
PEG-6000模拟干旱胁迫对冬黑麦生理指标的影响
2024-05-07 04:31乌日力嘎邓宇孙墨可
浙江农业科学 2024年4期
乌日力嘎,邓宇*,孙墨可
(1.长春大学 园林学院,吉林 长春 130022;2.白城市农业科学院,吉林 白城 137099)
黑麦(SecalecerealeL.)是禾本科黑麦属栽培作物,有春黑麦和冬黑麦之分,冬黑麦可以在东北地区种植且越冬生长[1],抗逆性较强,广泛种植冬黑麦可以提高冬季地表覆盖,对提高粮食单产和保证粮食安全均有一定的积极作用[2]。冬黑麦富含膳食纤维、蛋白质、维生素、不饱和脂肪酸等生理活性成分,具有很高的营养价值和经济价值,在人类生活中具有重要意义[3]。干旱是严重影响植物生长发育的非生物胁迫之一,并且影响广泛而深远。中国作为农业大国,在世界上属于水资源短缺的国家之一,再受全球气候变暖的影响,旱灾更是频繁发生,水资源短缺的矛盾也日益突出,对农业生产带来严重危害,甚至会直接危害到粮食安全问题[4]。干旱胁迫下,植物会产生活性氧,导致细胞膜系统的代谢紊乱,损坏细胞功能,使细胞保水能力下降,导致植物无法正常生长发育甚至死亡。通过测定冬黑麦叶片各项生理指标可以反映出其在干旱胁迫条件下的生长规律等。目前,科研人员对冬黑麦研究多以抗寒为主,采用PEG-6000模拟干旱胁迫及分子生物学水平研究较少,因此,本试验以俄罗斯冬黑麦BK01为研究对象,采用15% PEG-6000对水培冬黑麦模拟干旱胁迫,测定各胁迫时期冬黑麦叶片多项生理指标,明确干旱胁迫下冬黑麦的生理特性,为进一步干旱胁迫下冬黑麦转录组测序研究及探究其抗旱分子机制奠定基础。
1 材料与方法
1.1 试验材料
俄罗斯冬黑麦BK01由吉林省白城市农业科学院保存。
1.2 试验方法
1.2.1 冬黑麦胁迫处理
选择健康、颗粒饱满的冬黑麦种子,用0.1%高锰酸钾溶液浸泡消毒15 min,将消毒后的种子平铺于培养皿中湿润的滤纸上,置于25 ℃培养箱中暗培养2 d至种子发芽。将发芽的冬黑麦种子转入含1 L霍格兰德营养液的培养盘中并移至光照培养室继续培养(25 ℃,光照16 h,黑暗8 h),营养液每天更换1次。将冬黑麦水培至三叶一心后,利用15% PEG-6000模拟干旱胁迫,处理时间为0(CK)、3、6、9、12和24 h,每组处理重复3次,收集冬黑麦叶片,用于生理指标测定。
1.2.2 冬黑麦生理指标的测定
分别采用烘干称重法测定相对含水率(RWC),采用TBA法测定丙二醛(MDA)含量[5],采用蒽酮法测定可溶性糖(SS)含量[6],采用考马斯亮蓝法测定可溶性蛋白(SP)含量[6],采用比色法测定脯氨酸(Pro)含量[7],采用可见光法测定过氧化氢酶(CAT)活性[8],采用比色法测定过氧化物酶(POD)活性[9],采用羟胺法测定超氧化物歧化酶(SOD)活性[10]。
2 结果与分析
2.1 干旱胁迫对冬黑麦叶片相对含水率的影响
相对含水率通常能够直观地表现出植物的有效含水量。在15% PEG-6000模拟干旱胁迫下随着时间的延长冬黑麦叶片的相对含水率呈现下降趋势。与0 h相比,冬黑麦叶片在干旱胁迫3、6、9、12、24 h的RWC极显著降低,在胁迫24 h时RWC达最低,为59.028%(图1)。因此,干旱胁迫导致冬黑麦叶片失水,相对含水率下降,对植物造成损害。

“**”表示差异极显著(P<0.01)。图1 干旱胁迫对冬黑麦叶片相对含水率的影响Fig.1 Effects of drought stress on RWC content of winter rye leaves
2.2 干旱胁迫对冬黑麦叶片丙二醛含量的影响
丙二醛含量能够反映出膜脂过氧化的程度,也能间接反映抗旱能力的强弱。在15%PEG-6000模拟干旱胁迫下随着时间的延长冬黑麦叶片的丙二醛含量呈现上升趋势。在干旱胁迫时间达24 h时,冬黑麦叶片丙二醛含量最高,为7.741 μmol·g-1,且9、12、24 h的丙二醛含量与0 h相比极显著升高,6 h的丙二醛含量与0 h相比显著升高(图2)。随着干旱胁迫时间延长,冬黑麦叶片丙二醛含量的累积,使膜脂过氧化,导致细胞膜流动性降低,透性增强。

“*”表示差异显著(P<0.05);“**”表示差异极显著(P<0.01)。图3~8同。图2 干旱胁迫对冬黑麦叶片丙二醛含量的影响Fig.2 Effects of drought stress on MDA content of winter rye leaves
2.3 干旱胁迫对冬黑麦叶片可溶性糖含量的影响
在15% PEG-6000模拟干旱胁迫下冬黑麦叶片可溶性糖含量变化呈先上升后下降趋势,当胁迫时间达9 h时,冬黑麦叶片可溶性糖含量最高,为8.536 μg·g-1。冬黑麦叶片在干旱胁迫3、12 h的可溶性糖含量与0 h相比显著升高,胁迫6和9 h叶片的可溶性糖含量与0 h相比极显著升高(图3)。干旱导致冬黑麦叶片细胞失水,可溶性糖含量升高能够降低细胞失水性,维持新陈代谢正常进行。
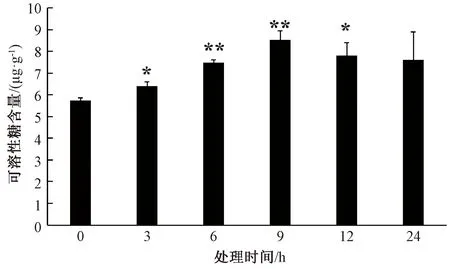
图3 干旱胁迫对冬黑麦叶片可溶性糖含量的影响Fig.3 Effects of drought stress on soluble sugar content in winter rye leaves
2.4 干旱胁迫对冬黑麦叶片可溶性蛋白含量的影响
可溶性蛋白与可溶性糖是植物重要的渗透调节物质,在干旱胁迫中起到提高植物保水能力的作用。在15% PEG-6000模拟干旱胁迫下,冬黑麦叶片可溶性蛋白含量变化,呈先上升后下降的趋势。冬黑麦叶片在干旱胁迫处理3、6 h的可溶性蛋白含量与0 h相比显著上升,9、12、24 h的可溶性蛋白含量与0 h相比极显著上升。当胁迫时间达9 h时冬黑麦叶片可溶性蛋白含量达最高,为17.702 mg·g-1(图4)。冬黑麦叶片可溶性蛋白含量的增加能够提高细胞保水能力,增强抗旱性。

图4 干旱胁迫对冬黑麦叶片可溶性蛋白含量的影响Fig.4 Effects of drought stress on soluble protein content in winter rye leaves
2.5 干旱胁迫对冬黑麦叶片脯氨酸含量的影响
脯氨酸是植物重要的渗透调节物质,在干旱胁迫中会主动积累。在15% PEG-6000模拟干旱胁迫下冬黑麦叶片脯氨酸含量呈上升趋势,在24 h达到最高,为66.985 μg·g-1。冬黑麦叶片在干旱胁迫处理6、9 h的脯氨酸含量显著高于0 h,12、24 h的脯氨酸含量与0 h相比极显著上升(图5)。随着干旱胁迫时间延长,冬黑麦叶片脯氨酸含量逐渐升高,能够减缓叶片组织失水。

图5 干旱胁迫对冬黑麦叶片脯氨酸含量的影响Fig.5 Effects of drought stress on proline content in winter rye leaves
2.6 干旱胁迫对冬黑麦叶片酶活性的影响
在干旱胁迫条件下植物体内产生大量活性氧,保护性酶可以有效地清除积累的自由基,起到保护细胞的作用。在干旱条件下CAT和POD活性的作用是分解代谢中产生的H2O2,以避免H2O2积累对细胞的氧化破坏作用。在15% PEG-6000模拟干旱胁迫下冬黑麦叶片CAT活性随着干旱胁迫时间的延长,呈先上升后下降趋势,9、12、24 h的CAT活性与0 h相比极显著上升,且干旱胁迫处理9 h时冬黑麦叶片CAT活性达到最高,为9.109 U·mg-1,6 h的CAT活性与0 h相比显著上升(图6)。

图6 干旱胁迫对冬黑麦叶片过氧化氢酶活性的影响Fig.6 Effect of drought stress on CAT activity in winter rye leaves
在15% PEG-6000模拟干旱胁迫下,冬黑麦叶片POD活性变化,随着干旱胁迫时间的延长,冬黑麦叶片POD活性呈先上升后下降趋势。当胁迫12 h时,冬黑麦叶片POD活性达到最大值,9、12、24 h时的POD活性与0 h相比显著上升(图7)。

图7 干旱胁迫对冬黑麦叶片过氧化物酶活性的影响Fig.7 Effect of drought stress on POD activity in winter rye leaves
在逆境胁迫条件下,SOD可以有效地清除植物的自由基,减少活性氧的含量,起到保护细胞的作用。在15% PEG-6000模拟干旱胁迫下冬黑麦叶片SOD活性变化,随着干旱胁迫时间的延长,冬黑麦叶片SOD活性呈先上升后下降趋势,胁迫处理9、12、24 h的SOD活性与0 h相比极显著升高,在9 h时冬黑麦叶片SOD活性达到最高。冬黑麦叶片胁迫6 h的SOD活性与0 h相比显著上升(图8)。干旱胁迫导致冬黑麦叶片细胞内活性氧积累,SOD能够减少活性氧积累。

图8 干旱胁迫对冬黑麦叶片超氧化物歧化酶活性的影响Fig.8 Effect of drought stress on SOD activity in winter rye leaves
3 讨论
干旱胁迫下相对含水率能够表明植物的水分状况的生理意义,能够表现出植物的有效水分含量。刘硕等[11]以甘蔗为实验材料探索抗旱品种选育,发现干旱1 d时叶片相对含水率变化不明显,但随着干旱加剧,叶片相对含水率显著降低,干旱9 d时云蔗05-51和粤糖93-159的叶片相对含水率仅为11.64%和8.89%,复水10 d后虽有所回升,但仍显著低于对照。一些研究[12]发现在干旱胁迫期间,细胞的膨胀压力降低,这减少了芽长、生物量和植物生长。干旱会导致叶片枯萎,植物生长减少,花蕾形成减少,叶和叶面积减少,Rollins等[13-14]研究发现,干旱胁迫是植物鲜重和干重、叶片数、总叶面积、相对含水量、气孔大小和频率降低的原因。本研究中冬黑麦相对含水率呈逐渐下降趋势,在干旱胁迫0 h时达最高,24 h时最低,与上述结果相似。
干旱胁迫时,膜脂过氧化导致MDA积累,使蛋白质和核酸变性,导致膜脂流动性降低,透性增强。MDA含量的高低在一定程度上能反映膜脂过氧化作用水平和膜结构的受害程度,MDA含量越高品种抗旱能力越弱[7]。王晓雪等[7]研究表明,燕麦生长早期根系抗氧化酶活性和脯氨酸含量随干旱胁迫程度增加而增加。张翠梅等[15]研究干旱胁迫对不同抗旱苜蓿品种根系生长特征及生理特性的影响发现,干旱胁迫显著增加了供试苜蓿根系的MDA含量和质膜相对透性,说明供试苜蓿的膜脂过氧化水平均随干旱胁迫程度增加而显著增加。本实验结果与上述结果相似,随着干旱胁迫时间的延长,在24 h时叶片MDA含量达到最高值,在干旱胁迫下叶片受到伤害,反映了干旱胁迫下冬黑麦叶片逆境损伤,产生了大量的MDA。
可溶性糖是植物体内重要的渗透调节物质和代谢产物,在干旱胁迫中,可溶性糖类是参与渗透调节的重要物质[16]。可溶性糖含量与植物叶片生理代谢调节能力和光合作用有关,叶片中可溶性糖含量随着干旱胁迫时间的延长而增加,抗旱品种的可溶性糖含量高。苏玮娟等[17]研究表明,随胁迫时间的延长,可溶性糖含量总体呈上升趋势,在胁迫72 h时达到峰值,青引1号和定燕2号的可溶性糖含量较高,甜燕麦的可溶性糖含量最低。随着胁迫时间的进一步延长,胁迫96 h时可溶性糖含量稍微降低。本研究中,冬黑麦可溶性糖含量呈先上升后下降趋势,9 h时可溶性糖含量达最高,通过升高细胞液的渗透势吸收水分,随着胁迫时间的延长,可溶性糖含量开始下降,自我调节能力下降。可溶性糖含量升高说明干旱胁迫下,冬黑麦通过可溶性糖含量的增加,降低细胞失水性,使自身新陈代谢能够正常进行。
可溶性蛋白是重要的渗透调节物质和营养物质。在干旱胁迫中,可溶性蛋白的增加和积累能提高细胞的保水能力,对细胞的生命物质及生物膜起到保护作用。在干旱胁迫下可以通过植物中可溶性蛋白含量这一指标来反映植物的抗旱性。很多研究证明,可溶性蛋白含量的变化与干旱强度有直接关系,随着干旱胁迫强度的增加可溶性蛋白的含量表现为先增强后减弱;干旱胁迫对蛋白含量的影响各异,抗旱性强的品种能诱导基因更强的表达以适应干旱胁迫,而抗旱性较弱的品种可能减弱或丧失其自我调节能力[6]。本实验冬黑麦叶片可溶性蛋白含量9 h最高,自我调节能力最强,随着胁迫时间调节能力下降,表明冬黑麦叶片细胞通过增加可溶性蛋白含量使细胞保水能力提高,使得适应性增强,这与上述结果相似。
脯氨酸是植物细胞内重要的渗透调节物质,在逆境条件下脯氨酸会主动积累,受干旱胁迫的植物在植物的叶子、芽和根尖等区域表现出较高的脯氨酸浓度。植物中脯氨酸的积累可引起细胞渗透势下降,提高细胞的吸水能力或使水分外渗,有助于细胞和组织的持水,使植物能够吸收水分进行生长和代谢[18]。Nasrin等[19]研究发现,水稻品种BRRI Dhan-24在干旱胁迫下脯氨酸积累量增加。许多抗旱研究将脯氨酸含量的积累量作为植物抗旱指标之一,本实验中冬黑麦的脯氨酸含量随着胁迫时间的延长呈上升的趋势,24 h时达最高,与上述研究相似。脯氨酸含量升高说明了干旱胁迫下冬黑麦细胞通过增加脯氨酸的含量,使细胞渗透势降低,阻止水分丧失,使自身新陈代谢能够正常进行,响应了干旱胁迫。
逆境条件下植物细胞会产生过氧化物,导致细胞膜系统代谢紊乱,损坏细胞功能。此时植物会产生CAT、POD、SOD、APX等保护酶,减少活性氧的含量,起到保护细胞的作用。苏玮娟等[17]研究表明,除甜燕麦的SOD活性随胁迫时间的延长一直上升外,其余材料的CAT、POD及SOD活性均随胁迫时间的延长呈先升后降趋势。本实验中冬黑麦叶片的3个酶活性随胁迫的加剧均呈先上升后下降趋势。其中,CAT与SOD活性在9 h时达到最高,POD活性在12 h时达到最高,与上述研究结果相似。结合本实验,在干旱胁迫初期叶片内的过氧化物被CAT、POD和SOD活性分解或清除,缓解氧化的速度,随着胁迫时间的延长冬黑麦叶片活性氧清除功能逐渐降低。
猜你喜欢
中老年保健(2022年3期)2022-11-21
中国农学通报(2022年21期)2022-08-22
科学与财富(2019年15期)2019-10-21
现代家庭·生活版(2019年1期)2019-01-24
中成药(2018年8期)2018-08-29
发酵科技通讯(2018年2期)2018-07-06
湖南林业科技(2017年1期)2017-02-06
医学研究杂志(2015年9期)2015-07-01
三联生活周刊(2015年21期)2015-05-21
中国当代医药(2015年9期)2015-03-01
