
菠萝AcNINV家族全基因组分离及表达分析
2024-04-30 04:30吴建阳陈妹姚艳丽张秀梅
果树学报 2024年4期
吴建阳 陈妹 姚艳丽 张秀梅

摘 要:【目的】鑒定出菠萝NINV家族全基因组成员,初步阐明NINV基因与蔗糖代谢的关系。【方法】采用生物信息学方法鉴定并分析菠萝NINV家族全基因组成员,通过实时荧光定量PCR分析其表达特性,利用HPLC对蔗糖含量进行测定。【结果】在菠萝中共鉴定出6个NINV基因,分布于5条不同染色体上,其二级结构主要由α-螺旋和无规则卷曲组成,该基因家族启动子区域光反应元件数量最多。AcNINV外显子数量介于4~6个之间,其中AcNINV3、6外显子的数量为6个,亚细胞定位预测发现这2个基因都分布于叶绿体,属于α亚族;AcNINV1、2、4、5外显子的数量为4个,亚细胞定位预测发现这4个基因都分布于质膜,属于β亚族。在果柄、果皮、果心中表达量最高的是AcNINV2基因,在果肉中表达量最高的是AcNINV6基因。蔗糖含量随着菠萝果实成熟呈先升高后降低的变化趋势,而AcNINV4基因在菠萝果实成熟过程中呈下调表达。【结论】AcNINV4可能是催化果实蔗糖降解的水解酶基因。
关键词:菠萝(Ananas comosus);蔗糖含量;碱性/中性转化酶;基因家族;基因表达
中图分类号:S668.3 文献标志码:A 文章编号:1009-9980(2024)04-0598-13
Genome-wide identification and expression analysis of AcNINV family in pineapple
WU Jianyang1, 2, CHEN Mei3, 4, YAO Yanli3, 4, ZHANG Xiumei3, 4*
(1Zhanjiang Preschool Education College, Zhanjiang 524037, Guangdong, China; 2Basic Education College of Lingnan Normal University, Zhanjiang 524037, Guangdong, China; 3South Subtropical Crop Research Institute, Chinese Academy of Tropical Agricultural Science (CATAS)/Key Laboratory of Tropical Fruit Biology, Ministry of Agriculture and Rural Affairs, Zhanjiang 524091, Guangdong, China; 4National Key Laboratory for Tropical Crop Breeding, Sanya 572024, Hainan, China)
Abstract: 【Objective】 Sucrose plays a crucial role in plant growth and development. The alkaline/neutral invertase (NINV) proteins irreversibly cleave sucrose into fructose and glucose. To date, the genome-wide identification, characterization, and expression profile analysis of the NINV gene families have been reported in many species but not in pineapple. 【Methods】 Genome sequence and annotation data were acquired from the pineapple genome database through the blastp against protein database and the tblastn against genome databases using the query sequences of Arabidopsis thaliana NINV proteins to identify the potential members of the NINV gene families in pineapple. The physicochemical characteristics, including molecular weight (MW), theoretical isoelectric points (pI), and instability index were obtained using the ExPASy tool. The CDS sequence and protein length were acquired from the pineapple genome database. The synteny analysis between different species and exon/intron structures were visualized using the TBtools. The conserved motifs of the AcNINV gene families were analyzed using the online MEME tool. The phylogenetic tree was constructed using the MEGA 5.0 through the neighbor-joining approach. The total RNA was isolated in accordance with the method described by Wu et al. RT-qPCR was performed using the LightCycler480 Ⅱ System (Roche, Switzerland) and the DyNAmo Flash SYBR Green qPCR kit (Thermo, USA). RT-qPCR was performed in a total volume of 10 μL, which contained 1 μL diluted cDNAs and 5 ?L SYBR Green PCR Master Mix, and the primer final concentration was 250 nmol·L-1. The sucrose content was determined by HPLC. 【Results】 In this study, six NINV genes (AcNINV1-6) were identified. The six AcNINV genes were distributed on five different chromosomes (LG1, LG3, LG14, LG17, LG21). There were two genes on LG21: AcNINV5 and AcNINV6, while another AcNINV gene on the other chromosomes. AcNINV proteins had a length of 556-673 aa and MW between 63.03 and 74.80 ku. Only one collinear NINV gene pair between pineapple and A. thaliana, and eight collinear NINV gene pairs between pineapple and Oryza sativa were identified. The number of exons in the AcNINV ranged from 4 to 6, and the number of introns ranged from 3 to 5. The AcNINV1, 2, 4, 5 possessed 4 exons and 3 introns, the AcNINV3 and AcNINV6 had 6 exons and 5 introns. A total of 10 conserved motifs were predicted in the AcNINV genes. Among them, the AcNINV1, 2, 4, 5 contained 10 conserved motifs, but the AcNINV3, 6 only had 9 conserved motifs and lacked motif10. A phylogenetic tree was constructed by combining six AcNINV genes from pineapple with the amino acid sequences of the NINV gene family from three species, including Arabidopsis, rice, and cassava. Six AcNINV genes were clustered into two subfamilies, i.e., α and β. The AcNINV1, 2, 4, 5 belonged to clade β, the AcNINV3 and AcNINV6 belonged to clade α. The promoter cis-elements of the AcNINVs were examined using the Plant-CARE database. There were many cis-elements involved in light (MRE, ATC-motif, Box 4, L-box, GATA-motif, G-box, GT1-motif, TCCC-motif, 3-AF1 binding site, LAMP-element, Sp1, TCT-motif, GA-motif, chs-CMA1a, AT1-motif, AE-box), stress (TC-rich repeats, LTR, ARE, GC-motif) and hormones (ABRE, TGA-element, AuxRR-core, P-box, O2-site, TATC-box, TCA-element, TCA-element, CGTCA-motif, TGACG-motif) in the promoter region. The number of light responsive elements were the largest group, followed by the hormone responsive elements, stress responsive elements. The AcNINV5 had the most cis-elements in light response and hormone response. The AcNINV1 had the most cis-elements in stress response. The sucrose content increased first and then decreased. The sucrose content remained constant on the 20th-40th days after anthesis (DAA). Afterwards, the sucrose content extensively increased from 40 DAA to 80 DAA and reached the highest value on the 80 DAA, which was 126-fold higher than that at 20 DAA. The sucrose content from 80th DAA to 100th DAA extensively decreased but was still 12-fold higher than that on 20th DAA. The expression of the six AcNINV genes was studied using RT-qPCR during fruit development in pineapple. The expression was significant different among the different AcNINV gene members. The expression of the six AcNINV genes were presented three types. The first type was upregulated, such as the AcNINV2, 3, 6. The second type was downregulated first and then upregulate, but the amplitude of change was not significant, the AcNINV1, 5 belonged to this type. The third type was downregulated, and the AcNINV4 gene belonged to this type. The expression level of the AcNINV4 gene remained almost unchanged on the 20th-40th DAA, but it rapidly decreased from the 40th DAA to the 100th DAA. On the 20th DAA, the AcNINV4 gene expression was five times more than that of the 100th DAA. Meanwhile, RT-qPCR was used to clarify the expression profile of the AcNINV gene families in the different tissues. The AcNINV1 had the highest expression in the peduncle and the lowest expression in the core. The expression level of the AcNINV2, AcNINV3 and AcNINV6 were highest in the pericarp and lowest in the flesh. The expression level of the AcNINV4 and AcNINV5 was the highest in the peduncle and lowest in the flesh. Furthermore, the AcNINV2 gene had the highest expression in the peduncle, pericarp, and core. While the AcNINV6 gene had the highest expression level in the flesh. 【Conclusion】 A total of six NINV genes (AcNINV1-6) were identified and they were distributed on 5 chromosomes in pineapple. The sucrose content increased with the ripening of pineapple fruit, and the AcNINV4 gene was downregulated during the ripening process of pineapple fruit. Therefore, it was speculated that AcNINV4 may be a hydrolytic enzyme gene that may catalyze the degradation of fruit sucrose.
Key words: Pineapple (Ananas comosus); Sucrose content; Alkaline/neutral invertase; Gene family; Gene expression
菠萝(Ananas comosus)是凤梨科的草本植物[1],是我国重要的热带水果,中国是世界十大菠萝生产国之一。在中国种植菠萝的主要省份包括广东、广西、海南、云南和福建。2018年,中国菠萝产量为162万t[2]。
蔗糖是光合作用的主要产物[3],蔗糖的含量直接影响果实的品质和风味。蔗糖转化酶是水解蛋白酶,定向水解蔗糖生成果糖和葡萄糖[4],依据其在植物细胞内的位置可分为胞质转化酶、细胞壁转化酶和液泡转化酶;依据其最佳pH 值的不同,液泡转化酶和细胞壁转化酶又被称为酸性转化酶(acid invertase,AIN),胞质转化酶则被称为碱性/中性转化酶(alkaline/neutral invertase,NINV)[5]。AIN不仅可以水解蔗糖,还能水解其他含β-果糖的寡糖,比如水苏糖、核糖,但NINV却专一性地催化蔗糖[4]。由于NINV不稳定、易失活,因此对它的研究报道不多,生理功能知之甚少[6],但近些年来,随着生物技术的快速发展,关于NINV的研究越来越多。
NINV在植物生长发育中具有至关重要的作用,特别是在生殖器和根系的发育中;在成熟组织中,由于AIN活性低,因此NINV对蔗糖的分解更为关键[7]。在杏果实发育过程中NINV活性与蔗糖的积累呈显著负相关[8],在草莓果实发育过程中也发现相同的结论[9]。另外,NINV通过分解蔗糖,激活糖信号通路和改变渗透势进而参与植物胁迫防御[10]。
NINV基因首次从大豆下胚轴中分离获得[11],NINV由多基因家族编码,辣椒中有7个成员[12],水稻[13]、茶树[10]、柑橘[14]中有8个成员,拟南芥[15]和铁皮石斛[16]中有9个成员,木薯中有11个成员[17],苹果中有12个成员[18],毛果杨中有16个成员[19],但在菠萝中至今还未有NINV基因家族的报道。笔者借助于菠萝基因组数据库,首先全面鉴定出NINV基因家族成员,再分析不同成员在菠萝果实成熟过程中的表达特性,以此初步阐明NINV基因与蔗糖代谢的关系。本研究结果丰富了NINV基因在调控菠萝蔗糖中的研究内容,并将为利用分子生物学手段培育菠萝新品种提供重要理论支撑。
1 材料和方法
1.1 植物材料
菠萝(Ananas comosus ‘Comte de paris)于2019年1—4月采摘于中国热带农业科学院南亚热带作物研究所菠萝种资资源库。共选择150株长势一致的菠萝植株进行挂牌,从谢花后(days after anthesis,DAA)20 d到果实成熟期间每10 d随机采取9个大小一致的果实。将9个果实分为3组,每组为1个生物学重复,分离后的果肉用于基因表达分析。另外,选取9株长势一致的成熟菠萝进行组织特异性表达分析。样品在田间收获后立即带回实验室,并进行果肉、叶基、叶柄、果皮、果心的分离,分离的组织放入液氮速冻后存放于-80 ℃冰箱。
1.2 菠萝AcNINV基因家族的鉴定
从TAIR数据库(http://www.arabidopsis.org/index.jsp)中下载拟南芥所有NINV基因序列,利用拟南芥NINV基因序列在菠萝基因组(http://pineapple.angiosperms.org/pineapple/html/index.html)[20]中进行Blast序列比对,再利用SMART在线软件(http://smart.embl-heidelberg.de/)对所有序列进行验证,从而获得菠萝AcNINV基因家族所有成员序列。
1.3 序列分析
利用ExPASy工具(https://web.expasy.org/protparam/)获得AcNINV基因家族成员分子质量(MW)、理论pI、亲水性总平均值(GRAVY)和不稳定指数,从菠萝基因组数据库中获得CDS序列、蛋白质长度等指标,利用TBtools[21]绘制AcNINV基因家族在染色体上的定位和分布图。AcNINV蛋白序列的二级结构利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)在线软件进行分析。基因亚细胞定位预测利用CELLO:Subcellular Localization Predictive System(http://cello.life.nctu.edu.tw/)在线软件进行分析。利用TBtools[21]进行不同物种间AcNINV基因共线性分析。
1.4 基因结构和保守基序分析
外显子/内含子结构利用TBtools可视化分析[21]。利用MEME在线工具(http://meme-suite.org/tools/meme)和TBtools对AcNINV基因家族的保守基序进行分析和可视化,最大基序数设置为10。
1.5 菠萝AcNINV基因系统发育分析
使用ClustalX和MEGA-5.0采用鄰接(NJ)方法构建AcNINV基因与其他物种NINV基因系统进化树。
1.6 启动子顺式作用元件分析
利用TBtools提取起始密码子(ATG)上游2000 bp的基因序列,然后利用PlantCARE在线软件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测各个基因的顺式作用元件,再利用TBtools绘制顺式作用元件图。
1.7 蔗糖含量的测量
根据Zhang等[22]的方法采用高效液相色谱法(HPLC)提取并测定蔗糖含量。反应条件为:柱温35 ℃,流动相为乙腈和蒸馏水,体积比7∶3,流速1 mL·min-1。
1.8 RNA提取、质量控制和cDNA合成
采用Wu等[23]的方法分离总RNA,用DNaseⅠ(TaKaRa,Otsu,Japan)去除DNA污染。安捷伦2100生物分析仪(Agilent Technologies,Santa Clara,CA,USA)用于检测RNA的数量和质量。用M-MLV cDNA合成试剂盒(Promega)合成第一链cDNA。
1.9 实时荧光定量PCR分析
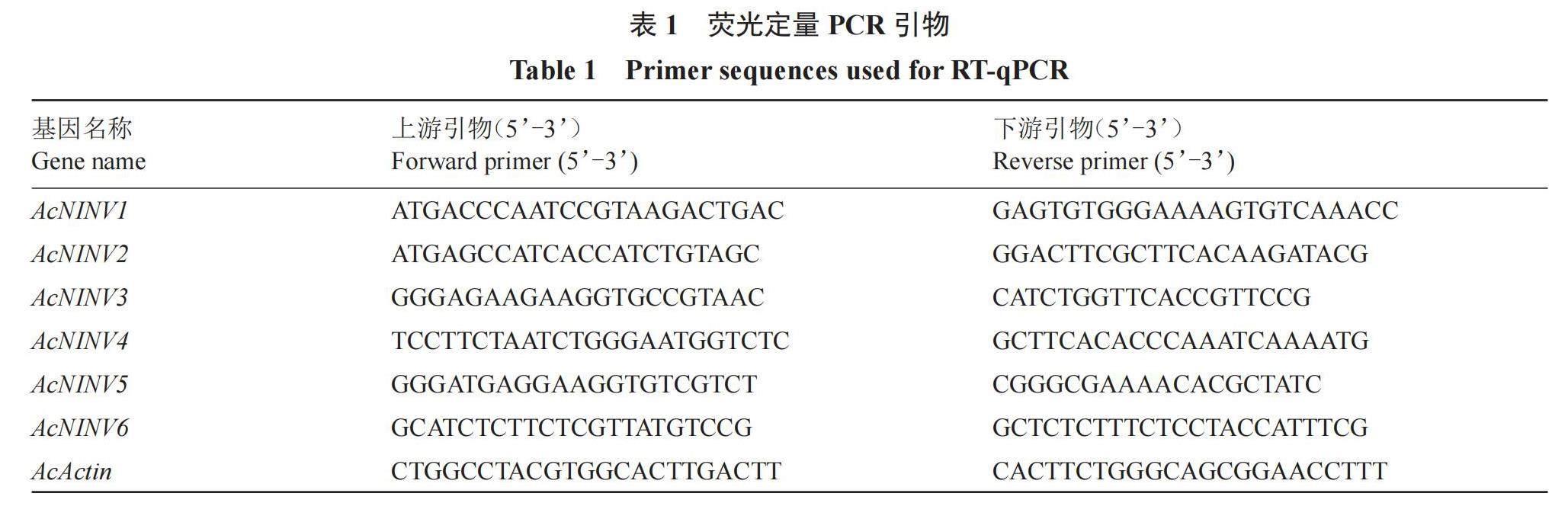
在LightCycler480 Ⅱ (Roche, Switzerland)上使用DyNAmo Flash SYBR Green qPCR kit (Thermo,USA)试剂盒进行实时荧光定量PCR(RT-qPCR)分析。用Primer Premier 5.0软件设计荧光定量引物(表1)。RT-qPCR反应总体积为10 μL:1 μL稀释cDNA、5 μL SYBR Green PCR Master Mix、引物最终浓度为250 nmol·L-1,以AcActin(HQ148720)作为内参基因[24]。反应条件如下:50 ℃ 2 min,95 ℃ 5 min,然后在95 ℃ 10 s,55 ℃ 30 s,72 ℃ 30 s下进行40个循环。每个反应都设3个技术重复,候选基因的相对表达水平用2?ΔΔCT方法进行分析[25]。
2 结果与分析
2.1 菠萝AcNINV基因家族的鉴定及特性分析
从菠萝基因组中分离出6个AcNINV基因家族成员,分别命名为AcNINV1~6。AcNINV蛋白的长度为556~673 aa,分子质量(MW)在63.03~74.80 ku之间变化,理论等电点为5.68~6.35,不稳定指数大于40,亲水性数值小于0。对这些AcNINV基因进行亚细胞定位预测,发现AcNINV3和AcNINV6分布于叶绿体上,其余家族成员都分布于质膜上(表2)。
2.2 菠萝AcNINV蛋白质二级结构分析
利用SOPMA在线分析工具预测菠萝AcNINV蛋白序列的二级结构(表3),菠萝AcNINV基因家族均含有α-螺旋、β-转角、无规则卷曲及延伸链等4种构型,但各部分所占比例明显不同。菠萝AcNINV基因家族的二级结构主要由α-螺旋和无规则卷曲组成,其次为延伸链,β-转角所占比例最低。
2.3 菠萝AcNINV基因家族染色体定位分析
利用TBtools软件绘制基因在染色体上的位置(图1),6个AcNINV基因分别定位于5条不同的染色体(LG)上。其中LG21上有2个基因(AcNINV5和AcNINV6),其余染色体上都只有1个AcNINV基因。
2.4 菠萝AcNINV基因共线性分析
为了探明AcNINV基因的进化关系,构建了菠萝与拟南芥、水稻的共线性图谱,从图2中可以看出菠萝AcNINV基因与拟南芥中仅有一对基因(AT3G05820.1与Aco006374.1)有共线性关系,但与水稻有8对基因(LOC_Os02g32730.1與Aco007782.1、LOC_Os04g3-
3490.1与Aco007782.1、LOC_Os02g34560.1与Aco007893.1、LOC_Os04g35280.1与Aco007893.1、LOC_Os04g35280.1与Aco003240.1、LOC_Os02g3-
4560.1与Aco003240.1、LOC_Os11g07440.1与Aco003240.1、LOC_Os04g35280.1与Aco011221.1)有共线性关系。
2.5 菠萝AcNINV基因结构和保守基序分析
对已鉴定的AcNINV基因的外显子/内含子结构进行分析,发现AcNINV外显子数量介于4到6个之间,其中AcNINV3和AcNINV6基因外显子数量都是6个,其余基因外显子都为4个;内含子数量介于3到5个之间,其中AcNINV3和AcNINV6基因内含子数量最高为5个,其余基因内含子数量为3个(图3)。利用MEME进行保守基序分析,在6个AcNINV基因中共预测了10个保守基序。其中AcNINV1、2、4、5均含有10个保守基序,但AcNINV3、6只有9个保守基序,缺少了motif 10。
2.6 菠萝AcNINV基因系统进化树分析
为了更好地阐明菠萝AcNINV基因家族之间的进化关系,将菠萝6个AcNINV基因与拟南芥、水稻、木薯等4个物种的NINV基因家族氨基酸序列构建了系统发育树。菠萝6个AcNINV基因被聚类到α和β两个亚族,其中AcNINV1、2、4、5属于β亚族,AcNINV3、6属于α亚族(图4)。
2.7 菠萝AcNINV基因启动子顺式作用元件分析
为了探析菠萝NINV基因是如何被调控表达的,利用Plant-CARE预测了目的基因上游2000 bp顺式作用元件。结果(图5)表明,光反应元件(MRE、ATC-motif、Box 4、L-box、GATA-motif、G-box、GT1-motif、TCCC-motif、3-AF1 binding site、LAMP-element、Sp1、TCT-motif、GA-motif、chs-CMA1a、AT1-motif、AE-box)的数量最多,其次是激素反应元件(ABRE、TGA-element、AuxRR-core、P-box、O2-site、TATC-box、TCA-element、TCA-element、 CGTCA-motif、TGACG-motif)和逆境反应元件(TC-rich repeats、LTR、ARE、GC-motif)。其中AcNINV5所含顺式作用元件数量最多,AcNINV2所含顺式作用元件数量最少。
2.8 菠萝果实发育过程中蔗糖含量的变化
在菠萝果实发育过程中,蔗糖含量呈先上升后下降的趋势(图6)。从20~40 DAA,蔗糖含量保持不变;此后,从50~80 DAA蔗糖含量急剧增加,并在80 DAA达到最高值,为20 DAA的126倍;从80~100 DAA急剧下降,但最终蔗糖含量仍比20 DAA高12倍[1]。正是由于蔗糖含量随着果实的生长发育而增加,因此果实的甜度会随着果实的生长发育而提高。
2.9 菠萝果实发育过程中AcNINV基因家族的表达分析
利用RT-qPCR技术研究了6个AcNINV基因在菠萝果实发育过程中的表达模式(图7),不同AcNINV基因成员其表达差异巨大。6个AcNINV基因在菠萝果实发育过程中的表达呈现出3种类型,第一种为上调表达,AcNINV2、3、6等3个基因属于这种类型;第二种为先下调再上调表达,但变化幅度不大,AcNINV1、5等2个基因属于这种类型;第三种为显著下调表达,AcNINV4基因属于这种类型。AcNINV4基因在20~40 DAA的表达量几乎没变化,但从40 DAA开始迅速下降,并持续到100 DAA,该基因在20 DAA的表达量是100 DAA的5倍。
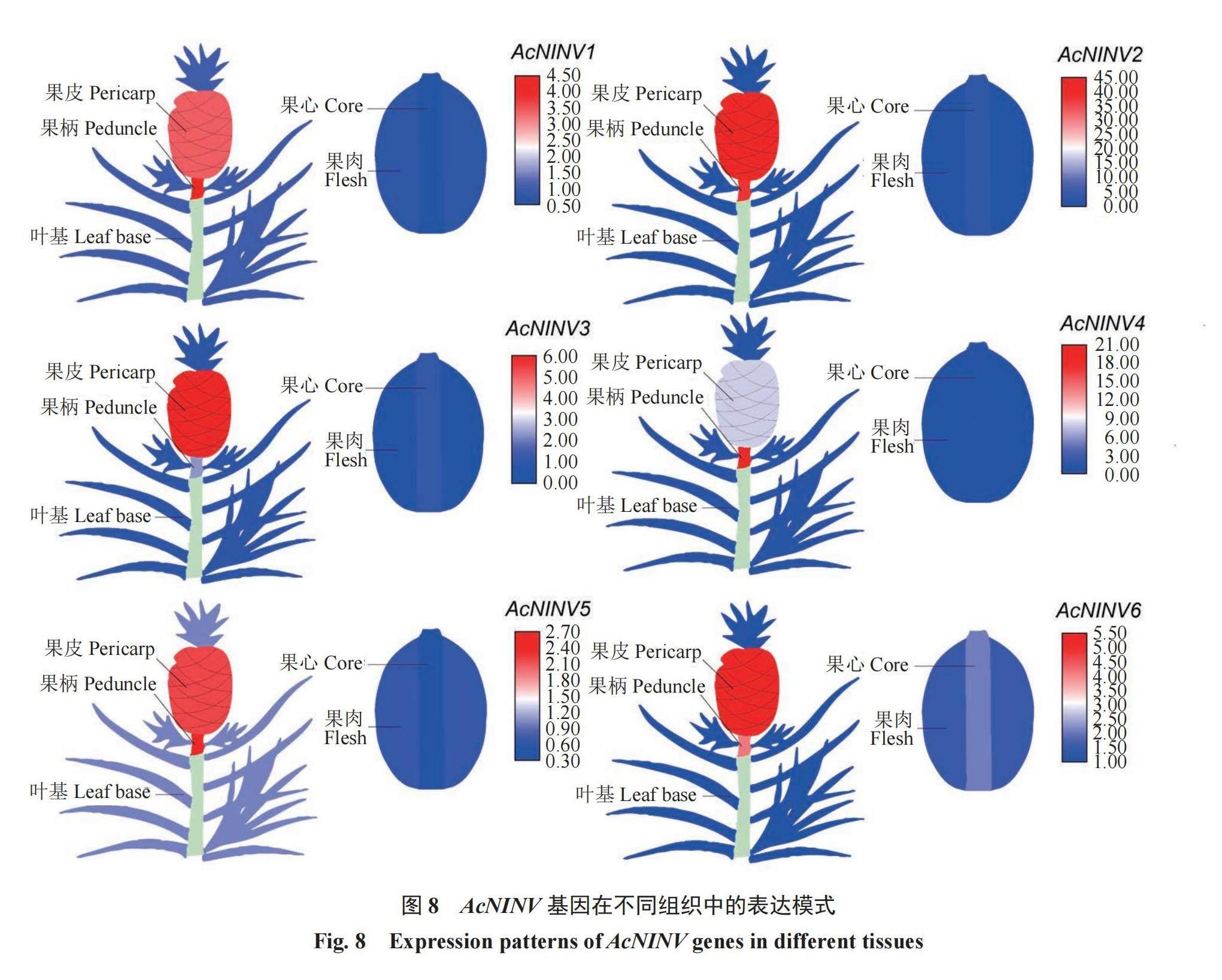
2.10 AcNINV基因在不同组织中的表达分析
为了阐明AcNINV基因不同家族成员的表达特性,利用RT-qPCR分析了不同组织的表达量,从图8可以看出,不同家族成员的表达特性不同,AcNINV1在果柄中的表达量最高,在果心中最低;AcNINV2、3、6在果皮中的表达量最高,在果肉中最低;AcNINV4、5在果柄中的表达量最高,在果肉中最低。综合AcNINV基因家族在不同组织中的表达特性发现,在果柄、果皮、果心中表达量最高的是AcNINV2基因,在果肉中表达量最高的是AcNINV6基因。
3 讨 论
蔗糖、果糖和葡萄糖是菠萝果实中的主要糖类[26],在本研究中蔗糖含量随果实发育而增加,这一结果和笔者课题组之前的结论一致[27]。笔者在本研究的菠萝中共鉴定出6个AcNINV基因成员,AcNINV蛋白的长度为556~673 aa,分子质量(MW)在63.03~74.80 ku之间变化,理论等电点为5.68~6.35,6个AcNINV基因位于5条染色体上。基因共线性分析发现菠萝与水稻NINV基因的共线性基因对有8对,但菠萝与拟南芥的共线性基因对只有1对。
NINV基因家族根据进化关系可以分为α和β亚族,其中α亚族定位在细胞核、叶绿体和线粒体上,β亚族定位在细胞溶质上[28-30]。另外,两个亚族成员的主要区别在于外显子/内含子数量不同,α亚族有6个外显子,β亚族只有4个外显子[17,19,31]。在辣椒中CaNINV基因外显子数量介于3到6个之间,CaNINV1、2、3基因均有6个外显子,属于α亚族,CaNINV4、5、6基因均有4个外显子,属于β亚族[12]。木薯11个MeNINV基因外显子数量介于4到6个之间,其中MeNINV1、6、7、8、9、10属于α亚族,MeNINV2、3、4、5和nINV1属于β亚族[17]。研究表明,AcNINV外显子数量介于4到6个之间,内含子数量介于3到5个之间,其中AcNINV3、6外显子的数量为6个,亚细胞定位预测发现这2个基因都分布于叶绿体上,属于α亚族;AcNINV1、2、4、5外显子的数量为4个,亚细胞定位预测发现这4个基因都分布于质膜上,属于β亚族。这些结果与前人的结论一致。
启动子分析表明,AcNINV家族的启动子区域含有激素反应元件、逆境反应元件和光反应元件。在毛白杨[32]和西瓜、甜瓜[33]中也发现类似结果。且本研究发现,在众多反应元件中光反应元件数量最多,据此推测光照条件能显著调控NINV基因的表达,进而影响NINV酶活性,最终影响蔗糖的含量,这也能解释为什么同一品种的菠萝在不同季节收获其果实糖积累的类型不同[34]。
NINV基因具有组织表达特异性[7]。Shen等[12]分析了辣椒7个基因家族成员在根、茎、叶、花芽、花、果实发育9个阶段的表达特性,发现CaNINV1和CaNINV6在所有組织中的表达量都较低,且未检测到CaNINV7表达。CaNINV4主要在根中表达,在果实仅有微弱表达。CaNINV2和CaNINV3在果实中的表达量高于其他组织。CaNINV5在所有组织中的表达量都是最高的。在铁皮石斛5个NI基因中,在根中表达量高的为DoNI1和DoNI3,在花中表达量高的为DoNI4和DoNI5,在茎中表达量高的为DoNI2[35]。在木薯中,MeNINV5在雄花中特异性表达,在叶片、雌花和果实中仅微弱表达,且在其他器官中未检测到;MeNINV1、4、7的表达模式相似,在茎、雄花和雌花中均有表达;MeNINV6、10主要在叶、茎、雄花和雌花中表达;MeNINV2、3和5在雄花中的表达量高于其他组织;MeNINV8在花中显著表达,在块茎根中仅微弱表达;MeNINV9在雌花中显著表达[17]。笔者研究发现,在6个AcNINV基因中,在果柄、果皮、果心中表达量最高的是AcNINV2基因,在果肉中表达量最高的是AcNINV6基因。推测这两个基因可能在植物器官生长中起到重要作用,因为前人研究发现在拟南芥中,At-A/N-Inv G基因影响侧根的生长[29];同时敲除At-A/N-Inv G和At-A/N-Inv I基因植株生长缓慢,矮化,根的伸长区生长缓慢[36];At-A/N-Inv C基因影响茎的伸长、花芽的分化[37-38];A/N-Inv H敲除突变体开花时间推迟、植株茎的伸长减弱[39]。百脉根NINV基因Ljinv1-1、Ljinv1-2、Ljinv1-3缺失导致植株生长缓慢,根部受损,影响花芽和小孢子分化[30,40]。敲除水稻OsCYT-INV1基因,水稻根系变短,开花延缓[41]。
同时前人研究表明NINV在蔗糖代谢中发挥重要作用[42],橡胶树乳管中HbNIN2参与蔗糖代谢,从而影响乳胶产量[42],橡胶HbNINa基因通过调控蔗糖代谢,进而在叶片发育过程中起到重要作用[43]。番茄果实SlCIN3基因在转色期和红熟期的表达量是绿熟期的4~5倍,据此推测该基因可能是番茄果实后期己糖代谢的关键基因[44]。杨宁等[32]将PtoNIN1过表达到拟南芥中,发现提高了转基因植株角果、茎、莲座叶的鲜质量,且促进了转基因植株蔗糖的代谢。铁皮石斛中DcNI4基因不可逆地将蔗糖分解为果糖和葡萄糖[16],NI的表达与葡萄果皮、果肉中葡萄糖和果糖的含量变化呈正相关[45]。在柑橘中研究发现,CitNI5可能参与了蔗糖代谢的调控,且该基因与果实蔗糖含量变化呈负相关[14]。Joubert等[46]利用RNAi技术,将NI基因转入甘蔗,发现NI活性显著下降,而蔗糖含量显著上升。笔者研究发现6个AcNINV基因在菠萝果实发育过程中呈现出不同的表达特性,AcNINV2、3、6等3个基因为上调表达;AcNINV1、5等2个基因为先下调再上调表达,但变化幅度不大;AcNINV4基因表现为显著下调,且从20~40 DAA,蔗糖含量基本保持不变,AcNINV4基因的表达量也基本保持不变;从40~80 DAA,蔗糖含量持续增加,AcNINV4基因的表达量呈先显著下降之后再有所回升的变化趋势;从80~100 DAA,蔗糖含量显著下降,AcNINV4基因的表达量稍有下降。综上所述,推测AcNINV4可能是催化果实蔗糖降解的水解酶基因。
4 结 论
笔者在菠萝中共鉴定出6个NINV基因,分布于5条不同染色体上,其二级结构主要由α-螺旋和无规则卷曲组成,该基因家族启动子区域光反应元件数量最多。AcNINV3、6外显子的数量为6个,分布于叶绿体上,属于α亚族;AcNINV1、2、4、5外显子的数量为4个,分布于质膜上,属于β亚族。在果柄、果皮、果心中表达量最高的是AcNINV2基因,在果肉中表达量最高的是AcNINV6基因。蔗糖含量随着菠萝果实成熟呈先升高后降低的变化趋势,而AcNINV4基因在菠萝果实成熟過程中呈下调表达,据此推测,AcNINV4可能是催化果实蔗糖降解的水解酶基因。
参考文献References:
[1] WU J Y,CHEN M,YAO Y L,FU Q,ZHU Z Y,ZHANG X M. Identification,characterisation,and expression profile analysis of the sucrose phosphate synthase gene family in pineapple (Ananas comosus)[J]. The Journal of Horticultural Science and Biotechnology,2022,97(2):201-210.
[2] 袁俊杰,陈文,李志勇,魏霜,马新华,卢乃会,杨卓瑜,渭婷玉,龙阳. 湛江关区鲜菠萝产业发展及出口对策探讨[J]. 质量安全与检验检测,2020,30(5):107-110.
YUAN Junjie,CHEN Wen,LI Zhiyong,WEI Shuang,MA Xinhua,LU Naihui,YANG Zhuoyu,WEI Tingyu,LONG Yang. Discussion on the development of pineapple industry and its export strategy in Zhanjiang[J]. Quality Safety Inspection and Testing,2020,30(5):107-110.
[3] PERSIA D,CAI G,DEL CASINO C,FALERI C,WILLEMSE M T M,CRESTI M. Sucrose synthase is associated with the cell wall of tobacco pollen tubes[J]. Plant Physiology,2008,147(4):1603-1618.
[4] BASSON C E,GROENEWALD J H,KOSSMANN J,CRONJ? C,BAUER R. Sugar and acid-related quality attributes and enzyme activities in strawberry fruits:Invertase is the main sucrose hydrolysing enzyme[J]. Food Chemistry,2010,121(4):1156-1162.
[5] 雷钰欣,曹子千,李纪璇,刘福庆,张双,许志茹. 高等植物转化酶的研究进展[J].分子植物育种,2021:1-20. (2021-07-08)[2023-10-20]. DOI:10.13271/j.mpb.022.001242.
LEI Yuxin,CAO Ziqian,LI Jixuan,LIU Fuqing,ZHANG Shuang,XU Zhiru. Research progress of invertase in higher plants[J]. Molecular Plant Breeding,2021:1-20. (2021-07-08)[2023-10-20]. DOI:10.13271/j.mpb.022.001242.
[6] XIANG L,LE ROY K,BOLOURI-MOGHADDAM M R,VANHAECKE M,LAMMENS W,ROLLAND F,VAN DEN ENDE W. Exploring the neutral invertase-oxidative stress defence connection in Arabidopsis thaliana[J]. Journal of Experimental Botany,2011,62(11):3849-3862.
[7] DAHRO B,WANG F,PENG T,LIU J H. PtrA/NINV,an alkaline/neutral invertase gene of Poncirus trifoliata,confers enhanced tolerance to multiple abiotic stresses by modulating ROS levels and maintaining photosynthetic efficiency[J]. BMC Plant Biology,2016,16:76.
[8] 陈美霞,赵从凯,陈学森,房师梅,张宪省. 杏果实发育过程中糖积累与蔗糖代谢相关酶的关系[J]. 果树学报,2009,26(3):320-324.
CHEN Meixia,ZHAO Congkai,CHEN Xuesen,FANG Shimei,ZHANG Xiansheng. Relationship between accumulation of sugar and sucrose-metabolizing enzymes during apricot fruit development[J]. Journal of Fruit Science,2009,26(3):320-324.
[9] 张玲. 草莓蔗糖代谢与转运相关基因对果实糖分积累的影响机理[D]. 兰州:甘肃农业大学,2017.
ZHANG Ling. The effect of genes for sucrose metabolism and transportion on fruit sugar accumulation in strawberry (Fragaria × ananassa Duch.)[D]. Lanzhou:Gansu Agricultural University,2017.
[10] QIAN W J,YUE C,WANG Y C,CAO H L,LI N N,WANG L,HAO X Y,WANG X C,XIAO B,YANG Y J. Identification of the invertase gene family (INVs) in tea plant and their expression analysis under abiotic stress[J]. Plant Cell Reports,2016,35(11):2269-2283.
[11] CHEN J Q,BLACK C C. Biochemical and immunological properties of alkaline invertase isolated from sprouting soybean hypocotyls[J]. Archives of Biochemistry and Biophysics,1992,295(1):61-69.
[12] SHEN L B,YAO Y,HE H,QIN Y L,LIU Z J,LIU W X,QI Z Q,YANG L J,CAO Z M,YANG Y. Genome-wide identification,expression,and functional analysis of the alkaline/neutral invertase gene family in pepper[J]. International Journal of Molecular Sciences,2018,19(1):224.
[13] JI X M,VAN DEN ENDE W,VAN LAERE A,CHENG S H,BENNETT J. Structure,evolution,and expression of the two invertase gene families of rice[J]. Journal of Molecular Evolution,2005,60(5):615-634.
[14] 李恒. 柑橘CitNI5和CitMYB52參与蔗糖代谢的转录调控初探[D]. 杭州:浙江大学,2020.
LI Heng. Preliminary research on transcriptional regulation of citrus CitNI5 and CitMYB52 in sucrose metabolism[D]. Hangzhou:Zhejiang University,2020.
[15] MARUTA T,OTORI K,TABUCHI T,TANABE N,TAMOI M,SHIGEOKA S. New insights into the regulation of greening and carbon-nitrogen balance by sugar metabolism through a plastidic invertase[J]. Plant Signaling & Behavior,2010,5(9):1131-1133.
[16] 刘晨. 铁皮石斛多糖代谢途径关键催化酶中性/碱性转化酶DcNI4的挖掘与功能研究[D]. 杭州:浙江农林大学,2021.
LIU Chen. Mining and functional characterization of key catalase enzyme alkaline/neutral invertase DcNI4 involved in polysaccharide biosynthesis pathway in Dendrobium catenatum[D]. Hangzhou:Zhejiang A & F University,2021.
[17] YAO Y,GENG M T,WU X H,LIU J,LI R M,HU X W,GUO J C. Genome-wide identification,expression,and activity analysis of alkaline/neutral invertase gene family from cassava (Manihot esculenta Crantz)[J]. Plant Molecular Biology Reporter,2015,33(2):304-315.
[18] HYUN T K,EOM S H,KIM J S. Genomic analysis and gene structure of the two invertase families in the domesticated apple (Malus × domestica Borkh.)[J]. Plant Omics,2011,4(7):391-399.
[19] BOCOCK P N,MORSE A M,DERVINIS C,DAVIS J M. Evolution and diversity of invertase genes in Populus trichocarpa[J]. Planta,2008,227(3):565-576.
[20] MING R,VANBUREN R,WAI C M,TANG H B,SCHATZ M C,BOWERS J E,LYONS E,WANG M L,CHEN J,BIGGERS E,ZHANG J S,HUANG L X,ZHANG L M,MIAO W J,ZHANG J,YE Z Y,MIAO C Y,LIN Z C,WANG H,ZHOU H Y,YIM W C,PRIEST H D,ZHENG C F,WOODHOUSE M,EDGER P P,GUYOT R,GUO H B,GUO H,ZHENG G Y,SINGH R,SHARMA A,MIN X J,ZHENG Y,LEE H Y,GURTOWSKI J,SEDLAZECK F J,HARKESS A,MCKAIN M R,LIAO Z Y,FANG J P,LIU J,ZHANG X D,ZHANG Q,HU W C,QIN Y,WANG K,CHEN L Y,SHIRLEY N,LIN Y R,LIU L Y,HERNANDEZ A G,WRIGHT C L,BULONE V,TUSKAN G A,HEATH K,ZEE F,MOORE P H,SUNKAR R,LEEBENS-MACK J H,MOCKLER T,BENNETZEN J L,FREELING M,SANKOFF D,PATERSON A H,ZHU X G,YANG X H,SMITH J A C,CUSHMAN J C,PAULL R E,YU Q Y. The pineapple genome and the evolution of CAM photosynthesis[J]. Nature Genetics,2015,47(12):1435-1442.
[21] CHEN C J,CHEN H,ZHANG Y,THOMAS H R,FRANK M H,HE Y H,XIA R. TBtools:An integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant,2020,13(8):1194-1202.
[22] ZHANG M F,LI Z L. A comparison of sugar-accumulating patterns and relative compositions in developing fruits of two oriental melon varieties as determined by HPLC[J]. Food Chemistry,2005,90(4):785-790.
[23] WU J Y,ZHANG H N,LIU L Q,LI W C,WEI Y Z,SHI S Y. Validation of reference genes for RT-qPCR studies of gene expression in preharvest and postharvest longan fruits under different experimental conditions[J]. Frontiers in Plant Science,2016,7:780.
[24] LI Y H,WU Q S,HUANG X,LIU S H,ZHANG H N,ZHANG Z,SUN G M. Molecular cloning and characterization of four genes encoding ethylene receptors associated with pineapple (Ananas comosus L.) flowering[J]. Frontiers in Plant Science,2016,7:710.
[25] LIVAK K J,SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2?ΔΔCT method[J]. Methods,2001,25(4):402-408.
[26] 张秀梅,杜丽清,谢江辉,陈佳瑛,弓德强,李伟才. 蔗糖代谢相关酶在卡因菠萝果实糖积累中的作用[J]. 果树学报,2006,23(5):707-710.
ZHANG Xiumei,DU Liqing,XIE Jianghui,CHEN Jiaying,GONG Deqiang,LI Weicai. Roles of sucrose-metabolizing enzymes in accumulation of sugars in Cayenne pineapple fruit[J]. Journal of Fruit Science,2006,23(5):707-710.
[27] ZHANG X M,LIU S H,DU L Q,YAO Y L,WU J Y. Activities,transcript levels,and subcellular localizations of sucrose phosphate synthase,sucrose synthase,and neutral invertase and change in sucrose content during fruit development in pineapple (Ananas comosus)[J]. The Journal of Horticultural Science and Biotechnology,2019,94(5):573-579.
[28] 宋莉璐,張荃. 植物中参与活性氧调控的基因网络[J]. 生命科学,2007,19(3):346-352.
SONG Lilu,ZHANG Quan. Reactive oxygen gene network of plants and its regulation[J]. Chinese Bulletin of Life Sciences,2007,19(3):346-352.
[29] LOU Y,GOU J Y,XUE H W. PIP5K9,an Arabidopsis phosphatidylinositol monophosphate kinase,interacts with a cytosolic invertase to negatively regulate sugar-mediated root growth[J]. The Plant Cell,2007,19(1):163-181.
[30] WINGLER A,ROITSCH T. Metabolic regulation of leaf senescence:Interactions of sugar signalling with biotic and abiotic stress responses[J]. Plant Biology,2008,10(Suppl. 1):50-62.
[31] FUJII S,HAYASHI T,MIZUNO K. Sucrose synthase is an integral component of the cellulose synthesis machinery[J]. Plant and Cell Physiology,2010,51(2):294-301.
[32] 杨宁,杨雄,李国雷,陈仲. 毛白杨碱性/中性转化酶基因PtoNIN1的克隆与功能研究[J]. 北京林业大学学报,2023,45(5):35-46.
YANG Ning,YANG Xiong,LI Guolei,CHEN Zhong. Cloning and functional study of the alkaline/neutral invertase gene PtoNIN1 in Populus tomentosa[J]. Journal of Beijing Forestry University,2023,45(5):35-46.
[33] 熊思亦,张聪聪,马荣雪,闫星,魏春华,张显. 西瓜、甜瓜蔗糖转化酶基因家族鉴定及表达分析[J].分子植物育种,2023,21(12):3829-3839.
XIONG Siyi,ZHANG Congcong,MA Rongxue,YAN Xing,WEI Chunhua,ZHANG Xian. Identification and expression analysis of invertase gene family in watermelon and melon[J]. Molecular Plant Breeding,2023,21(12):3829-3839.
[34] 张秀梅,李建国,窦美安,姚艳丽,杜丽清,孙光明. 不同季节菠萝果实糖积累的差异[J]. 园艺学报,2010,37(11):1751-1758.
ZHANG Xiumei,LI Jianguo,DOU Meian,YAO Yanli,DU Liqing,SUN Guangming. Difference in sugar accumulation of pineapple fruit harvested in different seasons[J]. Acta Horticulturae Sinica,2010,37(11):1751-1758.
[35] 苗小荣,牛俊奇,王爱勤,黄维,何龙飞. 铁皮石斛中性/碱性转化酶家族基因的克隆和表达分析[J].分子植物育种,2022,20(19):6362-6370.
MIAO Xiaorong,NIU Junqi,WANG Aiqin,HUANG Wei,HE Longfei. Cloning and expression analysis of neutral/alkaline invertase gene in Dendrobium officinale[J]. Molecular Plant Breeding,2022,20(19):6362-6370.
[36] BARRATT D H P,DERBYSHIRE P,FINDLAY K,PIKE M,WELLNER N,LUNN J,FEIL R,SIMPSON C,MAULE A J,SMITH A M. Normal growth of Arabidopsis requires cytosolic invertase but not sucrose synthase[J]. Proceedings of the National Academy of Sciences of the United States of America,2009,106(31):13124-13129.
[37] TAMOI M,TABUCHI T,DEMURATANI M,OTORI K,TANABE N,MARUTA T,SHIGEOKA S. Point mutation of a plastidic invertase inhibits development of the photosynthetic apparatus and enhances nitrate assimilation in sugar-treated Arabidopsis seedlings[J]. The Journal of Biological Chemistry,2010,285(20):15399-15407.
[38] BAILEY T L,BODEN M,BUSKE F A,FRITH M,GRANT C E,CLEMENTI L,REN J Y,LI W W,NOBLE W S. MEME SUITE:Tools for motif discovery and searching[J]. Nucleic Acids Research,2009,37(Web Server issue):W202-W208.
[39] BATTAGLIA M E,MARTIN M V,LECHNER L,MART?NEZ-NO?L G M A,SALERNO G L. The riddle of mitochondrial alkaline/neutral invertases:A novel Arabidopsis isoform mainly present in reproductive tissues and involved in root ROS production[J]. PLoS One,2017,12(9):e0185286.
[40] WELHAM T,PIKE J,HORST I,FLEMETAKIS E,KATINAKIS P,KANEKO T,SATO S,TABATA S,PERRY J,PARNISKE M,WANG T L. A cytosolic invertase is required for normal growth and cell development in the model legume,Lotus japonicus[J]. Journal of Experimental Botany,2009,60(12):3353-3365.
[41] JIA L Q,ZHANG B T,MAO C Z,LI J H,WU Y R,WU P,WU Z C. OsCYT-INV1 for alkaline/neutral invertase is involved in root cell development and reproductivity in rice (Oryza sativa L.)[J]. Planta,2008,228(1):51-59.
[42] LIU S J,LAN J X,ZHOU B H,QIN Y X,ZHOU Y H,XIAO X H,YANG J H,GOU J Q,QI J Y,HUANG Y C,TANG C R. HbNIN2,a cytosolic alkaline/neutral-invertase,is responsible for sucrose catabolism in rubber-producing laticifers of Hevea brasiliensis (para rubber tree)[J]. New Phytologist,2015,206(2):709-725.
[43] 肖小虎,戚繼艳,方永军,曹冰,龙翔宇,唐朝荣. 巴西橡胶树中碱性转化酶HbNINa基因的克隆和表达模式分析[J]. 热带作物学报,2013,34(7):1264-1269.
XIAO Xiaohu,QI Jiyan,FANG Yongjun,CAO Bing,LONG Xiangyu,TANG Chaorong. Cloning and expression analysis of an alkaline/neutral invertase gene in Hevea brasliensis[J]. Chinese Journal of Tropical Crops,2013,34(7):1264-1269.
[44] 张琼琼. 番茄碱性/中性转化酶SlCIN3基因功能研究[D]. 沈阳:沈阳农业大学,2019.
ZHANG Qiongqiong. The primary study of an alkaline/neutral invertase SlCIN3 gene function in tomato[D]. Shenyang:Shenyang Agricultural University,2019.
[45] NONIS A,RUPERTI B,PIERASCO A,CANAGUIER A,ADAM-BLONDON A F,DI GASPERO G,VIZZOTTO G. Neutral invertases in grapevine and comparative analysis with Arabidopsis,poplar and rice[J]. Planta,2008,229(1):129-142.
[46] JOUBERT D,BOSCH S,BOTHA F C,KOSSMANN J,GROENEWALD J H. Down-regulation of neutral invertase activity in sugarcane cell suspension cultures leads to increased sucrose accumulation[J]. South African Journal of Botany,2007,73(2):293.
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
江苏农业科学(2022年17期)2022-09-24
四川大学学报(自然科学版)(2021年4期)2021-07-15
中国生殖健康(2020年4期)2021-01-18
金桥(2020年10期)2020-11-26
中国农业科学(2020年5期)2020-03-30
阅读与作文(小学高年级版)(2019年5期)2019-05-27
好孩子画报(2019年12期)2019-01-13
中国生殖健康(2018年4期)2018-11-06
上海师范大学学报·自然科学版(2016年5期)2016-12-14
