
井式节水灌溉下红肉苹果花芽分化期间内源激素的动态变化
2024-04-30 14:22:17程辉玲杨建军程平吴亚楠魏刚刚
江苏农业科学 2024年6期
关键词:花芽分化
程辉玲 杨建军 程平 吴亚楠 魏刚刚

摘要:为了解红肉苹果花芽各分化时期及特点,探明井式节水灌溉模式下该品种花芽分化期间内源激素的动态变化规律,利用X射线成像技术观察不同灌水梯度(W1、W2、W3处理)及漫灌(CK)下红肉苹果花芽内部结构,对其分化时期进行划分,并利用高效液相色谱法测定内源激素IAA、ABA、GA3、ZR的含量。结果表明,红肉苹果花芽分化开始时间早,持续时间长,各时期均有重叠,整个过程可分为未分化期、分化初期、萼片形成期、花瓣形成期、雄蕊形成期、雌蕊形成期等6个时期。不同灌水梯度下内源激素含量及比值变化趋势相同,总体上,IAA含量呈上升趋势,GA3含量一直维持较低水平,ZR含量呈先上升后下降的趋势,ABA含量呈上升态势,ABA/IAA、ABA/GA3呈逐渐上升的趋势,ZR/GA3、ZR/IAA呈先上升后下降的趋势,GA3/IAA、ZR/ABA呈下降趋势。W1、W2、W3处理完成花芽分化的时间略早于CK;高水平的ABA、ZR及较高的ABA/IAA、ABA/GA3、ZR/GA3、ZR/IAA能够促进红肉苹果花芽分化。
关键词:花芽分化;井式节水灌溉;内源激素;红肉苹果
中图分类号:S661.107;S661.01 文献标志码:A
文章编号:1002-1302(2024)06-0166-07
收稿日期:2023-05-11
基金项目:新疆维吾尔自治区林业发展补助资金(编号:新[2019]TG8号);新疆维吾尔自治区创新环境建设专项-科技创新基地建设项目(编号:PT2107)。
作者简介:程辉玲(1997—),女,新疆乌鲁木齐人,硕士研究生,主要从事果树花芽分化研究。E-mail:2291201424@qq.com。
通信作者:杨建军,博士,教授,博士生导师,主要从事矿山修复研究。E-mail:yjj@xju.edu.cn。
花芽分化是指果树每年形成的顶端分生组织由营养生长向生殖生长转化的过程,包括形态分化和生理分化,是内外因素共同作用、相互协调的结果[1]。在果树年生长周期中,花芽分化占据着重要的地位,对该过程的调控是高产高效栽培的关键环节之一[2]。在花芽分化过程中,水分是重要影响因素之一,适度干旱可以促进部分植物的花芽分化[3-5]。在水分调控研究方面,适当的水分控制能够为植物花芽分化创造条件[6-8],大多数果树可通过节水灌溉达到控制果树营养生长、增加营养积累、促进花芽分化的目的[9],如樱桃[10]、苹果[11]的花芽分化调控。此外,内源激素也会影响花芽分化,如马文春以长富2号为试验材料,利用外源葡萄糖调控花芽分化[12];刑利博通过喷施不同浓度的果树促控剂PBO溶液探究促进长富2号和富红早嘎花芽分化的生理机制[13]。花芽分化是苹果树丰产、稳产的关键,因此,研究其影响因素与调控措施对提高苹果树产量具有重要的实践意义。红肉苹果(Malus niedzwetzkyana Dieck)果实中富含花青苷和多酚等抗氧化物质,具有较高的营养价值[14],但由于红肉苹果育种周期长,缺乏优异种质,其产量一直较低,无法满足市场需求[15]。阿克苏地区是新疆的苹果主产区,红肉苹果是该地区近年来引进并积极推广的主要品种,但阿克苏地区苹果产业的发展长期受制于水资源条件,因此,研究节水灌溉等水分调控条件下苹果树的丰产、高产机制显得尤为重要。本试验以红肉苹果为试验材料,研究井式节水灌溉模式下红肉苹果花芽分化时期及激素的动态变化,通过设置不同的灌水梯度,以大田漫灌为对照,探讨红肉苹果的花芽分化进程,定量分析花芽分化期内源激素含量的动态变化,明晰花芽分化对灌水梯度的响应规律,以期揭示阿克苏地区红肉苹果花芽分化期水分调控机制,进而为红肉苹果的高产丰产研究提供理论依据。
1 材料与方法
1.1 研究区概况
试验地位于新疆阿克苏地区温宿县五团十三连,属典型的温带大陆性干旱气候,四季分明,春季升温快而不穩,夏季炎热干燥,秋季短暂而降温迅速;年日照时数2 747 h,年平均总辐射量 6 000 MJ/m2,平均海拔高度1 103 m;年降水量42.4~94.0 mm,年潜在蒸发量2 956.3 mm,年均气温10.1 ℃,无霜期195 d;浅层地下水位3.3 m;试验地土壤类型为沙壤土,土壤容重为1.44 g/cm3,田间持水率为19.6%。
1.2 试验材料与试验设计
试验地为八年生红肉苹果园,样树均为矮化密植栽培,砧木为平邑甜茶,平均株高约3.7 m,东西、南北平均冠幅分别约为2.2、2.7 m,果园常规管理为病虫害防治、杂草清除等。
试验采用单因子设计,设置3个灌溉水平,分别为井式节水灌溉模式下的低水处理(W1)、中水处理(W2)、高水处理(W3),对照组为大田漫灌(CK),每个处理28棵试验样树。井式节水灌溉模式是在同行每2棵树中间打坑安装高50 cm、半径为6 cm的竖井管(管身分布直径约2.5 mm的出水孔,密度为 2孔/cm2),用引流管将竖井管和地面的滴灌管连接,进行灌溉;同时利用与施肥罐(体积约250 L)连接的滴灌管将水溶肥溶化后输送至竖井管中,各处理肥料种类与用量一致。
灌水梯度根据试验地净灌水定额设置,由灌水时间控制,分别为W1处理:灌水6 h、W2处理:灌水 9 h、W3处理:灌水12 h,滴头流量为10 L/h(可通过调节压力大小控制流量),对照(CK)为常规漫灌,灌水量为每次1 250 kg/hm2。为避免水分侧渗,漫灌与井式灌溉间设置保护行,并沿保护行设置高约 20 cm、宽15 cm的土埂。W1、W2、W3处理年灌水总量(2022年3—10月,每月灌水1次)分别约为净灌水定额的2、3、4倍。
净灌水定额计算公式:
M=R×H×P(θmax-θmin)。
式中:M为净灌水定额,mm;R为土壤容重,g/cm3;H为土壤计划湿润层深度,取800 mm;P为湿润比,取0.4;θmax、θmin分别为土壤含水量(质量含水量)上限和下限,%。
1.3 采样与室内试验测定
1.3.1 花芽内部形态结构扫描
从2021年6月20日至10月8日,每隔10 d采取树冠外围短枝花芽20个,用FAA固定液固定后置于4 ℃冰箱保存备用。利用X射线显微成像系统扫描FAA固定液中的花芽并观察花芽形态分化时期,该系统采用独特的X光光学显微成像技术,通过不同角度的X射线透视图像,结合数字重构、渲染技术,提供花芽内部复杂结构数字图像,对花芽内部的微观结构进行微米尺度上的数字化表征,以便掌握红肉苹果花芽形态分化进程。
1.3.2 花芽内源激素含量测定
分别于2021年6月30日、7月30日、8月29日、9月28日及10月8日取红肉苹果树冠中上部外围结果母枝上的花芽,每次采集4个样本,共计20个样本,分装在冻存管后立即放入液氮罐中保存,用来测定花芽中内源激素含量。红肉苹果树花芽中内源激素含量利用高效液相色谱法测定,每个样本做3个平行重复,采用生长素(IAA)、赤霉素(GA3)、玉米素核苷(ZR)、脱落酸(ABA)作为标准样品(Sigma,USA),根据内部标准生成的校准曲线计算样品中IAA、GA3、ZR、ABA含量。
样品前处理:将适量待测样品迅速从液氮罐中取出后置于球磨仪(RETSCH-MM400)中,在频率为30 Hz条件下粉碎10 s,用Sartorius BP211D十万分之一电子天平(Sartorius公司)称取粉碎后的样品0.500 0 g(±0.000 5 g)置于15 mL离心管中,加入50%色谱级乙腈3 mL,用涡旋混合仪(IKA公司)混匀后,置于4 ℃冰箱浸提16 h。待测液准备:浸提后用冷冻离心机在10 000 r/min转速下离心10 min,取HBL柱分别用2 mL甲醇活化,再用2 mL纯水平衡后加入0.5 mL上清液,用抽真空机将样液抽至柱下24孔样品池中,再加入 2 mL 30%质谱级乙腈洗脱,将样品池中的洗脱液涡旋混匀,利用0.22 μm微孔滤膜过滤,吸入液相进样小瓶中待测。
1.4 数据处理
使用Excel 2019对试验数据进行统计分析;试验图片使用Origin 2021制作。
2 结果与分析
2.1 红肉苹果树花芽形态分化时期的观察及划分
通过观察发现,红肉苹果树花芽形态分化自6月中旬开始至10月初结束,约需要110 d。红肉苹果树花芽分化开始时间早,延续时间较长,外在形态略有差异。
红肉苹果树花芽自6月下旬正式进入分化初期,其中有极少部分还处于未分化期;7月中旬,仍有少数花芽处于分化初期甚至是未分化期;8月初至9月初,多数花芽处于萼片形成期或花瓣形成期;9月中旬,花芽分化逐渐进入雄蕊形成期;9月底至10月初,大多数花芽处于雌蕊形成期,分化基本完成。不同灌水梯度处理下红肉苹果进入分化初期的时间虽然相差较大,但在落叶前雌蕊分化均已完成。根据对红肉苹果花芽的扫描观察,将该品种的花芽形态分化划分为6个时期,如图1所示,分别是未分化期、分化初期、萼片形成期、花瓣形成期、雄蕊形成期、雌蕊形成期。
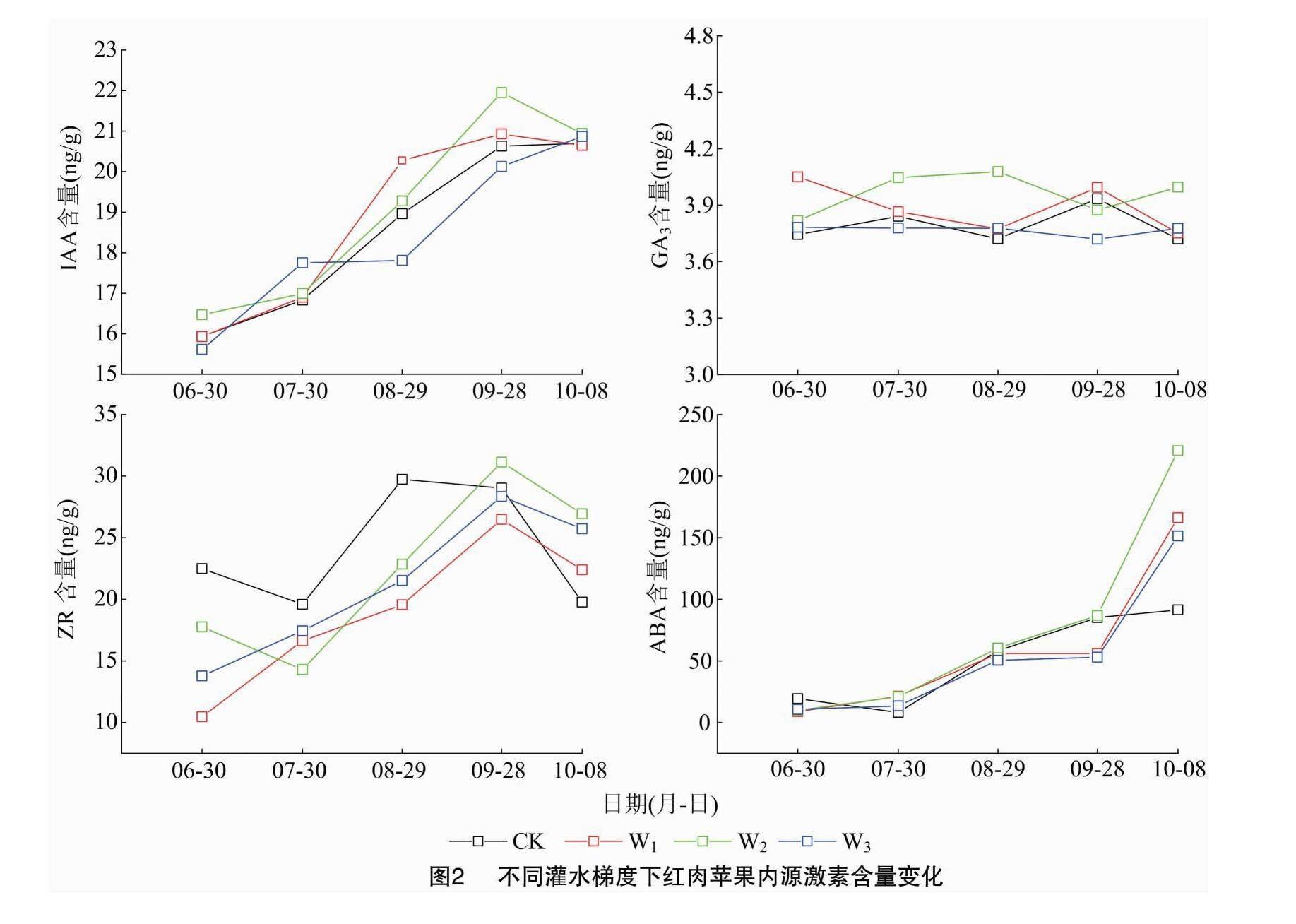
2.2 不同灌水梯度下红肉苹果树花芽内源激素含量的变化特征
由图2可知,在花芽分化的整个时期,IAA含量总体呈缓慢上升趋势,不同灌水梯度下红肉苹果花芽中的IAA含量水平变化相似。CK处理IAA含量变化范围在15.93~20.70 ng/g之间;W1处理IAA含量变化范围在15.93~20.64 ng/g之间,W2处理IAA含量变化范围在16.47~21.95 ng/g之间,W3处理IAA含量变化范围在15.61~20.87 ng/g之间。由此可知,6月下旬红肉苹果花芽开始分化时,CK、W1、W2处理IAA含量最低,之后缓慢上升,至10月时下降,而此时花芽形态分化已基本完成。
在花芽过程中ABA含量总体呈上升的趋势。CK处理ABA含量变化范围在19.29~91.46 ng/g之间,W1处理ABA含量变化范围在8.76~166.27 ng/g 之間,W2处理ABA含量变化范围在9.86~220.66 ng/g之间,W3处理ABA含量变化范围在10.69~151.34 ng/g之间。可见,红肉苹果花芽在分化过程中会出现ABA含量累积的情况。ABA能够抑制淀粉酶的活性,从而使淀粉积累,而高淀粉含量与花芽的分化密切相关。
CK、W1、W2、W3处理的GA3含量均在3.6~4.2 ng/g 之间,该激素含量在整个花芽形态分化期一直处于较低水平,且无较大波动出现,保持在稳定水平。
ZR含量在整个花芽形态分化期总体呈先上升后下降的趋势,其中W1、W2、W3处理均在9月底达到最高水平,分别为26.49、31.14、28.35 ng/g,此时3种节水处理花芽大多数已进入雌蕊分化期;CK处理的ZR含量则是在8月底达到峰值,为 29.74 ng/g,此时漫灌条件下花芽分化大多进入雄蕊分化期。
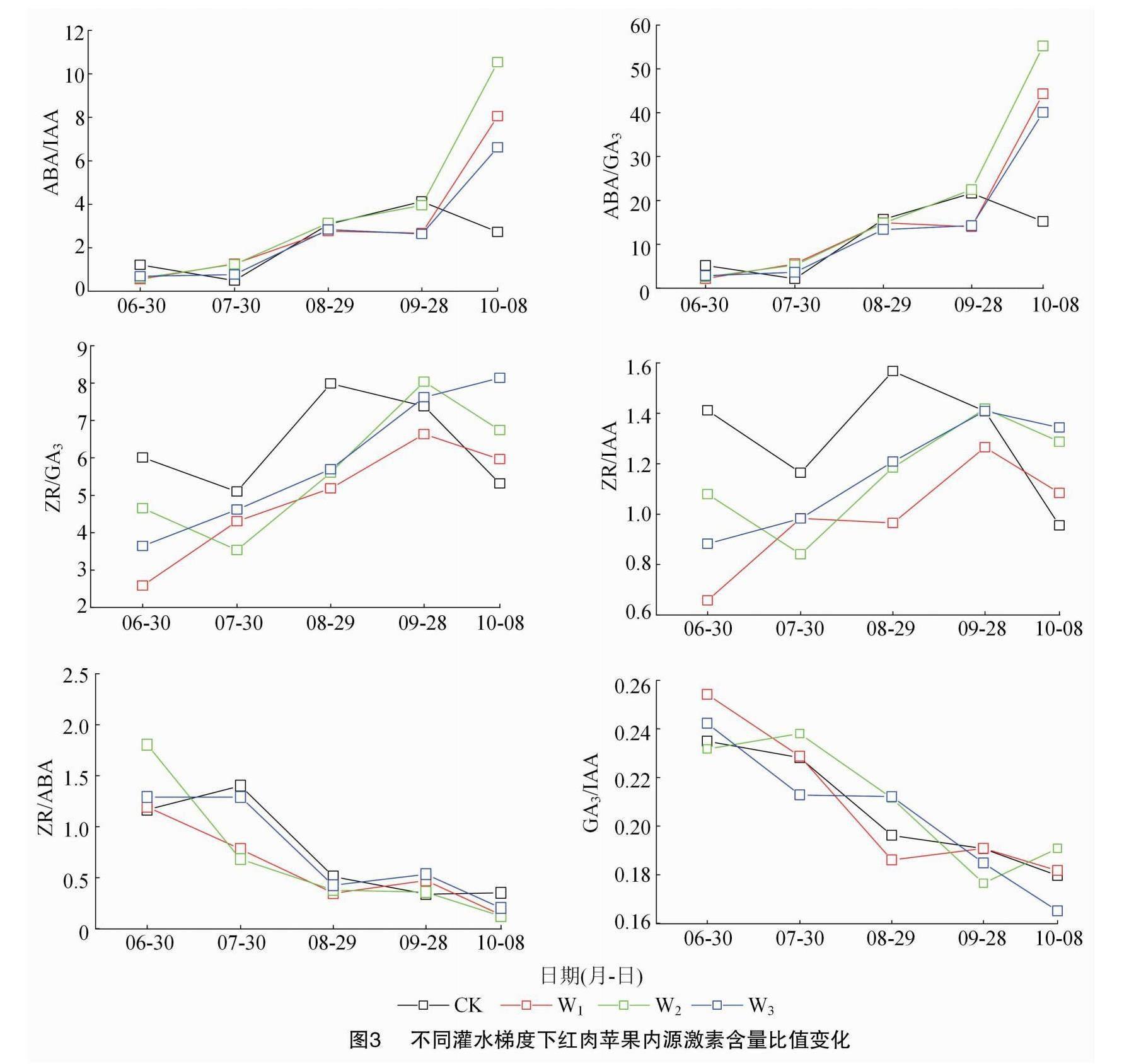
2.3 不同灌水梯度下红肉苹果树花芽内源激素含量比值的变化特征
由图3可知,ABA/IAA、ABA/GA3总体呈逐渐上升的趋势,且波动变化相似,结合图2可知,各处
理下的红肉苹果树花芽于6月30日已经进入分化初期,该阶段下的IAA、GA3、ABA含量水平都不高,但ABA含量要高于其他2种激素;随着花芽的不断分化,IAA、ABA含量逐渐上升,GA3含量变化一直维持在某一区间,波动较小,且ABA含量始终大于IAA与GA3含量。10月之后,2组比值上升,此时的花芽已基本完成分化。
ZR/GA3、ZR/IAA总体呈先上升后下降的趋势。由图2可知,ZR含量虽有波动,但一直处于较高水平,而由图3可知,ZR/GA3、ZR/IAA在7—9月上升较快,该阶段是绝大多数花芽的分化盛期。
GA3/IAA总体呈下降态势,在6月时最高,在之后的分化阶段中该比值逐渐下降,至10月时变化趋于平缓。ZR/ABA总体呈下降趋势。由图2可知,ZR与ABA在分化期都有所积累,但ABA含量水平要高于ZR。在图3中,ZR/ABA在9月时有略微提高,此时大部分花芽处于雌蕊分化期,10月之后略微下降或趋于稳定。
以上结果表明,在花芽形态分化期,各激素含量的比值高低影响着花芽的分化进程。同时也表明红肉苹果成花诱导期需要较高的ABA/IAA、ABA/GA3、ZR/IAA及ZR/GA3。
3 讨论
3.1 灌水梯度对花芽分化的影响
花芽分化是一个复杂的过程。首先是花芽的生理分化期,该时期决定着花芽的数量,在外界条件和内部激素适宜时,新梢上的芽生长点在生理上发生改变,逐渐向花芽进化,这一阶段是芽本身生长发育的重要时期;之后花芽进入形态分化期,生长点开始分化出萼片、花瓣、雌蕊与雄蕊等花器官,并在落叶前结束形态分化[16]。本研究通过X射线显微成像技术从显微结构上观察花芽内部形态的变化过程,可以为观察及划分花芽内部形态变化提供依据。水分是影响花芽分化的重要外界因素之一,花芽分化过程中细胞的分裂和膨大离不开水分的作用,同时,水分还可以通过影响花芽内部成分变化,如矿质营养、内源激素水平等间接影响花芽分化[17]。本研究中,不同灌水梯度下花芽分化时期在分化过程中有重叠现象,适度控制水分有利于红肉苹果花芽的分化。朱牧笛等在对苹果花芽形态分化特性的研究中发现,在花芽分化的过程中控制水分,能够抑制新梢的生长,从而有利于营养物质的积累,促使花芽的分化[18]。这是本研究中不同灌
水梯度处理下红肉苹果花芽分化持续时间及分化完成时间有所不同的原因之一。
3.2 内源激素含量变化与花芽分化的联系
植物激素指植物体内产生的一些微量而能调节(促进、抑制)自身生理过程的有机化合物,也被称为内源激素。花芽分化会受内源激素的调控,其中生长素(IAA)的作用比较复杂,低浓度起促进作用而高浓度起抑制作用;脱落酸(ABA)可以促进果树的花芽分化;赤霉素(GA)可抑制多种果树的花芽分化;细胞分裂素(CTK)的主要功能是促进细胞分裂,促进芽的分化是它最重要的生理效应之一[19]。
目前,激素平衡学说在花芽分化研究中颇受关注,不同激素对花芽分化的影响已基本被认知[20]。内源激素在植物中含量较低,且不同采样地域、采样时间之间可能存在差异,导致所研究的结果也存在差异。生长素(IAA)是一种极性分子,它对花芽分化的作用存在很大争议。李天红等认为,高浓度的IAA促进了富士苹果花芽的孕育[21];但李有梅等的研究证明,IAA通过促进富士苹果树体和枝条的旺长,使短枝比例减少,从而抑制成花[22]。在本研究中,6月30日各处理下的花芽已完成生理分化进入形态分化期,该阶段IAA含量处于最低水平,说明低浓度的IAA促进了红肉苹果成花。由此可见,IAA的作用效果因品种而异,有效作用浓度也有待进一步研究。
关于ABA的作用,曾骧认为,该激素具有双重性,一方面能使枝条停止生长,从而累积营养物质,以达到促使作物成花的目的;另一方面会诱导生长点休眠,导致作物无法成花[23]。在本研究中,不同灌水处理下的花芽在整个分化过程中ABA含量始终保持较高水平,表明ABA的累积能够促进红肉苹果花芽的分化。这与其他学者在长富2号苹果[24]、风味玫瑰[25]和李光杏[26]等杏李、大樱桃[27]等研究中得出的结论相同。但另有學者在对梨[28]、枇杷[29]进行研究时发现,高浓度的ABA会对其花芽分化产生抑制作用。综上所述,不同蔷薇科植物的花芽分化所需的ABA含量存在差异,因此,ABA对该科植物花芽分化的影响可能是多种因素共同决定的,高浓度ABA促进红肉苹果花芽分化的原因还有待进一步探讨。
GA3在植物成花过程中具有重要作用,对多数木本植物来说是主要的成花抑制激素。曹尚银等通过对首红苹果的研究发现,GA类物质在花芽形态分化期时开始下降,之后一直保持稳定水平[30]。马玲研究富士苹果时发现,喷施GA3后,树体花芽分化明显受到抑制,使得成花率降低[31]。本研究结果表明,红肉苹果在6月30日花芽形态分化开始后,花芽中的GA3含量水平一直较低,且无大幅度的变化,说明低浓度的GA3能促使红肉苹果花芽分化。同时,在大樱桃[27]、桂花[32]、桃花[33]、油桐[34]等作物上得到同样的结论。
细胞分裂素(CTK)是一种能够促进细胞分裂与膨大、打破顶端优势,并诱导作物成花的激素。玉米素核苷(ZR)是CTK在木质部中运输的主要形式。张宁发现,高水平的ZR能促进文冠果花芽分化[35]。徐石兰等也发现,高含量的细胞分类素能促进龙眼成花[36]。在本试验中,红肉苹果中的ZR含量在整个分化期处于较高水平,说明高浓度ZR能够促进红肉苹果花芽分化,在毛棉杜鹃[37]、勒杜鹃[38]、油橄榄[39-41]及板栗[42]的研究中具有类似结果。
3.3 内源激素含量比值变化与花芽分化的联系
激素可以调节植物生命活动周期的各个阶段,在果树花芽分化阶段受到的影响更加明显。有研究者认为,促花物质与抑花物质之间的平衡关系,即激素含量比值平衡,会在很大程度上决定果树成花[43]。在一定的营养水平下,内源激素的平衡对成花起主导作用:赤霉素可提高淀粉酶活性促进淀粉水解,而脱落酸对赤霉素有拮抗作用,有利于淀粉的积累;在夏季对果树新梢进行摘心,会使赤霉素和生长素含量减少,细胞分裂素含量增加,这样能改善营养物质的分配,促进花芽分化[19]。综上,当植物体内淀粉、蛋白质等营养物质丰富,细胞分裂素和脱落酸含量较高时,有利于花芽分化。
内源激素对花芽分化的作用并不取决于单独的某一种激素,而是通过激素之间的相互作用达到某种动态平衡,即通过不同内源激素含量的比值变化来实现的。通过对不同木本植物花芽分化过程中不同激素比值的研究发现,ABA/IAA、ABA/GAs、ZR/GAs和ZR/IAA等比值高有助于花芽分化的进行[44]。
曹尚银等研究发现,苹果在ABA/IAA、ABA/GA3、ZR/IAA、ZR/GA3提高后,花芽分化率有所提升,表明ABA/IAA、ABA/GA3、ZR/IAA、ZR/GA3的提高有利于苹果成花[45]。在花芽分化期,刺梨花芽的ABA/IAA、ABA/GA、ZR/GA、ZR/IAA高于叶芽,这能够促进刺梨的花芽分化[46]。鄧灿在关于枇杷的相关试验中得出相似结论[47]。由此可见,单一某种激素对花芽分化产生的影响不大,多种内源激素可以通过相互间的平衡状态来调控花芽分化。红肉苹果花芽形态分化期ZR/IAA、ZR/GA3、ABA/IAA、ABA/GA3较高,说明该品种需要一定的内源激素含量比值促进花芽分化。
本研究对不同灌水梯度下红肉苹果花芽形态分化的变化特征以及花芽分化过程中内源激素的动态变化进行分析,但花芽分化的一系列过程非常复杂,其他因素(光照、温度及营养物质等)对红肉苹果花芽分化过程的影响尚不明确,有关灌水梯度对红肉苹果花芽分化的影响还有待探讨。
4 结论
不同灌水梯度下红肉苹果花芽分化存在以下特点:(1)井式节水灌溉处理下红肉苹果花芽分化持续时间长,且各时期有所重叠,分化过程分为未分化期、分化初期、萼片形成期、花瓣形成期、雄蕊形成期、雌蕊形成期等6个时期;(2)在红肉苹果花芽分化期内两大促花激素ABA、ZR含量的升高能促使红肉苹果花芽分化;(3)ZR/IAA、ZR/GA3、ABA/IAA、ABA/GA3的升高能促进红肉苹果花芽形态分化。
参考文献:
[1]王文,张宜仓,宋建平. 影响苹果花芽分化的原因及调节的途径[J]. 现代农业科学,2009,16(6):21-22.
[2]辛明志,陶炼,樊胜,等. 纬度和海拔对主要苹果品种花芽分化期的影响[J]. 园艺学报,2019,46(4):761-774.
[3]路瑶. 水分胁迫、夏季修剪对葡萄花芽分化的影响研究[D]. 长沙:湖南农业大学,2012.
[4]万春雁,糜林,李金凤,等. 苗期不同水分处理对草莓花芽分化及果实早熟化的影响[J]. 果树学报,2016,33(12):1523-1531.
[5]胡智勇. 土壤水分含量对龙眼成花的影响[D]. 福州:福建农林大学,2005.
[6]谢利娟,麦瑞娟,孙敏. 影响植物花芽分化的内部因素研究进展[J]. 安徽农业科学,2009,37(30):14585-14588,14631.
[7]李烨. 影响苹果花芽分化的因素和促进措施[J]. 果农之友,2009(6):38,40.
[8]杨盛,白牡丹,郭黄萍. 环境因子与花芽分化关系研究进展[J]. 内蒙古农业大学学报(自然科学版),2018,39(5):97-100.
[9]杨盛,白牡丹,高鹏,等. 果树花芽分化期间水分管理技术[J]. 中国农业信息,2017(12):70-73.
[10]王光森,党爱民,于吉祥. 小管出流技术在樱桃园灌溉中的应用[J]. 山东水利,2018(7):48-49.
[11]王晓华,高大鹏. 辽西北地区寒富苹果的需水特点与节水灌溉技术[J]. 现代农村科技,2013(7):38.
[12]马文春. 外源葡萄糖对苹果花芽分化的影响及候选基因MdSC35_2功能分析[D]. 杨凌:西北农林科技大学,2020.
[13]邢利博. PBO促进富士苹果花芽分化的生理机制[D]. 杨凌:西北农林科技大学,2013.
[14]冀晓昊. 红肉苹果愈伤组织诱导及红色性状发育机理研究[D]. 泰安:山东农业大学,2015.
[15]陈学森,王楠,张宗营,等. 我国高类黄酮(红皮与红肉)苹果育种取得突破性进展[J]. 中国果树,2020(2):6-9,22,141.
[16]Mert C,Barut E,I·pek A. Variation in flower bud differentiation and progression of floral organs with respect to crop load in olive[J]. Notulae Botanicae,Horti Agrobotanici,Cluj-Napoca,2013,41(1):79-85.
[17]Li-Mallet A,Rabot A,Geny L. Factors controlling inflorescence primordia formation of grapevine:their role in latent bud fruitfulness?A review[J]. Botany,2016,94(3):147-163.
[18]朱牧笛,張荣,石卓功.苹果花芽形态分化特性研究[J]. 安徽农业科学,2017,45(4):42-44.
[19]潘瑞炽.植物生理学[M]. 4版.北京:高等教育出版社,2001:314-335.
[20]Zuo X Y,Zhang D,Wang S X,et al. Expression of genes in the potential regulatory pathways controlling alternate bearing in ‘Fuji (Malus domestica Borkh.) apple trees during flower induction[J]. Plant Physiology and Biochemistry,2018,132:579-589.
[21]李天红,黄卫东,孟昭清. 苹果花芽孕育机理的探讨[J]. 植物生理学报,1996,22(3):251-257.
[22]李有梅,邢利博,张东,等. 喷施IAA抑制富士幼树花芽孕育的机制[J]. 西北农业学报,2015,24(4):84-89.
[23]曾骧. 果树生理学[M]. 北京:中国农业大学出版社,1992:158-159.
[24]苏静. ABA及其抑制剂钨酸钠对‘长富2号花芽分化的影响[D]. 兰州:甘肃农业大学,2017.
[25]莫文娟,张嘉嘉,杨绍彬,等. 杂交杏李‘风味玫瑰花芽分化过程及内源激素动态变化[J]. 中国农业大学学报,2020,25(11):54-61.
[26]赵通. 敦煌‘李光杏花芽分化形态观察及雌蕊败育的转录组学分析[D]. 兰州:甘肃农业大学,2020.
[27]王玉华,范崇辉,沈向,等. 大樱桃花芽分化期内源激素含量的变化[J]. 西北农业学报,2002,11(1):64-67.
[28]Liu Y,Zhang H P,Gu C,et al. Transcriptome profiling reveals differentially expressed genes associated with wizened flower bud formation in Chinese pear (Pyrus bretschneideri Rehd.)[J]. The Journal of Horticultural Science and Biotechnology,2016,91(3):227-235.
[29]刘宗莉,林顺权,陈厚彬. 枇杷花芽和营养芽形成过程中内源激素的变化[J]. 园艺学报,2007,34(2):339-344.
[30]曹尚银,张俊昌,魏立华. 苹果花芽孕育过程中内源激素的变化[J]. 果树科学,2000,17(4):244-248.
[31]马玲. 喷施GA3和6-BA对“富士”苹果顶芽内源激素、枝组构成及成花结果的影响[D]. 杨凌:西北农林科技大学,2018.
[32]陆婷,王艺光,骆义波,等. ‘四季桂不同季节的花芽分化与发育比较[J]. 园艺学报,2017,44(6):1145-1156.
[33]安丽君,金亮,杨春琴,等. 外源赤霉素对桃的成花效应及其作用机制[J]. 中国农业科学,2009,42(2):605-611.
[34]郭文丹,李建安,刘丽娜,等. 油桐花芽分化期内源激素含量的变化[J]. 经济林研究,2009,27(2):31-34.
[35]张宁. 文冠果花芽分化及与内源激素关系研究[D]. 北京:北京林业大学,2019.
[36]徐石兰. 龙眼不同时间修剪对树体内源激素、酶活性及生长结果的影响[D]. 南宁:广西大学,2019.
[37]谢利娟,孙敏,赵梁军,等. 毛棉杜鹃芽形态分化期间封顶叶内源激素含量变化的研究[J]. 中国农业大学学报,2010,15(4):33-38.
[38]杨义标,邵志芳,邱少松,等. 勒杜鹃花芽分化过程中内源激素含量和碳氮营养的变化研究[J]. 中国农学通报,2008,24(2):281-284.
[39]贺娜,马婷,徐田. 油橄榄花芽分化期叶片内含物变化规律研究[J]. 林业调查规划,2016,41(1):28-30,43.
[40]杜晋城,李丕军,辜云杰,等. 油橄榄花芽分化期内源激素及碳水化合物含量的变化研究[J]. 西部林業科学,2018,47(1):122-126.
[41]朱振家,姜成英,史艳虎,等. 油橄榄成花诱导与花芽分化期间侧芽内源激素含量变化[J]. 林业科学,2015,51(11):32-39.
[42]张立民,李凤鸣,苏淑钗,等. 板栗二次花芽分化过程中激素含量变化[J]. 经济林研究,2009,27(4):31-35.
[43]Luckwill L C. A new look at the process of fruit bud formation in apple[C]//Proceedings 19th International Horticultural Congress. Warszawa,1974,3:237-245.
[44]宋杨,窦连登,张红军. 蓝莓不同品种花芽形成过程中内源激素的变化[J]. 中国南方果树,2014,43(5):106-108,114.
[45]曹尚银,汤一卒,张俊昌. GA3和PP333对苹果花芽形态建成及其内源激素比例变化的影响[J]. 果树学报,2001,18(6):313-316.
[46]樊卫国,刘国琴,安华明,等. 刺梨花芽分化期芽中内源激素和碳、氮营养的含量动态[J]. 果树学报,2003,20(1):40-43.
[47]邓灿. ‘早钟6号枇杷夏梢花芽分化期生理机制研究[D]. 雅安:四川农业大学,2016.
猜你喜欢
黑龙江粮食(2022年6期)2022-11-23 12:51:31
青岛农业大学学报(自然科学版)(2021年2期)2021-07-07 09:46:16
果农之友(2020年5期)2020-06-21 15:11:28
落叶果树(2020年3期)2020-06-17 05:11:28
河北果树(2020年1期)2020-02-09 12:31:08
农业与技术(2019年5期)2019-05-29 11:08:08
农民致富之友(2018年21期)2018-12-27 03:49:52
现代园艺(2018年16期)2018-08-31 07:29:14
果农之友(2017年6期)2017-06-29 07:51:25
广东农业科学(2016年3期)2016-07-27 02:07:13
