
紫黑稻6号×糯稻89-1 F?主要性状遗传变异及相关性分析
2024-05-22 16:42:06蒋菲黄倩何珊梁慧苑宋子慧赵正武
江苏农业科学 2024年7期
蒋菲 黄倩 何珊 梁慧苑 宋子慧 赵正武

蒋 菲,黄 倩,何 珊,等. 紫黑稻6号×糯稻89-1 F2主要性状遗传变异及相关性分析[J]. 江苏农业科学,2024,52(7):48-55.
doi:10.15889/j.issn.1002-1302.2024.07.007
(重庆师范大学生命科学学院/重庆市特色作物资源工程技术研究中心,重庆401331)
摘要:为揭示彩叶水稻与绿叶水稻杂交后代农艺性状、品质性状以及农艺性状与品质性状之间的遗传变异与相互关系,以彩叶水稻紫黑稻6号与绿叶水稻糯稻89-1杂交得到的F2群体的160株单株为研究材料,采用相关分析、主成分分析及典型相关性分析等方法分析F2群体的9个农艺性状与10个品质性状之间的遗传变异及相关性。结果显示,F2群体农艺性状变异系数范围为11.08%~70.43%,品质性状变异系数范围为3.49%~75.22%,主要农艺和品质性状组内各自呈现出不同程度的正向或负向相关关系,千粒重、株高和穗长呈极显著正相关,与着粒密度呈极显著负相关;结实率与有效穗数、每穗实粒数呈极显著正相关,与每穗空粒数呈极显著负相关;精米率与整精米率、糙米长、糙米宽极显著正相关,与垩白粒率极显著负相关;整精米率与糙米长极显著正相关。主要农艺性状与品质性状组间相关系数绝对值最大为0.157,典型相关分析结果表明农艺性状与品质性状2组变量间相关性强度不高。以上结果表明,增加水稻的产量需在适当提高水稻的有效穗数、每穗实粒数以及增加水稻穗长基础上,再适当降低着粒密度;品种选育过程中,每穗实粒数与外观品质是首要考虑的因素,同时应综合考虑多个性状的平衡发展。
关键词:水稻;紫黑稻6号;糯稻89-1;遗传变异;相关性分析;主成分分析
中图分类号:S511.032 文献标志码:A 文章编号:1002-1302(2024)07-0048-08
水稻(Oryza sativa L.)是三大粮食作物之一,同时也是我国主要的粮食作物,我国50%以上的人口以稻米为口粮[1]。近年来,随着经济文化的发展,人们在追求物质生活的同时更加重视精神生活上的需求,水稻除了传统意义上的饱腹功能外,还被赋予了文化传承和观赏创新等新功能。彩色水稻是组织或器官呈现常规绿色以外的特殊色泽的水稻,是兼具观赏性与营养价值的农作物[2]。大力推进彩色水稻品种的培育与遗传改良,运用技术手段培育兼具多种色彩的水稻作物,彩色稻田与艺术设计结合,与现代乡村文化结合形成了极具特色的稻田画[2-3]。此外,彩色水稻因其色素含量在叶片和颖壳中的不同,在杂交育种中可以用于特定的表型标记辅助除杂,还可以作为特殊的种质资源做叶色相關性状基因的定位与克隆、色素的合成代谢以及光合作用等机制的理论研究[4]。
在水稻发展过程中还有许多问题亟待解决,首先是优质品种的指标优而不全,充分挖掘现有种质资源性状的遗传多样性,有助于减少亲本组配的试错,提高育种成效,为水稻产量、品质提升改良提供选择依据[5]。前人对于水稻的主要性状指标进行了一定的研究,但这些研究大多集中在普通绿色水稻方面。相比之下,现有的关于彩色水稻及其杂交后代相关性状的研究较少,且主要停留在简单的生产应用层面,对其遗传基础和理论研究较少。因此,本研究以紫黑稻6号和绿色水稻糯稻89-1杂交F2群体160个单株为材料,对其农艺性状、品质性状以及农艺性状与品质性状之间的遗传变异及相关性进行了研究,以期为彩色水稻品种选育和利用优异品种资源进行遗传育种提供一定的理论参考。
1 材料与方法
1.1 试验材料
以糯稻89-1为母本、紫黑稻6号为父本杂交获得F2群体,共160个单株为材料。糯稻89-1是糯稻,直链淀粉含量为1.50%,是一个耐冷水稻品种,能够通过腋芽抵御低温休眠越冬,其芽期和幼苗期的耐冷性均达到了1级耐冷标准[6-8]。紫黑稻6号是重庆师范大学选育的彩叶稻品种,2022年通过重庆市农作物品种审定委员会审定(审定编号:渝审稻20220027),叶色为紫黑色,从分蘖盛期至成熟期叶色一致性好,有较强的观赏性。
[BT2+*5]1.2 试验方法
试验于2019年春季在重庆师范大学配组糯稻89-1/紫黑稻6号杂交组合,同年冬季在海南省陵水黎族自治县种植F1,2020年春季在重庆市璧山区种植F2群体,共160个单株。株行距为16.67 cm×26.67 cm,有机肥与化肥配合施用,重底早追,施纯氮120~150 kg/hm2,N、P、K肥施用质量比为6 ∶3 ∶1。F2成熟后进行室内考种。考察性状有株高、穗长、千粒重、结实率、着粒密度、每穗实粒数、每穗空粒数、每穗总粒数、有效穗数。记载标准主要参照文献[9]。根据GB/T 17891—2017《优质稻谷》测定亲本和F2 糙米率、精米率、整精米率、糙米长、糙米宽、糙米长宽比、垩白粒率、垩白度、胶稠度、直链淀粉含量等品质性状。
1.3 数据统计与分析
用Microsoft Excel 2010对农艺性状与品质性状等数据进行整理分析;用IBM SPSS 26.0进行方差分析、相关性分析等;用OriginPro 2022作图。
2 结果与分析
2.1 F2群体主要农艺性状遗传相关分析
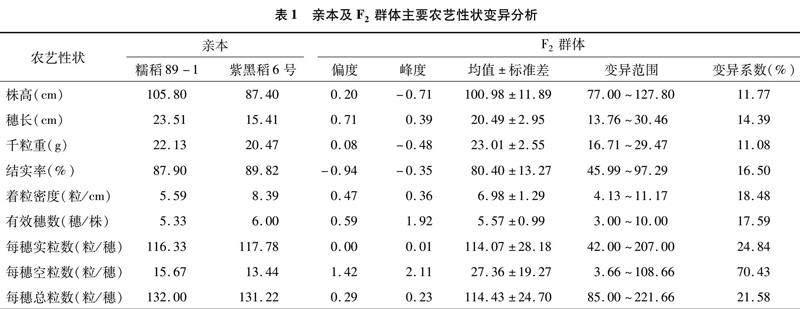
2.1.1 F2群体主要农艺性状遗传变异分析 由表1可知,F2群体9个农艺性状的变异程度各有不同,变异系数的范围为11.08%~70.43%,由大到小依次为每穗空粒数>每穗实粒数>每穗总粒数>着粒密度>有效穗数>结实率>穗长>株高>千粒重,其中每穗空粒数变异系数最大,达到70.43%;千粒重的变异系数最小,为11.08%,穗长、株高和千粒重的变异性较小,变异系数均在15%以下。F2群体9个农艺性状中,除了每穗空粒数的偏度和峰度、有效穗数的峰度之外,其他性状偏度和峰度绝对值都小于1。F2群体中有4个性状表型值的均值介于亲本之间,即株高、穗长、着粒密度和有效穗数;结实率、每穗实粒数和每穗总粒数的均值较亲本低;千粒重和每穗空粒数的均值高于双亲,表现出正向超亲优势。
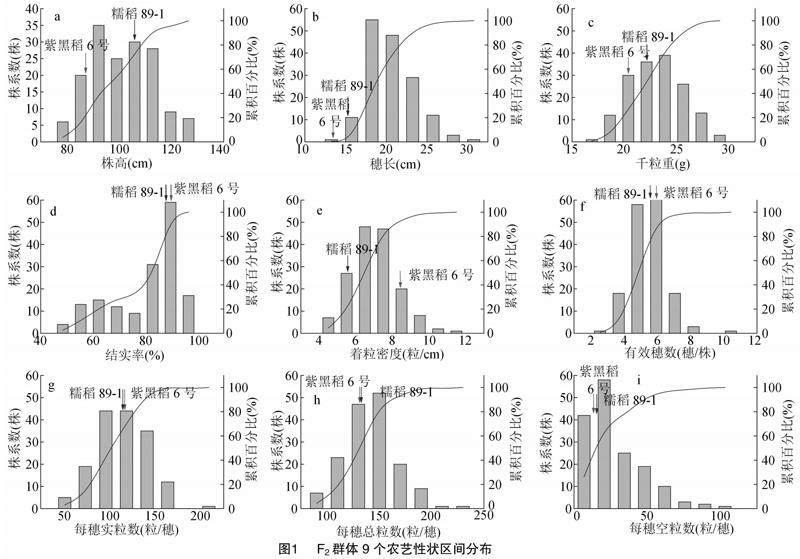
2.1.2 F2群体主要农艺性状分布表现 F2群体主要农艺性状分布区间表现见图1,除了每穗空粒数之外,其他8个性状的区间分布近似正态分布。可以看出,株高(图1-a)和穗长(图1-b)均呈连续变异,株高的偏度和峰度分别为0.20和-0.71,穗长的偏度和峰度分别为0.71和0.39。株高主要分布在88~116 cm之间,位于此范围内的第3~6个分布区间分布株系数分别为35、25、30、28个,该株高范围占群体的73.75%。穗长主要集中在17~22 cm 之间,其中有103个株系的穗长处于该范围内,占群体的64.38%。每穗实粒数、结实率、有效穗数、千粒重是水稻主要的产量构成因素。该供试群体千粒重(图1-c)大于20 g的株系占到了89.37%,并且有38个株系的千粒重超过了25 g;结实率(图1-d)主要集中在86.25%~93.25%之间,超过90%[JP3]的有46个株系; 有效穗数(图1-f)为4~6穗/株[JP]的株系偏多,有22个株系超过6穗/株;每穗实粒数(图1-g)大部分分布于85~130粒/穗区间内,超过110粒/穗的有86个株系,且有48个株系每穗实粒数超过了130粒。
2.1.3 F2主要农艺性状相关分析 对F2群体9个农艺性状的相关分析结果(表2)表明,不同性状之间存在不同程度的相关性。其中结实率和每穗空粒数的相关系数最大,数值为-0.936,具有极显著负相关性。千粒重、株高和穗长呈极显著正相关,与着粒密度呈极显著负相关,与其他性状无显著相关性。表明千粒重同时受株高、穗长、着粒密度等性状共同影响,单株千粒重值大的材料往往株高、穗长值较大,着粒密度较小。结实率与有效穗数、每穗实粒数呈极显著正相关,與穗长呈显著正相关,而与每穗空粒数呈极显著负相关,多穗长穗株系结实率较高,每穗空粒数较少,因此,在选育品种时,不能一味关注每穗粒数,可同时考虑穗长和单株穗数再综合选择。每穗实粒数与穗长、结实率、着粒密度、有效穗数以及每穗总粒数呈极显著正相关,与株高呈显著正相关,而与每穗空粒数呈极显著负相关。此外,株高与穗长,穗长与每穗总粒数,着粒密度与每穗空粒数、每穗总粒数,这些性状间相互关系表现为极显著正相关,而着粒密度与株高、穗长、千粒重,每穗空粒数与有效穗数均呈极显著负相关。9个农艺性状间存在不同程度的相关性或制约关系,增加水稻的产量需在适当提高水稻的有效穗数、每穗实粒数以及增长水稻穗长的基础上,再适当降低水稻着粒密度。
2.1.4 F2群体主要农艺性状主成分分析及聚类分析 对F2群体160份水稻株系的9个农艺性状进行主成分分析,得到9个性状遗传相关矩阵的特征根和对应的特征向量值(表3)。前4个因子的累计贡献率达到86.499%,代表了F2群体86.499%的性状信息,可用这4个因子代表9个农艺性状对160份材料进行分析和评价,概括农艺性状的绝大部分信息。第1主成分的特征值为2.802,贡献率为31.131%,每穗实粒数、结实率的特征向量值较大,并呈明显正相关关系,第1主成分主要反映了水稻产量特性,可称为产量性状因子。第2主成分的特征值为2.288,贡献率为25.426%,主要指标为着粒密度,特征向量值为0.946,称为着粒密度因子。第3主成分的特征值为1.694,贡献率为18.819%,以穗长为主要指标,称为穗长因子。第4主成分贡献率为11.122%,主要指标为有效穗数,可称为有效穗数因子,向量值为0.845。
以结实率大于等于90%为选择指标,筛选出46个株系进行聚类分析(图2),在遗传距离为50处可将结实率大于等于90%的株系分为4类,其中第Ⅰ类涵盖株系数最多,占比89.13%,除每穗空粒数,第Ⅰ类中大部分株系的性状高于供试群体性状的均值,第Ⅱ类占比652%,第Ⅱ类株系中结实率与有效穗数多高于群体均值,第Ⅲ类与第Ⅳ类占比均为2.17%,表现为所有性状均高于均值。分析表明,供试群体中结实率大于等于90%的株系大部分表现为各个性状均衡发展,且优于群体均值,表明在田间选育优良株系时,应考虑性状的综合表现,不能单一靠某一性状判断选择。
2.2 F2群体主要品质性状遗传相关分析
2.2.1 F2群体主要品质性状遗传变异分析
由表4可知,F2群体主要品质性状变异系数范围为349%~75.22%,大小顺序依次为垩白度>直链淀粉含量>垩白粒率>整精米率>精米率>糙米长宽比>糙米宽>糙米长>胶稠度>糙米率。其中胶稠度、糙米率、糙米长的变异较小,变异系数均在10%以下。供试群体的品质性状中,除胶稠度的均值大于亲本,其他性状均值均小于或介于亲本之间。此外,除精米率、糙米长、糙米长宽比、垩白度的偏度及精米率、垩白粒率的峰度的绝对值接近于1以外,其他性状的偏度和峰度的绝对值均远离1,供试群体的品质性状分布大多不呈正态分布,分布的偏移程度严重。
2.2.2 F2群体主要品质性状相关分析
由表5可知,F2群体的10个品质性状有16对相关系数达到极显著差异水平(P<0.01),7对相关系数达到显著差异水平(P<0.05)。其中糙米率与精米率、糙米长极显著正相关,与整精米率、糙米宽显著正相关;精米率与整精米率、糙米长、糙米宽极显著正相关,与垩白粒率极显著负相关,与糙米长宽比、垩白度显著相关;整精米率与糙米长极显著正相关;糙米长与糙米宽、糙米长宽比、直链淀粉含量极显著正相关,与垩白粒率、垩白度极显著负相关;糙米宽与糙米长宽比极显著负相关,与垩白粒率显著负相关;糙米长宽比与胶稠度和直链淀粉含量显著正相关;垩白粒率与垩白度极显著正相关,与直链淀粉含量极显著负相关,垩白度与直链淀粉含量极显著负相关。
2.2.3 F2群体主要品质性状主成分分析 F2群体的10个品质性状主成分分析结果见表6,前3个因子的贡献率分别为28.894%、18.695%、16.649%,累计贡献率为64.238%,因此选择前3个主成分对10个品质性状的信息作综合评价分析。其中,第1主成分的垩白度、垩白粒率有较大的负系数,而糙米长有较大的正系数,表明该主成分反映垩白度、垩白粒率和糙米长的对比,主要表现为外观品质特征;第2主成分中糙米率和精米率有较大正系数,主要表现为碾磨品质特征;第3主成分的糙米长宽比有较大正系数,表明该成分是糙米长宽比的综合反映,第3因子同样为外观品质特征。
2.3 F2群体主要性状间的相关性分析
由图3可知,主要性状之间都呈现出不同程度的正相关或负相关关系,但二者相关系数绝对值均不超过0.2,系数绝对值最大只达到0.152,说明F2群体主要农艺性状和品质性状的相关性不高。
为进一步研究F2群体主要农艺性状和品质性状的定量关系,将研究数据分为农艺性状组(U)和品质性状组(V),对其进行典型相关分析,得出9对典型变量,结果见表7。其中前4对典型变量的相关系数大于0.2,存在线性相关关系,但2组变量组间的相关性强度不大,与简单相关分析结果相似。第1对典型相关变量的相关性系数最大,为0.370。后5对典型相关系数小于0.2,相关性较弱,可以忽略。第1对典型变量的标准化线性组合如下:U1=-0.554x1+0.321x2+0.212x3+0.054x4-0.602x5+0.139x6-2.028x7-0.905x8+2.137x9;V1=-0167y1+0.295y2-0.099y3+3.779y4-2.750y5-4.186y6+0.719y7-0.949y8-0.093y9-0.236y10(x1~x9分别指株高、穗长、千粒重、结实率、着粒密度、有效穗数、每穗实粒数、每穗空粒数和每穗总粒数;y1~y10分别指糙米率、精米率、整精米率、糙米长、糙米宽、糙米长宽比、垩白粒率、垩白度、胶稠度和直链淀粉含量)。
根据以上公式可知,在第1对典型变量中分别以每穗总粒数和糙米长宽比的系数较大,表明第1对典型变量主要由每穗总粒数和糙米长宽比决定,农艺性状各指标可以被第1典型变量解释的比例分别为 9.6%、11.7%;品質性状各指标可以被解释的比例分别为9.1%、10.5%。
3 讨论与结论
杂交水稻F2代是性状分离最大的时期,也是表型最为丰富的时期,陈越等在对60份杂交水稻资源的分析中筛选到株高、千粒重等6个水稻资源综合评价的关键指标[10];石春海等提出控制植株农艺性状的遗传效应与影响稻米品质性状的遗传效应间存在较强的相关性,有效穗数与糙米率等成对性状间的显性效应相关性,有助于同时改良杂交稻组合的这些性状[11];冯建成研究表明,水稻株高增加对产量提高也起到间接的促进作用[12];朱业宝等在以1 040份福建省水稻地方品种为材料的分析中发现质量性状遗传多样性指数均小于数量性状的遗传多样性指数[13]。
育种工作者为了满足生产需求而创造遗传变异,遗传参数的变异系数可以在一定程度上反映性状的多样性。本研究以糯稻89-1与紫黑稻6号杂交获得的F2群体进行性状分析发现,除了每穗空粒数之外,其他的8个性状数值分布情况接近于正态分布且呈连续性变异,这些性状是由微效多基因控制的数量遗传性状,可进一步进行QTL定位分析[14]。品质性状大多不呈正态分布,分布的偏移程度严重。9个农艺性状变异系数的变化范围为11.08%~70.43%,其中每穗空粒数变异系数最大,千粒重最小,F2群体主要品质性状变异系数范围为3.49%~75.22%,变异系数小的性状主要受到自身遗传效应的影响,变异潜力不大;而变异系数较大的性状,易受外界环境因素的影响,性状变异类型就较为丰富,为选育新品种以及改良品种性状提供了良好的遗传基础[15]。F2代的主要性状相关性分析表明,千粒重、株高和穗长表现为极显著正相关,而与着粒密度呈极显著负相关;结实率与有效穗数、每穗实粒数呈极显著正相关,而与每穗空粒数呈极显著负相关,研究结果与王新俊等的研究结论[16-17]略有不同,可能是因为水稻品种、栽培地理环境以及气候等的不同而造成的。何秀英等提出对杂交后代进行材料改良和品种选育时,需要同时兼顾各性状间的相互影响,选择各性状合理发展型水稻品种是获得高产的有效途径之一[18]。通过本研究分析结果发现,由于不同农艺性状对产量的影响不同,增加水稻的产量需在适当提高水稻的有效穗数、每穗实粒数以及增长水稻的穗长的基础上,再适当降低水稻的着粒密度。
采用主成分分析法,通过统计分析将数据简化成比较少的综合指标对水稻性状进行综合分析,可以减少分析误差[19-21]。农业性状的主成分分析中第1因子为产量因子,其中每穗实粒数的特征向量值最大,其次是结实率,最后是每穗总粒数,说明每穗实粒数对水稻产量有极大的影响;品质性状的第1因子是外观品质特征,因此,要达到选育优质水稻株系的目的,每穗实粒数与外观品质是首要考虑的因素,这与韩锋等提出对加工品质、外观品质进行综合选择是优质水稻品种选育的主要途径的结果[22]一致。
为研究2组变量间的相关关系,对主要农艺和品质性状进行相关分析和典型相关分析,F2群体主要农艺和品质性状之间呈现出不同程度的正向或负向相关关系,但简单相关系数绝对值均不超过0.2,典型相关变量系数最大,为0.37,相关性不高。这与陈慢慢等在对400个重组自交系群体性状分析中测得的直链淀粉含量与穗长呈极显著负相关的结果[23]有所不同,可能是因为供试材料本身存在差异。
育种进程中,农艺性状和品质性状是种质资源鉴定评价的重要依据。遗传分离群体后代极易产生双向超亲现象,成为具有良好综合性状的个体,是育种过程中筛选优质和目标种质的有效资源[24-26]。在水稻品种选育和改良的过程中,要选择变异系数比较大的性状进行,变异类型丰富,遗传基础良好。此外,水稻各个性状之间都有非常密切的关系,在实际的育种过程中,要综合考虑各性状,协调选择。本研究对F2群体多个主要性状的相关分析表明,不同性状之间存在不同程度的相关性,但部分表型性状易受到遗传或环境因素的影响,因此本研究中分析的主要农艺性状与品质性状的相关关系不能直接说明他们之间的相关关系,且选育品种不能单纯追求某一性状,需考虑多个性状互作,才能选育出综合性状较好的水稻株系。
本研究表明,增加水稻的产量需在适当提高水稻的有效穗数、每穗实粒数以及增长水稻穗长的基础上,再适当降低水稻的着粒密度;品种选育过程中,每穗实粒数与外观品质是首要考虑的因素,同时应注重性状间的互作,考虑多个性状的平衡发展再综合选择。
参考文献:
[1]张海淼,李 洋,刘海峰,等. 水稻重要农艺性状调控基因及其育种利用研究进展[J]. 生物技术通报,2020,36(12):155-169.
[2]黄小芬. 彩色水稻及其在乡村旅游风景文化塑造中的应用[J]. 植物遗传资源学报,2022,23(6):1867-1868.
[3]宋文健,梅 忠,李 玉,等. 彩色水稻研究与利用现状[J]. 中国水稻科学,2020,34(3):191-204.
[4]徐智慧,高作利,任邵琦,等. 彩色水稻花色苷的研究进展[J]. 分子植物育种,2022,20(18):6226-6235.
[5]曾 波,钟育海,郭利磊,等. 我国优质水稻品种发展现状与展望[J]. 种子,2019,38(8):53-56.
[6]赵正武,李仕贵,雷树凡. 一份特异多年生水稻种质主要农艺性状的遗传变异[J]. 中国水稻科学,2006,20(5):481-486.
[7]Deng X S,Gan L,Liu Y,et al. Locating QTLs controlling overwintering seedling rate in perennial glutinous rice 89-1 (Oryza sativa L.)[J]. Genes & Genomics,2018,40(12):1351-1361.
[8]Zhao Z W,Luo A C,Le T,et al. Identification of quantitative trait locus for overwintering germinability in rice (Oryza sativa L.)[J]. Journal of Integrative Agriculture,2012,11(11):1767-1774.
[9]韩龙植,魏兴华. 水稻种质资源描述规范和数据标准[M]. 北京:中国农业出版社,2006.
[10]陈 越,张敦宇,丁明亮,等. 多个省份水稻资源的表型多样性与优异资源的筛选[J]. 浙江农业学报,2019,31(11):1779-1789.
[11]石春海,朱 军. 水稻植株农艺性状与稻米碾磨品质的遗传相关性分析[J]. 浙江农业大学学报,1997,23(3):331-337.
[12]冯建成. 高产水稻新品种漳丰8号的选育[J]. 江西农业学报,2007,19(8):35-36,43.
[13]朱业宝,陈立喆,张 丹,等. 福建省水稻地方品种表型性状遗传多样性分析[J]. 福建农业学报,2021,36(10):1119-1125.
[14]陈 娇,甘 露,罗 兰,等. 黏糯杂交后代主要农艺性状遗传变异及相关性和主成分分析[J]. 安徽农业科学,2019,47(6):33-35,38.
[15]胡立勇,丁艳锋. 作物栽培学[M]. 北京:高等教育出版社,2008.
[16]王新俊,曾晓芳,赵德刚. 贵州地方水稻品种品质性状及其相关分析[J]. 种子,2013,32(9):70-74.
[17]任芹勇,樊巧利,李 涛,等. 65份谷子品种农艺性状聚类和相关性[J]. 分子植物育种,2017,15(12):5178-5188.
[18]何秀英,陈钊明,廖耀平,等. 水稻收獲指数遗传及其与主要农艺性状的相关研究[J]. 作物学报,2006,32(6):911-916.
[19]苏理云,陈彩霞,高红霞. SPSS 19统计分析基础与案例应用教程[M]. 北京:北京希望电子出版社,2012:150-247.
[20]游书梅,曹应江,郑家奎,等. 73份亚洲水稻恢复系农艺性状的主成分与聚类分析[J]. 植物遗传资源学报,2015,16(2):250-256.
[21]田 颖,于雪然,杜怀东,等. 水稻粒形基因的遗传研究进展[J]. 江苏农业科学,2022,50(21):16-26.
[22]韩 锋,徐 娟. 2011—2015年辽宁水稻育成品种品质性状分析[J]. 北方水稻,2017,47(5):12-15.
[23]陈慢慢,田 翔,陈 凌,等. 谷子重组自交系主要农艺性状和品质性状相关分析[J]. 植物遗传资源学报,2023,24(1):172-180.
[24]王 忠,顾蕴洁,陈 刚,等. 稻米的品质和影响因素[J]. 分子植物育种,2003,1(2):231-241.
[25]莫惠栋. 我国稻米品质的改良[J]. 中国农业科学,1993,26(4):8-14.
[26]陈建民,李 刚,顾建强,等. 水稻超高产育种研究现状与实践[J]. 福建稻麦科技,2009,27(4):48-53.
基金项目:国家自然科学基金(编号:31670326);重庆市技术创新与应用发展重点专项(编号:CSTB2022TIAD-KPX0015、CSTB2022TIAD-KPX0018)。
作者简介:蒋 菲(2000—),女,四川遂宁人,硕士研究生,主要从事水稻生理学研究。E-mail:2068331201@qq.com。
通信作者:赵正武,博士,研究员,主要从事水稻遗传育种研究。E-mail:zhaozhengwu513@sina.com。
猜你喜欢
江苏农业科学(2024年14期)2024-08-23 00:00:00
安徽农学通报(2024年12期)2024-07-13 20:26:34
保健与生活(2023年9期)2023-05-19 21:07:36
麦类作物学报(2022年7期)2022-08-09 08:04:46
江苏农业科学(2020年22期)2020-03-03 14:37:13
现代农业科技(2018年14期)2018-10-31 10:50:20
江西农业学报(2016年5期)2016-06-21 05:45:20
江苏农业科学(2016年4期)2016-06-14 01:41:21
麦类作物学报(2016年4期)2016-05-27 01:28:18
食品工业科技(2014年13期)2014-03-11 18:16:55
