙藤蕹丧失了结实能力。
2.6 花期调控和品种间杂交结实率
有性杂交成功的前提是杂交亲本的花期相遇,蕹菜是专性短日照植物,在11 h·d-1光周期处理13 d都能现蕾,蕹菜是随生长持续现蕾开花的植物,花期都能相遇。以4种蕹菜互为父母本进行杂交;以渔场蕹菜和长沙藤蕹的杂交子一代作为母本,渔场蕹菜和长沙藤蕹分别作为父本进行回交,组成的杂交组合授粉、结实情况如表7所示。在籽蕹和藤蕹的杂交组合中,以长沙藤蕹为父本的杂交组合都能够结实,收到杂种种子,汝城蕹菜和长沙藤蕹的杂交结实率达到75.00%,泰国大叶蕹菜与长沙藤蕹杂交结实率达到51.00%,渔场蕹菜与长沙藤蕹杂交结实率达到21.67%,说明籽蕹和藤蕹杂交是亲和的,长沙藤蕹的花粉是可育的且有授粉活性。在回交组合中,渔场蕹菜作为父本的结实率比长沙藤蕹作为父本的结实率高,说明渔场蕹菜和长沙藤蕹的杂交一代与其母本杂交更为亲和。所有以长沙藤蕹为母本的杂交组合的授粉成功率均极低,即使有少量授粉花朵可坐果,但果实在之后的发育过程中会凋亡脱落,不能收获杂交种子,说明长沙藤蕹有可能丧失了结实能力。
3 讨论与结论
3.1 蕹菜的杂交授粉技术
植物花粉活力高低和保持时间长短直接影响到植物传粉和受精的成功率,不同的植物花粉活力和保持时间有巨大差异[15-16],不同检测方法的检测效果也不一致[17],尤其是花粉染色法在不同物种间的染色效果差异较大,找到适合的方法才能准确鉴定花粉活力。何瑞等[18]采用TTC染色法测定唐古特瑞香的花粉活力,各时期花粉均可染色,无法判定花粉活力;而丁仁翔等[19]采用TTC法测定绿榕蒿的花粉活力,却效果显著。笔者采用碘液染色和TTC染色的效果均不佳,联苯胺-甲萘酚染色法和MTT法显色明显,准确且快速,重复的一致性好,适合蕹菜花粉活力的快速鉴定。试验结果表明,蕹菜在开花前一天就具有了活力,但当天09:00—15:00花粉活力较高,且该时间段柱头的可授性好,随后花粉活力和柱头可授性显著降低,说明蕹菜的花粉属于短寿命花粉,因此授粉工作适宜安排在开花当天09:00—13:00。不同品种在自然光周期条件下的现蕾开花期存在明显差异,藤蕹属于专性短日照植物,可以通过短日照诱导使对短日照要求严格的长沙藤蕹提早现蕾开花,与其他花期不同的籽蕹花期相遇,顺利实现有性杂交。为避免发生自交现象,在蕹菜开花前一天的15:00前去雄后套袋,在开花当天上午授粉后捻花闭合花冠,防止外来花粉干扰,保证种子纯度。
3.2 蕹菜的繁育系统和杂交育种体系构建
植物繁育系统具有多样性,自然界很少有绝对自交或杂交的类群,大多是两者兼有的混合交配模式[20]。过度的近交和远交都会导致繁育系统适合度下降,中间程度的杂交即混合交配系统为最适交配方式[21]。本试验结果表明,渔场蕹菜的P/O值和OCI值判定结果有所差异,前者为专性异交,后者为部分自交,以异交为主,可以判定其繁育系统也倾向于异交的混合交配型,而长沙藤蕹的P/O值和OCI值判定结果与渔场蕹菜一致,但在控制授粉试验中表现为自交不亲和。在控制授粉试验中,在室外隔离条件下渔场蕹菜自然结实率超过20%,其原因可能是花药裂开后,虽然花粉颗粒大且黏重,但等长花柱花的花药与花柱紧挨在一起,即使是长花柱花和短花柱花的花药和花柱的距离多数相距0.1~0.2 cm,最远不超过0.5 cm,风吹动植株有可能将花粉风吹到了柱头上完成受精,还可能因花粉量多,花药裂开时的冲击力使部分花粉飘散到柱头上而授粉,同时也观察到有蚂蚁等昆虫从植株基部钻进网袋,到达花朵内部完成了传粉工作。
3.3 长沙藤蕹不结实现象
自然界植物的繁殖方式具有多样性,无论植物是有性繁殖还是无性繁殖,或既能有性繁殖又能无性繁殖,都是植物对环境适应的结果,在某些环境下无性繁殖策略更具有竞争力[22-23]。菊芋(Helianthus tuberosus)遮阴、浇水和施肥结实的研究结果表明了环境能够影响植物在有性繁殖和无性繁殖之间的完全转换[24],有性繁殖与无性繁殖的转换可能与限制性资源的分配有关,但不一定表现出完全的相关性[25]。园艺植物中的很多植物采用无性繁殖方式繁殖,如多数果树,部分蔬菜如芋头[26-27]、黄花菜[28]等都可以正常开花结实或通过人工诱导开花结实,这是人类基于保持品种优良性状选择的结果,无性繁殖可以避免优良性状的分离,但也有少数无性繁殖的园艺植物则完全丧失了结实能力[29]。无性繁殖植物丧失有性生殖能力,其原因有染色体异常[30]、花粉发育异常导致的小孢子败育[31],营养生长与生殖生长的竞争[32]和不利环境条件导致的有性繁殖竞争失败的长期积累[33-34]。长沙藤蕹是蕹菜中食用品质比较优良的品种,对短日照要求严格,在长沙地区因花芽分化期太晚而无法正常现蕾开花,生产上长期采用无性繁殖方式留种。长沙藤蕹在人工短日照诱导下能够正常现蕾开花,但无论是自交、异交还是作为母本进行品种间杂交都结实失败,但花粉可育有活性,作为父本给其他品种授粉有較高的结实率,因此长沙藤蕹结实失败的原因不是杂交不亲和雄性不育,而应该是雌性不育。雌性不育在很多作物中都有发现[35],并对其机制进行了广泛研究。长沙藤蕹雌性不育的原因可能是柱头障碍、受精不良或栽培环境不能满足授粉受精后种胚发育的要求,这种因不利生态因子导致的不能结实转化为遗传雌性不育而被固定下来,其确切的原因有待于进一步研究。
通过设置11 h·d-1短日照,实现了蕹菜品种间的花期相遇;在开花当天09:00—13:00适宜蕹菜授粉;蕹菜繁育系统倾向于异交混合交配型;蕹菜品种间杂交是亲和的,长沙藤蕹的花粉是可育且有授粉活性的,但有可能丧失了结籽能力。
参考文献
[1] 王希强.空心菜优质高产栽培技术[J].现代农业科技,2010(21):128-140.
[2] 朴立喜,刘娟,艾辛,等.长沙藤蕹开花的光周期特性研究[J].中国蔬菜,2018(7):44-47.
[3] 李绪杰,盛雅娟,张彤,等.多花黄精花粉萌发与传粉生物学特性[J].中山大学学报(自然科学版),2021,60(6):110-120.
[4] 张庚,张笑玲,孟义江,等.掌叶半夏开花特性及繁育生物学研究[J].河北农业大学学报,2020,43(2):47-52.
[5] 林鹏,杨虎彪.吊罗山薹草开花生物学特征及繁育系统研究[J].热带作物学报,2021,42(3):689-694.
[6] 叶家桐,毛世忠,胡兴华,等.矮紫金牛的开花生物学与繁育系统研究[J].广西植物,2023,43(2):379-389.
[7] 王洁,杨志玲,杨旭.濒危植物繁育系统研究进展[J].西北农林科技大学学报(自然科学版),2011,39(9):207-213.
[8] CRUDEN R W.Pollen ovule ratios:A conservative indicator of breeding systems in flowering plants[J].Evolution International Journal of Organic Evolution,1977,31(1):32-46.
[9] DAFNI A.Pollination ecology:A practical approach[M].New York:Oxford University Press,1992.
[10] 杨澜,彭强,彭婷,等.影响多肉植物花粉活力及离体萌发率的因素研究[J].种子,2020,39(8):12-16.
[11] 梁露.不同荷花资源的花粉活力比较分析及花粉储藏技术探究[D].郑州:河南农业大学,2022.
[12] 王苗苗,唐灿明.高温胁迫后的棉花花粉活力测定方法[J].江苏农业学报,2010,26(2):236-239.
[13] 张浩锋,乜晓凤,麦晋贤,等.岗梅花部特征、开花物候及繁育方式分析[J/OL].分子植物育种,[2023-09-04]. https://link.cnki.net/urlid/46.1068.s.20230901.1853.020.
[14] 伏秦超,杨浩,唐燕翔,等.肉果秤锤树花粉活力测定及柱头可授性分析[J].分子植物育种,2023,21(22):7518-7523.
[15] 郭红超,严成,魏岩.木地肤的开花动态与花粉活力及柱头可授性研究[J].草业学报,2014,23(4):87-93.
[16] 苏芸芸,王康才,薛启.不同产地藿香花粉活力与柱头可授性研究[J].草业学报,2016,25(9):189-196.
[17] 贾娟霞,韩雯毓,乔文杰,等.蓖麻花粉活力检测方法及其特性研究[J].内蒙古民族大学学报(自然科学版),2023,38(4):335-339.
[18] 何瑞,闫芳,祝睿彦,等.唐古特瑞香开花物候及繁育系统研究[J].草地学报,2024,32(1)188-197.
[19] 丁仁翔,刘浩,朱科燃,等.夹金山3种同域分布绿绒蒿的传粉生态学研究[J].草业学报,2024,33(1):207-216.
[20] 陈之光,徐言,葛红,等.萼脊蝴蝶兰开花特性与繁育系统研究[J].浙江农林大学学报,2023,40(6):1197-1204.
[21] 何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151-163.
[22] 肖月娥,田旗,周翔宇,等.玉蝉花繁殖生态学研究[J].云南植物研究,2010,32(2):93-102.
[23] 张玉芬,张大勇.克隆植物的无性与有性繁殖对策[J].植物生态学报,2006,30(1):174-183.
[24] 张玉凤,陈荣健,马辉,等.不同环境因素对两种结实率菊芋繁殖策略的影响[J].兰州大学学报(自然科学版),2018,54(5):705-710.
[25] 王灵艳.湿地植物燕子花(Iris laevigata)的繁殖对策研究[D].长春:东北师范大学,2020.
[26] 黄新芳,柯卫东,孙亚林,等.芋种质资源开花结实状况调查[J].植物遗传资源学报,2011,12(5):811-815.
[27] 郭巨先,尹艳,唐康,等.芋种质资源研究进展与展望[J].广东农业科学,2021,48(9):81-90.
[28] 李森,刘娟,公菲菲,等.黄花菜‘大同黄花开花特性与繁育系统研究[J].园艺学报,2021,48(8):1541-1551.
[29] 丁巨波,傅焕延.大蒜不孕性的研究 Ⅰ.雄配子體发生期内的养分供应与小孢子败育[J].山东农学院学报,1963(8):27-33.
[30] 刘冰江,张海燕,孔素萍,等.大蒜有性生殖研究进展[J].中国蔬菜,2008(8):41-44.
[31] 周群.湖北麦冬资源品质与不育机理研究[D].武汉:华中科技大学,2009.
[32] 徐培文,杨崇良,崔德才,等.大蒜有性生殖技术研究初报[J].园艺学报,2005,32(3):503-506.
[33] 麻全林.不同生态环境条件下沙生植物的结实规律研究[J].种子科技,2024,42(5):18-22.
[34] 欧明烛.极狭域特有植物辐花苣苔濒危原因及离体培育技术研究[D].贵阳:贵州大学,2023.
[35] 胡青,高述民,李凤兰.植物雌性不育的研究进展[J].北京林业大学学报,2004,26(1):87-91.
猜你喜欢
遗传(2023年9期)2023-09-25
农业科技通讯(2023年1期)2023-02-12
河北果树(2022年1期)2022-02-16
家庭医药·快乐养生(2019年4期)2019-04-23
森林工程(2018年4期)2018-08-04
少儿科学周刊·少年版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2017年7期)2017-09-29
长江蔬菜(2016年10期)2016-12-01
长江蔬菜(2016年10期)2016-12-01
恋爱婚姻家庭·养生版(2016年8期)2016-08-23
