
基于生物信息学方法探究白癜风铁死亡相关发病机制和潜在治疗药物筛选
2024-05-14 00:50刘祥冉李治建阿卜杜热伊木阿力木江魏文婧霍仕霞
现代药物与临床 2024年4期
刘祥冉,李治建, ,阿卜杜热伊木·阿力木江,魏文婧,霍仕霞
1.新疆医科大学 药学院,新疆 乌鲁木齐 830011
2.新疆维吾尔自治区维吾尔医医院,新疆 乌鲁木齐 830049
3.新疆中药医院制剂循证与转化重点实验室,新疆 乌鲁木齐 830049
白癜风是一种自身免疫性皮肤色素脱失性疾病,主要临床表现为皮肤出现局限性白色斑块。据统计,白癜风患病率占全球人口的0.5%~2%[1]。大多数研究认为白癜风发病主要与自身免疫[2]、氧化应激[3]、局部炎症及角质形成细胞功能异常[4]有关。尽管目前的研究结果对白癜风的发病机制提供了部分见解,但确切机制仍需进一步证实。
铁死亡的主要机制是铁和脂质活性氧(L-ROS)的积累以及一组特定基因的参与导致的铁相关性程序性细胞死亡[5]。谷胱甘肽过氧化物酶4(GPX4)是铁死亡的关键因子[6]。GPX4 活性下降,氧化脂质产生大量的活性氧(ROS)。这种氧自由基可增强细胞的氧化损伤,诱导细胞死亡[7]。氧化应激诱导的免疫反应是白癜风发病的触发因素,而ROS 过度积聚是激活氧化应激的主要原因[8]。氧化应激中细胞内质网应激可触发一种称为未折叠蛋白反应(UPR)的细胞反应,UPR 可引起ROS 诱导的CXC 趋化因子配体16(CXCL16)活性增加,CXCL16 是白癜风中细胞毒性T 细胞迁移所必需的趋化因子[9]。此外,ROS 可直接诱导角质形成细胞分泌三磷酸腺苷也可引发半胱氨酸蛋白酶-3(Caspase-3)介导的黑素细胞死亡,也可诱导黑素细胞附近的角质形成细胞产生CXCL9/10,从而招募CD8+T 细胞在局部组织浸润产生细胞杀伤作用[10]。故铁死亡、ROS 生成和氧化应激可能与白癜风患者体内免疫系统功能有密切联系。在皮肤组织中,多种原因(包括铁死亡)导致ROS 的过量产生所引起的氧化应激,能够激活γ 干扰素(IFN-γ)-CXCL9/CXCL10-CXCR3 轴、CXCL16-CXCR6 轴和高迁移率族蛋白 B1(HMGB1)-核因子-κB(NF-κB)p65/细胞外调节蛋白激酶(ERK)-CXCL8 轴导致下游的免疫细胞功能紊乱,使皮肤中免疫稳态失衡,导致白癜风的发病[9,11-13]。所以推测通过铁死亡途径治疗白癜风的根本机制是使白癜风患者免疫系统趋向稳态,调节功能异常的免疫细胞发挥正常的免疫功能。
雷公藤是卫矛科雷公藤属植物,具有抗炎、抗肿瘤、免疫调节等作用[14]。雷公藤作为皮肤科常用药,临床可用于治疗慢性荨麻疹、皮肌炎和银屑病等[15]。雷公藤甲素是雷公藤中主要活性成分之一,可抑制多种炎性介质和淋巴细胞增殖,具有很好的抗炎和抑制免疫活性作用[16]。相关研究报道雷公藤甲素对角质形成细胞有明显的抑制作用,并且研究进一步证实雷公藤甲素能有效抑制角质形成细胞中IFN-γ 响应性Janus 激酶(JAK)/信号传导及转录激活蛋白(STAT)信号通路的表达[17]。白癜风相关研究也发现IFN-γ 刺激角质形成细胞,细胞内JAK/STAT 信号激活使细胞分泌大量的CXCL9/10,从而招募黑素细胞特异性CD8+T 细胞聚集产生杀伤黑素细胞的作用,在此过程中角质形成细胞具有极其关键的作用,所以抑制角质形成细胞可能是雷公藤甲素治疗白癜风的潜在机制[18]。本研究利用生物信息学[19]方法寻找与白癜风发生发展相关的高质量铁死亡基因及相关的免疫信号机制,并通过高通量筛选获得雷公藤甲素作为潜在治疗药物,旨在为白癜风的发病机制研究及治疗策略提供参考依据。
1 材料和方法
1.1 数据来源
使用R 语言GEOquery 包[20]从高通量基因表达(https://www.ncbi.nlm.nih.gov/geo/)数据库中获取原始数据集GSE53146(GPL14951)作为分析的实验集(包括5 位白癜风患者、5 位健康志愿者);GSE75819(GPL6884)数据集作为关键基因外部验证集(包括15 位白癜风患者、15 位健康志愿者);GSE203262(GPL20301)数据集作为单细胞评估分析集(包括6 位白癜风患者、6 位健康志愿者)。此外,本工作中获取的铁死亡基因(FRGs)是基于FerrDB 数据库中Driver、Suppressor、Marker 模块获得铁死亡相关基因。
1.2 差异表达基因筛选
获取数据集中原始表达矩阵后,通过Normalize Between Arrays 校正除去批次效应,并通过R 软件limma 包[21]分析白癜风患者皮肤及健康对照皮肤之间的差异表达基因(DEGs),将|log2FC|>1,P<0.05作为具有显著性差异的标准。
1.3 筛选白癜风铁死亡关键基因
在R 语言中使用ggvenn 包获得DEGs 与FRGs交集基因作为白癜风铁死亡差异表达基因(FRDEGs)。利用glmnet 包[22],采用最小绝对收缩选择算子(LASSO)来减少数据维度。同时获取LASSO算法预测的基因[23]。利用kernlab 包[24]构建支持向量机递归特征消除(SVM-RFE)模型,并使用平均误判率与10 倍交叉验证进行比较[25],筛选模型预测基因。2 种算法所得基因取交集即白癜风铁死亡关键基因(也是通过铁死亡途径治疗白癜风的关键靶点)。使用pROC 包绘制受试者工作特征(ROC)曲线,并确定曲线下面积(AUC)值,以评估本研究选择的标记基因是否具有诊断价值。另外,本研究还利用circlize 包[26]用来分析关键基因之间的相互作用关系。
1.4 单细胞聚类分析及关键基因验证
此部分基于GSE203262 数据集进行。该数据集涵盖6 个白癜风和6 个正常样本。在本研究中使用Seurat R 包[27]进行质量控制。细胞剔除的标准为(1)RNA 计数<50,(2)线粒体基因表达率<5%,利用Seurat 中的Normalize Data 函数对数据进行规范化。在后期分析中,选择了15 个最显著的主成分和2 000 个最显著的可变基因。使用Seurat 的Find Clusters 函数(分辨率=0.5)检测细胞簇,并采用2Dt 分布随机嵌入(tSNE)显示[28]。本研究采用Single R 包,将不同簇中的细胞与带注释的参考数据集进行比较[29]。根据识别的细胞标记和比较结果完成细胞类型的聚类标注。利用HPA 数据库进一步验证关键基因在正常皮肤组织细胞群中的表达,表达水平显示为4 种类型:未检测到、低、中、高。染色细胞的比例(<25%、25%~75%、>75%)和染色强度(阴性、弱、中等、强)构成了评分系统。
1.5 免疫细胞相关性分析
白癜风病灶内免疫微环境对疾病发生发展具有十分重要的作用。ssGSEA 方法是使用23 个免疫细胞基因集构建的。本研究中通过R 软件GSVA 包中ssGSEA 方法评估关键基因免疫细胞浸润特征[30]。
1.6 小分子化合物的筛选
在cMAP 数据库中(https://clue.io/query)筛选通过铁死亡途径治疗白癜风的潜在小分子化合物。将关键靶点分别上传至cMAP 数据库,通过数据库高通量分析,筛选出对其具有潜在作用的小分子化合物。按照高通量筛选得分由底到高排列,删去无归类的小分子,选取前30 个化合物作为后续研究的主要对象。
1.7 分子对接
利用分子对接初步验证化合物与潜在靶点结合,结合能<0 kcal/mol 说明配体分子与受体蛋白可以自发结合。在本研究中,通过PDB 数据库(https://www.rcsb.org/)[31]下载靶点蛋白的3D 结构,在AutoDockTools 1.5.7 软件中处理后保存为PDBQT格式[32]。通过PubChem 下载小分子化合物的3D 结构式,以同样的方式处理后保存为PDBQT 格式[33]。将上述2 个结构文件导入到AutoDockTools 1.5.7 中进行对接并计算结合能,使用OpenBabel 软件[34]将对接结果的PDBQT 格式转换为PDB 格式,随后使用PyMOL 软件将对接结果可视化。
1.8 关键基因的GSEA-京都基因与基因组百科全书(KEGG)富集分析
利用 R 语言 GSEA 包分析关键基因与GSE53146 数据集中其他基因之间的相关性。将所有基因根据其相关性从高到低排序,作为用来测试的基因集。同时将KEGG 信号通路集作为预定义集调用,以检测关键基因在信号通路集中的富集。
1.9 统计学分析
在本研究中使用Wilcoxon 秩和检验进行两组之间的比较。采用Spearman 相关性分析23 个白癜风铁死亡基因之间的关系,Cytoscape 用于ceRNA网络可视化。
2 结果
2.1 获取差异表达基因及铁死亡基因
将GSE53146 矫正后进行差异表达基因分析(图1A、1B),共获得706 个DEGs,其中包括412个上调基因和294 个下调基因,DEGs 热图及火山图如图1C、D 所示。在FerrDB 数据库中获取3 个模块相关数据,汇总去重后共得到458 个铁死亡标志物。
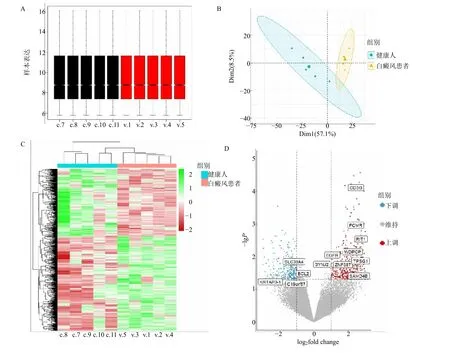
图1 GEO 数据集处理与差异表达基因分析Fig.1 GEO dataset processing and differential expression gene analysis
2.2 白癜风铁死亡关键基因的筛选
将706 个DEGs 对458 个FRGs 进行映射,筛选出23 个FR-DEGs(图2A)。FR-DEGs 之间的Spearman 相关性如图2B 所示。箱线图展现了白癜风患者大多数FR-DEGs 表达水平较健康人高(P<0.05、0.01,图2C)。获得FR-DEGs 后,利用LASSO算法筛选出6 个基因(图3A),利用SVM-RFE 筛选出18 个基因(图3B),取交集得核糖核苷二磷酸还原酶亚基M2(RRM2)、组织蛋白酶B(CTSB)、OTU 去泛素化酶 1(OTUB1)、α-突触核蛋白(SNCA)、脂质运载蛋白2(LCN2)、人含WW 域转录调节蛋白1(WWTR1)6 个关键基因(图3C)。如图3D 所示关键基因间具有较高相关性。关键基因的ROC 曲线说明,OTUB1和WWTR1在6 个特征基因中具有最高的AUC 值(AUC=0.960)。RRM2、CTSB、SNCA、LCN2的AUC 值分别为0.920、0.840、0.800、0.920(图3E)。以上结果表明,在白癜风患者和健康人之间6 个特征基因都具有较好的鉴别价值。
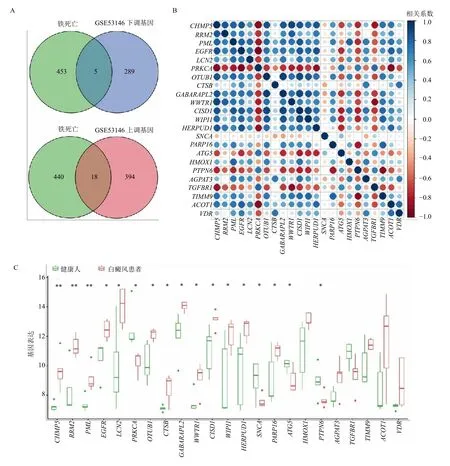
图2 铁死亡基因的筛选及表达分析Fig.2 Screening and expression analysis of ferroptosis genes

图3 关键基因的筛选与分析Fig.3 Screening and analysis of key genes
2.3 关键基因的外部数据集验证
在GSE75819 验证集中,WWTR1 在两组之间表达无显著差异,RRM2、CTSB、OTUB1、SNCA、LCN2 基因的表达趋势与实验集中完全一致且差异具有显著性。与健康人组相比,白癜风患者的RRM2、LCN2、OTUB1、CTSB 表达水平升高(P<0.01、0.001),而SCNA水平下降(P<0.05),见图4。由于WWTR1在验证集中基因表达无差异,故在后面的分析中只针对有差异的5 个关键基因。
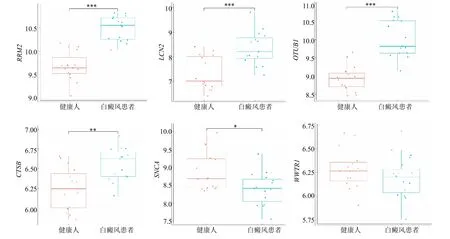
图4 关键基因在验证集中的差异分析Fig.4 Differential analysis of key genes in the validation set
2.4 关键基因在白癜风相关细胞群中表达
使用Seurat 函数分析了数据集GSE203262,并依据质量控制条件(nFeature_RNA>200 &nFeature_RNA<2 500 &percent.mt<5)从数据集中筛选33 694 个细胞,细胞主成分分析如图5A 所示。这些细胞被分类为9 种主要细胞类型,包括NK 细胞、朗格汉斯细胞、黑素细胞、B 细胞、血小板、树突状细胞、角质形成细胞和CD8+T 细胞(图5B)。随后在各细胞类型中对CTSB、LCN2、OTUB1、RRM2和SNCA基因进行富集标注。结果显示,白癜风患者皮肤样本中关键基因主要在成纤维细胞,角质形成细胞、黑素细胞和朗格汉斯细胞中。其中OTUB1在各种细胞中均被标注,说明其在白癜风发生发展过程中具有重要影响。此外,CTSB主要在成纤维细胞中标注。LCN2标注较弱几乎无表达。RRM2主要标注在CD8+T 细胞中,且富集结果具有高度特异性。成纤维细胞、角质形成细胞、黑素细胞和朗格汉斯细胞与SNCA高度相关(图5C)。

图5 关键基因在白癜风发病相关细胞群中的表达情况Fig.5 Immunoinfiltration assessment of key genes in the single cell data set
2.5 关键基因蛋白表达验证
在HPA 数据库中,对关键基因蛋白在正常人类皮肤细胞群(主要发现成纤维细胞、角质形成细胞、朗格汉斯细胞和黑素细胞)中的表达分析发现,CTSB、OTUB1在皮肤组织中表达最强,各细胞均能检测到而且细胞比例较高。LCN2与RRM2几乎检测不到。而SNCA很特殊,其只在黑素细胞中表达水平高,染色强度强并且细胞占比均为75%~25%,见图6。

图6 在HPA 数据库中验证关键基因的蛋白表达情况Fig.6 Validation of protein expression of key genes in the HPA database
2.6 关键基因的免疫细胞相关性
关键基因与白癜风病灶微环境23 个免疫细胞Spearman 相关性分析表明CTSB 与gamma delta T细胞和活化的树突状细胞呈显著正相关(图7A)。LCN2 与Th1 型辅助T 细胞、自然杀伤细胞、巨噬细胞、活化的CD8 T 细胞及肥大细胞呈显著正相关(图7B)。OTUB1 与自然杀伤细胞、活化的CD8 T细胞、活化的CD4 T 细胞、Th1 型辅助T 细胞、CD56+自然杀伤细胞、巨噬细胞及肥大细胞呈显著正相关(图7C)。RRM2 与活化的CD4 T 细胞、嗜酸性粒细胞、自然杀伤细胞及髓系抑制性细胞呈显著正相关,与效应性记忆CD4 T 细胞呈显著负相关(图7D)。此外,SNCA 是唯一一个与中性粒细胞具有相关性也是唯一一个几乎与所有的免疫细胞呈负相关的基因(图7E)。
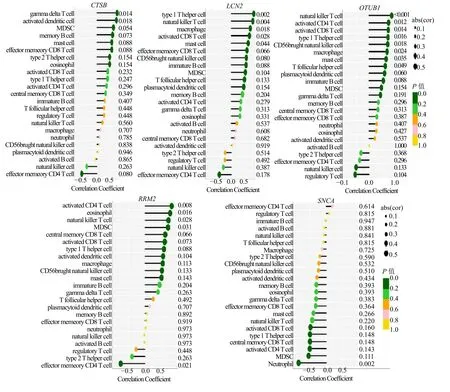
图7 关键基因与免疫细胞的相关性分析Fig.7 Analysis of the correlation between key genes and immune cells infiltration
2.7 关键基因的潜在信号通路富集
通过对关键基因进行单基因GSEA-KEGG通路富集分析(图8),发现这些基因主要涉及黏附分子、趋化因子、免疫反应(趋化因子及其受体相互作用、ECM-受体相互作用和Th17 细胞分化)、细菌或病毒感染(沙门菌感染、结核杆菌感染、流感病毒和趋化因子与病毒蛋白相互作用)和各种疾病途径(阿尔兹海默病、系统性红斑狼疮、肌萎缩侧索硬化和人类Ⅰ型白血病)。此外,关键基因也富集在趋化因子信号通路、机体代谢信号通路、NOD 样受体信号通路和T 细胞受体信号通路。同时,还发现趋化因子-趋化因子相互作用通路与CTSB、RRM2、SNCA3 个基因密切相关。同样,LCN2和OTUB12 个基因都与代谢通路密切相关,说明趋化因子相互作用及机体代谢在白癜风的发病及雷公藤甲素治疗中具有重要作用。

图8 关键基因CTSB、LCN2、OTUB1、RRM2 和SNCA 的GSEA-KEGG 分析Fig.8 GSEA-KEGG analysis of the key genes CTSB,LCN2,OTUB1,RRM2 and SNCA
2.8 筛选通过铁死亡途径治疗白癜风的潜在药物
在cMAP 数据库筛选的潜在药物中,将得分(norm_cs)<-1 的化合物作为候选化合物,在排名前30 位的成分中,选取具有明确化学结构式的雷公藤甲素[35](相对分子质量360.4,分子式C20H24O6)作为潜在药物,结构见图9。

图9 雷公藤甲素的药物结构Fig.9 Pharmacological structure of triptolide
2.9 雷公藤甲素与潜在靶点分子对接验证
潜在靶点CTSB(PDB:8b5f)、LCN2(PDB:3bx8)、OTUB1(PDB:4dhi)、RRM2(PDB:3bs9)和SNCA(PDB:4bxl)与雷公藤甲素(PubChem CID:107985)的分子对接结果如表1、图10 所示。结合能越低,配体与受体结合的构象就越稳定,产生作用的可能性就越大[36]。通过分子对接结果可以看出,雷公藤甲素对5 个潜在核心靶点的结合能力较强,从而提示其有望通过铁死亡途径治疗白癜风相关疾病。

表1 雷公藤甲素与潜在靶点的分子对接Table 1 Molecular docking information for triptolide and potential targets

图10 雷公藤甲素与潜在靶点的分子对接示意图Fig.10 Schematic diagram of molecular docking of triptolide with potential targets
3 讨论
铁死亡为铁依赖性脂质过氧化导致机体氧化还原失衡所介导的细胞死亡模式。在白癜风的研究中,与健康人组相比白癜风患者的血清中检测到花生四烯酸降低,这可能增加白癜风局部被CD8+T 细胞浸润及破坏黑素细胞的风险[37]。另外,铁死亡关键因子GPX4 在白癜风患者皮肤中表达降低,但在健康人组皮肤的各个表皮层中表现出高表达[38],这可能是白癜风局部铁死亡敏感性高和抗氧化能力弱的主要原因。然而,关于铁死亡在白癜风发病中是否占据着重要地位,研究甚少。
本研究共筛选了CTSB、LCN2、OTUB1、RRM2、SNCA5 个铁死亡基因进行一系列分析。其中RRM2是核糖核苷酸还原酶的催化亚基,它在增殖、迁移和血管生成等细胞过程中起重要作用[39-40]。研究表明,RRM2 的过表达激活NF-κB 通路并增加了胰腺癌细胞、乳腺癌细胞的侵袭、迁移[41-42]。而NF-κB通路也参与白癜风的发病,研究发现NF-κB 信号激活可促进白细胞介素(IL)-15、CXCL10 和IL-1β的分泌,进而促进CD8+T 细胞的活化和诱导白癜风的适应性免疫。此外,表皮中过量的CXCL10 在CD8+T 细胞的招募及黑素细胞杀伤中起关键性作用[43]。LCN2 是一种糖蛋白,可通过激活炎症途径或调节细胞铁稳态来调节细胞反应。研究发现,MPTP 中毒而引发的帕金森综合征小鼠模型中铁的积累使星形胶质细胞分泌LCN2 增加导致多巴胺神经元变性[44]。相关研究也发现在星形胶质细胞中NF-κB 信号的激活,能够调节LCN2 分泌和LCN2诱导脑中风的铁死亡[45]。因此,推测在白癜风病变中铁死亡基因RRM2上调导致NF-κB 信号被激活,CXCL10 与IL-1β 释放增多,招募CD8+T 免疫细胞局部浸润从而杀伤黑素细胞,同时NF-κB 信号激活也使LCN2 上调并诱导病变局部铁死亡,一方面使黑素细胞死亡,另一方面进一步增强RRM2的表达。
OTUB1 是一种去泛素化酶,影响细胞代谢、分化、增殖和凋亡,被称为是免疫细胞活性和炎症反应的重要调节剂[46]。在以往研究中发现,OTUB1 在多发性硬化症、支气管哮喘、肺癌、食管癌等多种疾病中有重要免疫调节作用[47-48];在多发性硬化症中,OTUB1 直接抑制STAT1mRNA 产生,还可以通过稳定SOCS1间接抑制IFN-γ诱导的JAK/STAT1信号传导[49]。在体外实验中,OTUB1 缺乏能增加IFN-γ 诱导的星形胶质细胞趋化因子(CXCL10、CXCL11、CCL2)和促炎分子(NOS2)的产生[50]。在白癜风研究中,IFN-γ 激活成纤维细胞中JAK/STAT1 通路使趋化因子CXCL9/10 分泌是CD8+T 免疫细胞在黑素细胞周围募集并杀伤细胞的关键一环[51]。在本研究中发现,白癜风病变皮肤中OTUB1 表达上调,可能是机体产生的免疫保护作用。OTUB1 上调抑制JAK/STAT1 信号传导,抑制CXCL9/10 产生的免疫细胞招募能力,减少CD8+T 在病变局部的浸润,从而避免黑素细胞的死亡。
关键基因GSEA-KEGG 通路分析发现,趋化因子相互作用通路和NOD 样受体信号通路与关键基因密切相关。趋化因子是由多种免疫细胞分泌的重要的炎症介质,其可以招募CD8+T 免疫细胞在黑素细胞周围募集,从而杀伤细胞使黑色素产生障碍[52]。NOD 样受体是细胞内受体,可以激活各种相关途径,以增加促炎细胞因子(如IFN-γ、IL-1、IL-6 和TNF)的产生。细胞因子的产生能增加免疫细胞反应并激活非特异免疫系统[53]。
Connectivity Map(cMAP)数据库是由哈佛、剑桥大学和麻省理工学院研究人员通过不同干扰物(包括小分子)处理人类细胞后检测基因表达差异所构建的生物应用数据库。研究团队认为以基因表达谱建立的基因与药物的关联性,可协助研究者快速利用基因表达数据比对出与基因高相关性的药物、并推论出药物分子的主要结构。本研究基于“新机制-老药”研究思路,通过cMAP 数据库筛选出影响5 个潜在靶点的潜在药物。分子对接结果表明,雷公藤甲素对5 个潜在靶点的结合较强,说明该小分子与筛选出的铁死亡相关靶点和通路之间存在高度关联性,其有望成为通过铁死亡途径治疗白癜风药物。目前相关研究也表明,雷公藤甲素在皮肤病与自身免疫性疾病的治疗方面具有很大潜力[53-54]。
白癜风发病机制复杂且不明确,导致对其治愈十分困难,因此阐明白癜风的发病机制对其根治尤为重要。本研究采用生物信息学方法从铁死亡的角度筛选了白癜风发病的5 个关键基因,并采用多种数据评估了5 个铁死亡基因与病变部位细胞群潜在联系,所得结果可靠性强。除此之外,还筛选雷公藤甲素作为铁死亡途径治疗白癜风小分子药物。所采用的“新机制-老药”研究思路,不仅可以大大降低药物研发中因不良反应所导致的失败,也为临床上老药的重新应用提供了新的方向。本研究为白癜风的发病及治疗研究提供了新的视角,适合在其他疾病研究中推广,通过新的机制与老药的结合研究,将会为临床更快地研发治疗疑难疾病的药物提供参考方案。
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
天然产物研究与开发(2018年4期)2018-05-07
中国继续医学教育(2015年2期)2016-01-06
中国医药生物技术(2015年4期)2015-12-26
山东医药(2015年38期)2015-12-07
中国医疗美容(2015年4期)2015-04-27
国际皮肤性病学杂志(2015年4期)2015-03-20
现代检验医学杂志(2015年6期)2015-02-06
中国医疗美容(2015年5期)2015-02-03
中国药理学通报(2014年2期)2014-05-09
中成药(2014年11期)2014-02-28
