
miR-660-3p靶向NFAM1对RSV诱导支气管上皮细胞凋亡及炎症因子分泌的影响
2024-05-07 05:12:00童珍珍高雅丽许娟娟黄方许诣陈霞
山东医药 2024年13期
童珍珍,高雅丽,许娟娟,黄方,许诣,陈霞
1 上海市嘉定区安亭医院呼吸内科,上海 201805;2 华中科技大学同济医学院附属协和医院呼吸内科;3 湖北文理学院附属医院 襄阳市中心医院儿科
支气管哮喘是一种以慢性气道炎症和气道高反应性为主要特征的异质性疾病,其发病机制至今仍不完全清楚[1]。气道上皮细胞是气道屏障的重要组成部分。气道上皮细胞损伤会引起气道炎症和气道重塑,从而导致支气管哮喘患者气道阻塞和气道高反应性[2]。因此,深入探索气道上皮细胞损伤的分子机制对支气管哮喘的防治具有重要意义。微小RNA-660-3p(miR-660-3p)在细胞增殖、凋亡及组织纤维化等生理和病理过程中发挥重要作用。有研究报道,miR-660-3p 在心力衰竭患者心肌组织及血浆中表达上调,并可作为诊断心力衰竭的潜在生物标志物[3];在肝纤维化小鼠肝组织中miR-660-3p 高表达,抑制其表达可减轻肝纤维化程度,这为肝纤维化治疗提供了潜在的靶标[4]。但目前miR-660-3p在支气管哮喘发生、发展中的作用尚不清楚。miRNA 可通过与靶基因mRNA的3′非翻译区(3′UTR)完全或不完全互补结合,引起靶基因mRNA 降解或翻译抑制,从而参与各种生命活动[5]。在线生物信息学预测发现,具有ITAM 基序1 的NFAT 活化蛋白(NFAM1)为miR-660-3p 的靶基因。NFAM1 是一种携带ITAM 的跨膜受体,可参与调控炎症反应。有研究报道,在克罗恩病、溃疡性结肠炎患者肠道活检和炎症性肠病小鼠血清中均观察到NFAM1 表达增加,并且在单核细胞和中性粒细胞中亦观察到NFAM1 高表达[6-7]。但NFAM1 在支气管哮喘发生、发展中的作用尚不完全清楚。呼吸道合胞病毒(RSV)是诱发支气管哮喘的常见病原体,可引起气道炎症反应,进而促进支气管哮喘的发生、发展[8]。2022 年6 月—12 月,本研究探讨了miR-660-3p 靶向NFAM1 对RSV 诱导支气管上皮细胞凋亡及炎症因子分泌的影响。现报告如下。
1 材料与方法
1.1 材料 人支气管上皮细胞系16HBE,购自武汉普诺赛生命科技有限公司。RSV-Long 病毒2 型株,购自中国预防医学科学院病毒学研究所。miR-660-3p 模拟物(miR-660-3p mimics)及其对照序列(NC mimics)、NFAM1过表达载体(pcDNA-NFAM1)及其对照空载体(pcDNA-NC)、NFAM1 突变型质粒(NFAM1-MUT)及其野生型质粒(NFAM1-WT)以及PCR 引物,购自上海生工生物工程股份有限公司。NanoDrop 2000C 超微量分光光度计,购自美国Thermo Scientific 公司;ABI Prism®7500 型实时荧光定量PCR 仪,购自美国Applied Biosystems 公司;iMark680 多功能酶标仪,购自美国BIO-RAD 公司;FACSCalibur流式细胞仪,购自美国Becton-Dickinson公司。TRIzol、LipofectamineTM2000,购自美国Invitrogen公司。Annexin Ⅴ-FITC/PI 细胞凋亡检测试剂盒,购自北京索莱宝科技有限公司;SYBR®Premix Ex TaqTMⅡ荧光定量试剂盒、SuperScript Ⅳ逆转录酶试剂盒,购自大连宝生物工程有限公司;双荧光素酶报告基因检测试剂盒,购自美国Promega公司;TNF-α、IL-6、IL-1β ELISA试剂盒,购自南京建成生物工程研究所。NFAM1 单克隆抗体及兔抗人活化的半胱天冬酶3(Cleaved-Caspase-3)、β-actin 一抗和山羊抗兔IgG二抗,购自美国Santa Cruz公司。
1.2 细胞传代培养 将冻存的16HBE 细胞快速解冻复苏,接种于含10% FBS 的DMEM 培养基,然后置于37 ℃、5% CO2的细胞培养箱培养。待细胞生长至80%左右融合时,胰蛋白酶消化后计数,按1∶2传代。取对数生长期16HBE细胞进行后续实验。
1.3 细胞分组转染 取对数生长期16HBE 细胞,以2.5 × 105个/孔接种于6孔板,然后置于DMEM 培养基中培养24 h。随机分为对照组、RSV组、miR-NC +RSV组、miR-660-3p + RSV组、pcDNA-NFAM1 + RSV组、pcDNA-NC + RSV 组、miR-660-3p + pcDNA-NC +RSV 组、miR-660-3p + pcDNA-NFAM1 + RSV 组,按LipofectamineTM2000 说明,miR-NC + RSV 组转染NC mimics,miR-660-3p + RSV 组转染miR-660-3p mimics,pcDNA-NFAM1 + RSV 组转染pcDNANFAM1,pcDNA-NC + RSV 组转染pcDNA-NC,miR-660-3p + pcDNA-NC + RSV组共转染pcDNA-NC和miR-660-3p mimics,miR-660-3p + pcDNA-NFAM1 +RSV组共转染pcDNA-NFAM1和miR-660-3p mimics。对照组和RSV组不予转染。转染6 h,更换新鲜培养基继续培养24 h。然后,除对照组外,其余各组均予RSV 感染,MOI设定为0.000 1[9]。感染24 h,收集各组细胞及其培养液。
1.4 miR-660-3p、NFAM1 mRNA 表达检测 采用RT-qPCR法。收集各组上述细胞,采用TRIzol 法提取细胞总RNA,经NanoDrop 2000C 超微量分光光度计鉴定,OD260/OD280为1.8~2.0。按SuperScript Ⅳ逆转录酶试剂盒说明将总RNA逆转录合成为cDNA。逆转录条件:37 ℃ 10 min,45 ℃ 30 min,72 ℃ 4 min。以cDNA为模板,按SYBR®Premix Ex TaqTMⅡ荧光定量试剂盒说明进行PCR 扩增。引物序列:miR-660-3p上游引物5'-CGTAGAGCGGCTAGGGATCG-3'、下游引物5'-CGAATACTCTCTGAGCTGA-3',内参U6 上游引物5'-CGAGATCGATGGAAAACCCCG-3'、下游引物5'-AGGCGCGCGGGAACCTTTA-3';NFAM1 上游引物5'-CGAAACCTCCGAAGCAA-3'、下游引物5'-AAATTTGCGTCCGTGAC-3',内参GAPDH 上游引物5'-CGAGAGAAGTACCAAGATGAG-3'、下游引物5'-CGAGGCAGGCTTCGACG-3'。PCR 扩增体系共20 µL:2 × SYBR®Premix Ex TaqTM10 µL,cDNA模板1 µL,上下游引物各0.4 µL,ddH2O补足至20 µL;反应条件:95 ℃ 5 min,95 ℃ 10 s、58 ℃ 30 s、72 ℃ 30 s共35 个循环。PCR 扩增反应结束,绘制熔解曲线,获取循环阈值(CT)数。分别以U6 或GAPDH 为内参,采用2-ΔΔCT法计算目的基因相对表达量。
1.5 细胞凋亡检测 采用流式细胞术。收集各组上述细胞,胰蛋白酶消化,用结合缓冲液重悬制成密度为1 × 105/mL 细胞悬液。取细胞悬液1 mL,1 000 r/min 离心5 min、离心半径8 cm,弃上清。向沉淀中加入结合缓冲液500 µL 重悬细胞。然后依次加入Annexin Ⅴ-FITC 10 µL、PI 5 µL,室温避光孵育15 min,上流式细胞仪检测细胞凋亡率。
1.6 NFAM1、Cleaved-Caspase-3 蛋白表达检测 采用Western blotting 法。收集各组上述细胞,加入RIPA裂解液提取细胞总蛋白,经BCA法蛋白定量合格。加入适量蛋白上样缓冲液,100 ℃金属浴充分变性。取变性蛋白30 µg,SDS-PAGE分离。电泳结束,将蛋白电泳产物转印至PVDF膜上。5%脱脂奶粉室温封闭1 h,分别加入Cleaved-Caspase-3、NFAM1、β-actin一抗(稀释比分别为1∶500、1∶1 000、1∶1 000),4 ℃孵育过夜。次日,洗膜后加入山羊抗兔IgG 二抗(稀释比1∶1 000),37 ℃孵育1 h。ECL发光,暗室内曝光、显影、定影。采用Image J软件分析各蛋白电泳条带灰度值,以目的蛋白电泳条带灰度值与内参蛋白电泳条带灰度值的比值作为目的蛋白相对表达量。
1.7 培养上清液TNF-α、IL-6、IL-1β 检测 收集各组细胞培养液,3 500 r/min 离心10 min、离心半径8 cm,留取上清液。采用ELISA 法检测培养上清液TNF-α、IL-6、IL-1β。
1.8 miR-660-3p 与NFAM1 的靶向关系预测与验证 通过starBase数据库预测miR-660-3p与NFAM1的靶向结合位点。取对数生长期16HBE 细胞,以2.5 × 105个/孔接种于6 孔板,置于37 ℃、5% CO2的细胞培养箱内培养。待细胞生长至80%左右融合时,按LipofectamineTM2000 说明,将NFAM1-WT、NFAM1-MUT 分别与NC mimics、miR-660-3p mimics共转染。转染6 h,更换新鲜培养基继续培养24 h。收集细胞,按双荧光素酶报告基因检测试剂盒说明检测荧光素酶活性。
1.9 统计学方法 采用SPSS22.0 统计软件。符合正态分布的计量资料以±s表示,结果比较采用独立样本t检验。P<0.05为差异有统计学意义。
2 结果
2.1 RSV 组与对照组miR-660-3p、NFAM1、Cleaved-Caspase-3 表达以及细胞凋亡率和培养上清液炎症因子水平比较 见表1、2。
表1 两组miR-660-3p、NFAM1、Cleaved-Caspase-3表达比较(± s)
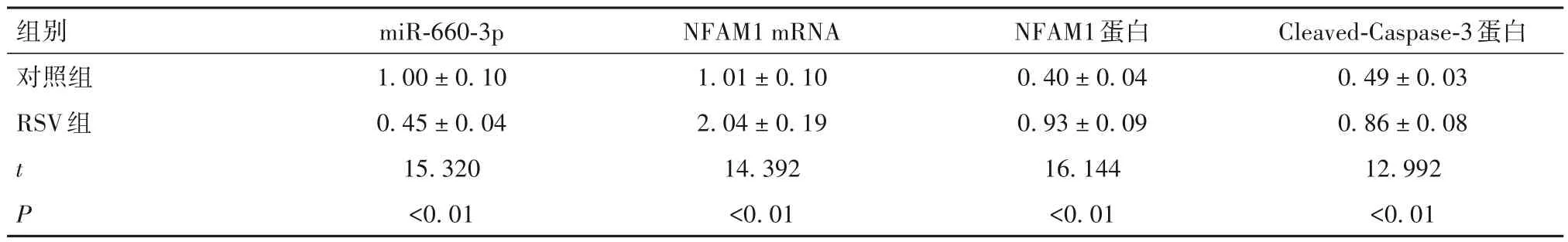
表1 两组miR-660-3p、NFAM1、Cleaved-Caspase-3表达比较(± s)
?
表2 两组细胞凋亡率及培养上清液炎症因子水平比较(± s)
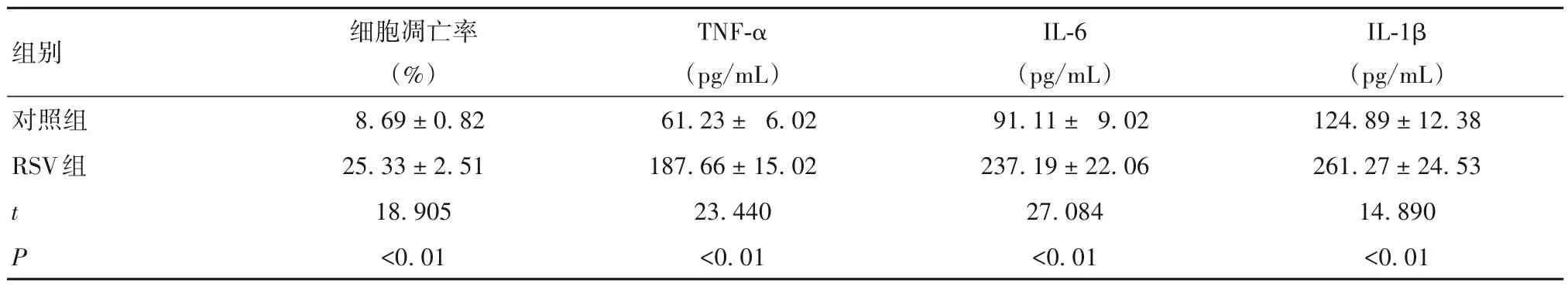
表2 两组细胞凋亡率及培养上清液炎症因子水平比较(± s)
?
2.2 上调miR-660-3p 表达对RSV 诱导支气管上皮细胞Cleaved-Caspase-3 蛋白表达以及细胞凋亡率和培养上清液炎症因子水平的影响 见表3。
表3 两组miR-660-3p、Cleaved-Caspase-3蛋白表达以及细胞凋亡率和培养上清液炎症因子水平比较(± s)
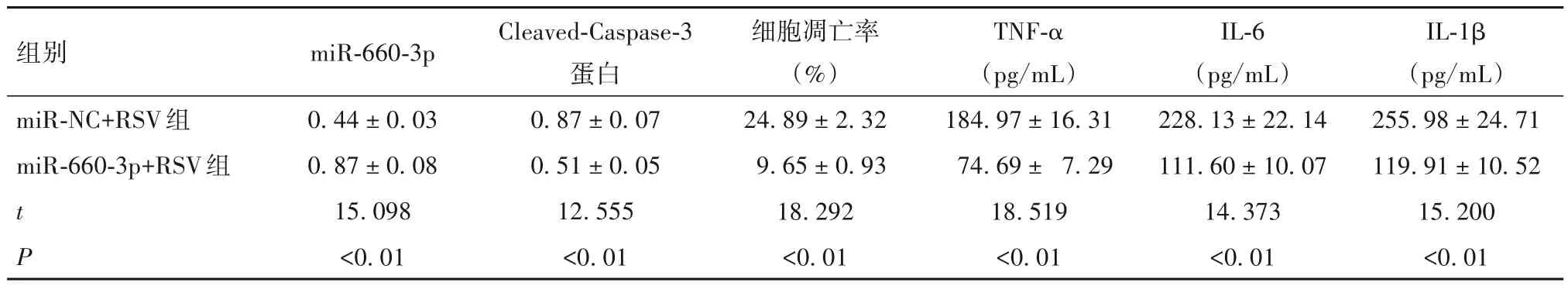
表3 两组miR-660-3p、Cleaved-Caspase-3蛋白表达以及细胞凋亡率和培养上清液炎症因子水平比较(± s)
?
2.3 上调NFAM1 表达对RSV 诱导支气管上皮细胞Cleaved-Caspase-3 蛋白表达以及细胞凋亡率和培养上清液炎症因子水平的影响 见表4。
表4 两组NFAM1 mRNA、Cleaved-Caspase-3蛋白表达以及细胞凋亡率和培养上清液炎症因子水平比较(± s)

表4 两组NFAM1 mRNA、Cleaved-Caspase-3蛋白表达以及细胞凋亡率和培养上清液炎症因子水平比较(± s)
?
2.4 miR-660-3p 与NFAM1 的靶向调控关系 经starBase 数据库预测,NFAM1 的3′UTR 存在与miR-660-3p 的结合位点,见图1。双荧光素酶报告基因实验结果显示,相较于共转染NC mimics 与NFAM1-WT 的16HBE 细胞,共转染miR-660-3p mimics与NFAM1-WT的16HBE细胞荧光素酶活性降低(0.40 ± 0.04 比1.00 ± 0.10;t=16.713,P<0.05);相较于共转染NC mimics 与NFAM1-MUT 的16HBE细胞,共转染miR-660-3p mimics 与NFAM1-MUT 的16HBE 细胞荧光素酶活性变化不明显(0.99 ± 0.08比1.02 ± 0.09;t=0.747,P>0.05)。

图1 miR-660-3p与NFAM1的靶向结合位点示意图
2.5 上调NFAM1、miR-660-3p 表达对RSV 诱导支气管上皮细胞Cleaved-Caspase-3 蛋白表达以及细胞凋亡率和培养上清液炎症因子水平的影响 见表5。
表5 两组Cleaved-Caspase-3蛋白表达以及细胞凋亡率和培养上清液炎症因子水平比较(± s)
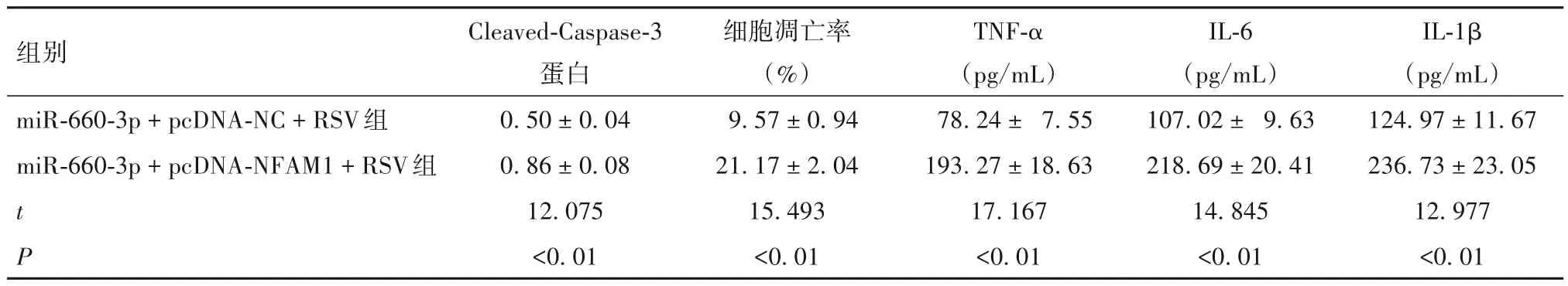
表5 两组Cleaved-Caspase-3蛋白表达以及细胞凋亡率和培养上清液炎症因子水平比较(± s)
?
3 讨论
支气管哮喘是一种慢性气道炎症性疾病,以慢性气道炎症和气道高反应性为主要特征。据统计,目前全球约有3 亿人罹患支气管哮喘,并且其患病率仍在不断上升[10]。支气管哮喘的发病机制至今仍不完全清楚,可能与气道上皮细胞损伤引起的炎症因子过度释放有关。
miRNA是一类在真核生物中广泛存在的内源性非编码小分子RNA,能够参与基因转录后的表达调控,从而广泛参与人体内各种生命活动,如细胞增殖、分化、凋亡等。有研究报道,多种miRNA在哮喘患者支气管黏膜或血清中异常表达,如miR-511-3p[11]、miR-144-3p[12],这些异常表达的miRNA 能够调控支气管上皮细胞凋亡以及不同程度炎症反应,可作为临床治疗支气管哮喘的分子靶点。miR-660-3p 是miRNA 家族成员之一,在细胞增殖、凋亡及组织纤维化等生理和病理过程中发挥重要作用。有研究报道,miR-660-3p在胃癌[13]、卵巢癌[14]和乳腺癌[15]等恶性肿瘤组织和细胞中低表达,其表达上调可抑制肿瘤细胞的增殖、侵袭和迁移等,可作为恶性肿瘤临床治疗的分子靶点。NFAM1为miR-660-3p的靶基因。NFAM1 是一种携带ITAM 的跨膜受体,在单核细胞和中性粒细胞中高表达,可促进炎症因子、趋化因子受体表达,是自身免疫性疾病潜在的治疗靶点。有研究报道,NFAM1 在克罗恩病、溃疡性结肠炎患者肠道活检中表达上调[6-7]。但目前miR-660-3p 和NFAM1在支气管哮喘发生、发展中作用的研究报道较少。
气道上皮细胞是气道防御的第一道屏障,在受到外界有害刺激后发生活化,释放一系列炎症介质、细胞因子和黏附分子等,加重哮喘患者气道炎症[16]。TNF-α、IL-6和IL-1β均属于促炎症细胞因子,可刺激炎症细胞合成和炎症介质释放,在气道炎症中发挥重要作用。据报道,支气管哮喘患者外周血TNF-α、IL-6、IL-1β水平升高,并且其水平越高,病情越严重[17]。过度炎症反应可诱导细胞凋亡。Caspase是一类与细胞凋亡密切相关的蛋白水解酶家族,Caspase活化是导致细胞凋亡的中心环节。作为Caspase级联反应的重要物质,Caspase-3可活化为Cleaved-Caspase-3,进而诱导细胞凋亡[18]。本研究结果显示,RSV 诱导后支气管上皮细胞培养上清液TNF-α、IL-6 和IL-1β 水平及细胞凋亡率、Cleaved-Caspase-3 表达均升高,表明RSV 能诱导支气管上皮细胞发生炎症反应,并促进其凋亡。本研究结果发现,RSV 感染可上调支气管上皮细胞中NFAM1表达;上调miR-660-3p表达可降低RSV 诱导的支气管上皮细胞培养上清液TNF-α、IL-6 和IL-1β 水平及细胞凋亡率、Cleaved-Caspase-3表达,提示上调miR-660-3p 表达可抑制RSV 诱导的支气管上皮细胞炎症反应和细胞凋亡。过表达NFAM1 则可使RSV 感染后支气管上皮细胞培养上清液TNF-α、IL-6、IL-1β 水平升高,促进细胞凋亡和上调Cleaved-Caspase-3 表达,提示过表达NFAM1 可促进支气管上皮细胞损伤,诱发并加重哮喘。
本研究通过在线生物信息学数据库对miR-660-3p的下游靶基因进行预测,结果发现miR-660-3p 与NFAM1的3′UTR存在互补结合位点,并通过双荧光素酶报告基因实验对miR-660-3p 与NFAM1 的靶向关系进行验证,结果证实二者存在靶向调控关系。本研究结果还发现,miR-660-3p 表达上调对支气管上皮细胞NFAM1 蛋白表达具有抑制作用,而下调miR-660-3p 表达则能促进NFAM1 蛋白表达,提示miR-660-3p 对NFAM1 具有靶向负调控作用。据此推测,miR-660-3p 可能通过靶向负调控NFAM1 表达参与RSV 诱导的支气管上皮细胞损伤。为了进一步验证miR-660-3p 的作用机制,本研究在上调miR-660-3p 表达的基础上通过pcDNA-NFAM1 质粒转染来过表达NFAM1,结果发现过表达NFAM1 可逆转miR-660-3p上调对RSV诱导的支气管上皮细胞凋亡和炎症因子分泌的抑制作用,提示miR-660-3p可能通过靶向负调控NFAM1 来抑制RSV 诱导的支气管上皮细胞凋亡和炎症因子分泌。
综上所述,miR-660-3p 可通过靶向负调控NFAM1 抑制RSV 诱导支气管上皮细胞凋亡及炎症因子分泌。
猜你喜欢
保健医苑(2022年1期)2022-08-30 08:40:04
保健医苑(2022年5期)2022-06-10 07:46:38
中国临床医学影像杂志(2021年6期)2021-08-14 02:21:56
基层中医药(2021年12期)2021-06-05 06:56:36
肝博士(2020年5期)2021-01-18 02:50:18
国际呼吸杂志(2019年22期)2019-12-09 09:20:26
国际呼吸杂志(2019年5期)2019-03-30 01:38:20
国际呼吸杂志(2019年3期)2019-03-01 05:39:06
国际呼吸杂志(2019年2期)2019-02-14 06:11:26
医学研究杂志(2015年7期)2015-06-22 11:01:01
