
锦红冰糖橙大果芽变果实结构解剖、激素变化和转录组分析
2024-04-30 00:38许园园谭世水张玲段少伟郭玲霞周铁李菲菲韩健李先信王聪田陈鹏
果树学报 2024年4期
许园园 谭世水 张玲 段少伟 郭玲霞 周铁 李菲菲 韩健 李先信 王聪田 陈鹏


摘 要:【目的】探究錦红冰糖橙大果芽变的形成原因。【方法】以锦红和大果芽变为试材,采用石蜡切片、HPLC-MS/MS和转录组测序技术对其在细胞形态、激素含量和基因表达方面进行比较分析。【结果】与锦红相比,大果芽变果肉内汁胞数量增多,且汁胞变大。石蜡切片结果表明,大果芽变汁胞内细胞数量多于锦红。大果芽变内生长素和赤霉素含量在花后70和120 d均高于锦红,玉米素含量在花后70、120和170 d均低于锦红。对盛花后70、120和170 d的汁胞进行转录组分析得到4597个差异基因,其中24个激素信号相关基因在两个及以上时期差异表达。在大果芽变中有4个生长素相关基因在花后70 d上调表达。细胞分裂素相关基因ARR2在大果芽变花后70和120 d均下调表达。细胞周期蛋白CycD3在大果芽变花后170 d上调表达。【结论】锦红冰糖橙大果芽变的变异可能与汁胞发育有关,并且受激素水平影响。转录组分析为解析锦红冰糖橙大果芽变的分子机制奠定了基础。
关键词:柑橘;芽变;果实大小;石蜡切片;激素
中图分类号:S666 文献标志码:A 文章编号:1009-9980(2024)04-0611-14
Anatomical structure, hormone change, and transcriptome analysis of the large-fruit mutant of Jinhong Bingtang Orange
XU Yuanyuan1, 3, TAN Shishui2#, ZHANG Ling2, DUAN Shaowei2, GUO Lingxia1, 3, ZHOU Tie1, 3, LI Fei-
fei1, 3, HAN Jian1, 3, LI Xianxin1, WANG Congtian1, 3, CHEN Peng1, 3*
(1Hunan Horticultural Research Institute, Hunan Academy of Agricultural Sciences/Scientific Observation and Experiment Station of Fruit Trees in Central China, Ministry of Agriculture and Rural Affairs, Changsha 410125, Hunan, China; 2Citrus Industrialization Office of Mayang County, Mayang 419400, Hunan, China; 3Yuelu Mountain Laboratory, Changsha 410125, Hunan, China)
Abstract: 【Objective】 Fruit size is a major agronomic trait for evaluating fruit appearance quality. Therefore, it is important to analyze the molecular mechanism of fruit size for high-quality citrus breeding. Bud sport is one of the sources of fruit size mutation. A large-fruit mutant of Jinhong was found previously. In this research, we explored the mechanism of the mutation in order to provide target genes for citrus molecular design breeding. 【Methods】 Jinhong Bingtang orange and its large-fruit mutant as experimental material were sampled at 30, 50, 70, 90, 120, 150, 170, 190 and 250 days after bloom (DAB). Juice sacs at 70, 120 and 170 d were collected for RNA-seq. Young leaves of large-fruit mutant were collected for ploidy testing. Volume of juice sacs was determined using water replacement method. Paraffin section was used for cytological observation of juice sacs. The plant hormones were determined by liquid chromatography-mass spectrometry. Transcriptome analysis were performed on the above samples. Differentially expressed genes (DEGs) between the wild type and the mutant at same developmental stage were screened on the standard of FDR<0.05. Common DEGs of those three developmental stages were obtained using Venn analysis. GO and KEGG enrichments were mapped using TBtools, and GO and KEGG were used to analyze the DEGs between Jinhong and its large-fruit mutant. Genes related to plant hormones were shown with the heat maps, which were drawn based on FPKM of the DEGs. 【Results】 Flow cytometry analysis showed that the large-fruit mutant was a diploid. Jinhong and large-fruit mutant had the same number (11) of carpels. Compared with Jinhong, the number of juice sacs of the large-fruit mutant increased, and volume of juice sacs enlarged. Results of paraffin section showed that the cell number of the large-fruit mutant was larger than that of Jinhong. IAA content of the mutant was always higher than that of Jinhong at 70 and 120 DAB, but it was lower at 170 DAB. The GA3 content in the large-fruit mutant was always greater. ZT content was low, and the ZT content of the large-fruit mutant was always lower than that of Jinhong at the three developmental stages. Except at 70 DAB, JA content of the large-fruit mutant was lower than that of Jinhong. At 70 DAB, there were 3118 DEGs between the two materials, including 1549 up-regulated and 1569 down-regulated genes, which were enriched in plant hormone signal transduction, MAPK signaling pathway, plant-pathogen, etc. At 120 DAB, there were 1952 DEGs, with 377 up-regulated and 1575 down-regulated, which were enriched in photosynthesis - antenna proteins, plant-pathogen interaction and MAPK signaling pathway, etc. At 170 DAB, there were 611 DEGs, with 372 up-regulated and 239 down-regulated, enriched in photosynthesis - antenna proteins, phenylpropanoid biosynthesis, etc. Besides, 88 DEGs were identified in the three stages and KEGG analysis showed that the top five most significantly enriched pathway were fatty acid elongation, photosynthesis-antenna proteins, cutin, suberine and wax biosynthesis, phenylpropanoid biosynthesis and plant hormone signal transduction. Furthermore, DEGs encoding hormone signaling were further analyzed. 24 DEGs were found at least two periods, including 4 genes related to ABA, 6 related to IAA, 3 related to BR, 2 related to CTK, and other hormone-related genes. Among them, GH3.6 was up-regulated in the large-fruit mutant at 70 and 120 DAB. IAA27 was up-regulated in the large-fruit mutant at 120 and 170 DAB, and the difference gradually increased. GH3.1, SAUR36 and auxin-induced protein 22D were up-regulated in the large-fruit mutant at 70 DAB. ARR2, TIFY10A and TGA9 were down-regulated in large-fruit mutant at 70 and 120 DAB, and was up-regulated at 170 DAB. 【Conclusion】 the development of juice sacs was possibly the cause of large-fruit phenotype in the mutant, and hormone levels may influence fruit size. The transcriptome analysis provided a relatively complete molecular platform for future studies on the difference of citrus fruit size.
Key words: Citrus; Bud sport; Fruit size; Paraffin section; Hormone
果实大小是一种外观品质,也是判定果实品级的重要农艺性状,直接影响果实的经济价值[1]。果实大小受自身遗传背景、栽培条件和植物激素等因素的影响。生产上常采用环剥、疏花疏果等措施来增大果实[2-4],或者通过喷施生长调节剂来达到疏花疏果的目的,激素调节剂还能配合环剥技术来改变果实大小[4-6]。此外,植物激素调节剂还能单独用于果实发育早期来促进果实膨大,如生长素[7-11]、细胞分裂素[12-14]。
果实大小是受多个基因调控的数量性状。研究表明,果实大小涉及多种植物激素的调控,尤其是生长素和细胞分裂素。Su等[15]发现,沉默SlIAA17后得到的果实变大,与野生型相比,沉默系番茄果皮内细胞数量增多、果皮厚度增加。Peng等[16]的研究表明,EjSAUR22-TRV2植株果实变小,石蜡切片结果显示EjSAUR22-TRV2果肉细胞变小。目前,生长素调控果实大小的信号通路已有少量报道。Zhao等[17]研究认为CsFUL1A通过结合PIN1和PIN7启动子来抑制他们的转录水平,使黄瓜果实内生长素的积累量减少,黄瓜变短。Zhang等[18]的研究表明,CsMYB77结合在PIN5启动子上并上调其表达,通过降低游离IAA的含量来参与调控柑橘果实大小。细胞分裂素促进细胞分裂,在果实发育早期起重要作用。细胞色素氧化酶(cytokinin oxidase,CKX)是细胞分裂素代谢通路上的关键酶。Gan等[19]研究表明,与野生型相比,AtCKX2超表达植株的番茄果实变小。除生长素和细胞分裂素外,其他激素如赤霉素[20]、油菜素内酯[21]均参与果实大小发育过程。这些激素相关基因多通过影响细胞数量和细胞大小来改变果实大小。
柑橘属于芸香科柑橘属,在中国已有4000多年的栽培历史,是中国南方种植面积最大、产量最高的经济果树。柑橘销售价格一方面受自身品种影响,另一方面也受果实大小调控。栽培管理技术的优化是目前提升柑橘优果率的主要措施,但存在着生产成本高、不受控因素多等问题。转录组测序技术在植物性状控制基因挖掘方面具有重要作用[22-23]。笔者课题组前期发现锦红冰糖橙的芽变材料,该芽变成熟果实的大小约是锦红冰糖橙的1.5倍。笔者以锦红冰糖橙大果芽变为材料,通过石蜡切片技术、植物激素含量测定和转录组测序来解释大果芽变的形成机制,并筛选出冰糖橙果实大小相关基因,为柑橘果实大小分子设计育种提供理论基础。
1 材料和方法
1.1 试验材料
供试材料是锦红[C. sinensis (L.) Osbeck ‘Jinhong]及其大果芽变。每个材料各挑选3株树龄一致、长势健壮的样本树。在盛花后30、50、70、90、120、150、170、190和250 d取样。每次取样均从树冠中上部外围随机采摘10个果实。采集大果芽变的幼嫩叶片进行染色体倍性检测。并将盛花后70、120和170 d汁胞保存在-80 ℃冰箱中,以备植物激素测定和转录组测序。
1.2 果实质量、囊瓣数、汁胞数及其体积测定
用分析天平测定锦红和大果芽变成熟期的果实质量。用游标卡尺测量锦红和大果芽变果实发育过程的果肉横径,并统计果实内囊瓣数目和囊瓣内汁胞数量。采用浸水法估算单个汁胞体积,即向容器中加入水,此时体积记为V1,再将已知数量(S)的汁胞放入容器内,并记下体积V2,则单个汁胞体积的计算方式是V汁胞=(V2-V1)/S。
1.3 汁胞组织切片
对锦红和大果芽变成熟期的汁胞进行石蜡切片。用FAA固定液固定囊瓣中部的汁胞后,依次在70%、80%、90%和100%乙醇溶液中脱水,每次10 min,再用二甲苯清洗1 h后包埋在石蜡中。使用Lei-ca RM2265切片机获取薄切片。依次将切片放入环保型脱蜡液Ⅰ、Ⅱ中,各20 min。用无水乙醇Ⅰ、无水乙醇Ⅱ和75%乙醇依次清洗5 min后用蒸馏水清洗。将清洗好的切片依次放入番红、固绿染色液中,并用95%乙醇清洗,经脱水、透明、封片后进行图像采集。
1.4 植物激素测定
用液氮将样品研磨成粉末,称取1 g粉末放入10 mL乙腈溶液中,并加入8 μL内标母液,然后在4 ℃条件下提取过夜。4 ℃,12 000 r·min-1离心5 min,抽取上清液,随后在沉淀中再次加入5 mL乙腈溶液,并过夜提取,2次重复后合并3次所得上清液。在上清液中加入适量C18和GCB净化其中杂质。4 ℃,12 000 r·min-1离心5 min,抽取上清液,并用氮气吹干,以400 μL甲醇复溶,过0.22 μm有机相滤膜,上机检测。标准品为D-IAA、D-JA、D-GA4、D-Zeatin(Sigma)。使用终质量浓度为0.1、0.2、0.5、2、5、20、50、200 ng·mL-1绘制标准曲线。
1.5 转录组测序和差异基因鉴定
利用Trizol方法提取锦红冰糖橙和大果芽变汁胞的基因组总RNA。使用Nanodrop 2000检测RNA的浓度和纯度来评估RNA质量,合格后进行文库构建。用Oligo(dT)的磁珠富集有polyA尾巴的mRNA,用打断buffer把获得的RNA片段化后,再用随机的N6引物进行反转录,合成cDNA二链形成双链DNA,把合成的双链DNA末端补平并5端磷酸化,3端形成突出一个个“A”的黏末端,再连接一个3端有凸出“T”的鼓泡状的接头,连接产物通过特异的引物进行PCR扩增,PCR产物热变性成单链,再用一段桥式引物将单链DNA环化,得到单链环状DNA文库。文库检测合格后,使用MGI高通量测序仪进行测序。
对各样本raw reads进行过滤,得到clean reads,与甜橙参考基因组(http://citrus.hzau.edu.cn/download.php Citrus sinensis v3.0)比对。利用RSEM,调用bowtie2的比对结果进行统计,得到每个样品比对到每个转录本上的reads数目,并将其换算为FPKM(Fragments Per Kilobase per Million bases),每个基因或转录本的表达水平用FPKM值来表示。以|log2(FoldChange)|>1 & FDR<0.05為筛选阈值获得差异基因。使用Blast2GO程序和KOBAS软件进行GO和KEGG富集分析,使用维恩图获得不同发育时期共有的差异基因,使用GraphPad Prism 8软件绘制表达热图。
1.6 数据分析
采用Microsoft Excel 2019软件完成数据处理和绘图,采用SPSS软件进行差异显著性分析。
2 结果与分析
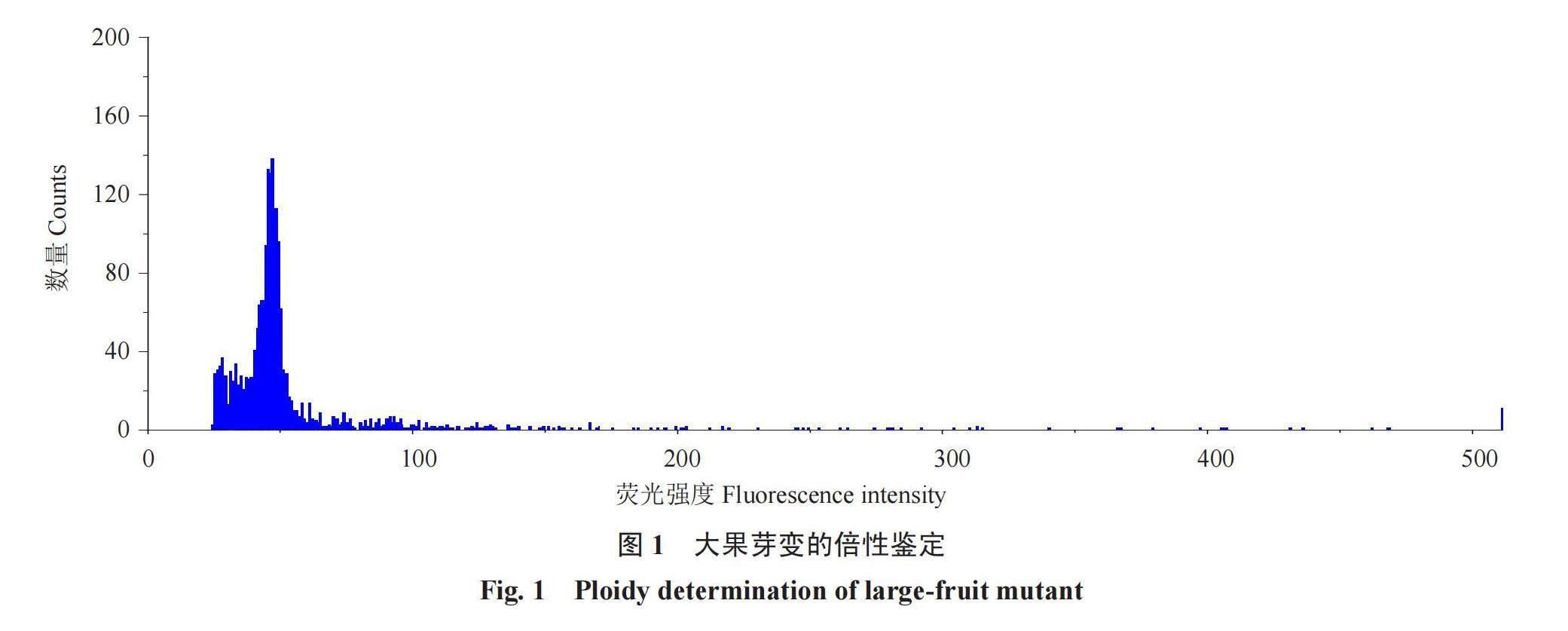
2.1 大果芽变的倍性鉴定
染色体数目加倍会引起果实变大。如图1所示,流式细胞仪结果表明,该大果芽变是二倍体,同锦红的倍性相同。说明大果型不是由染色体加倍造成的。
2.2 大果芽变的果实发育
如图2所示,锦红和大果芽变的果实逐渐增大。与锦红相比,自盛花后70 d(70 DAB),大果芽变的果实更大、果肉更厚。为探究大果芽变果肉增厚的原因,统计并测量锦红和大果芽变120 DAB、170 DAB和250 DAB时的囊瓣数量、汁胞数量和体积。结果表明,同一时期锦红和大果芽变之间的囊瓣和汁胞数目没有显著差异,但大果芽变果实内汁胞数目都略高于锦红果实内的汁胞数目,并且汁胞体积显著大于锦红的汁胞体积(表1)。石蜡切片的结果表明,大果芽变汁胞内细胞数量多于锦红(图3)。
2.3 锦红及其大果芽变汁胞内源激素含量分析
对锦红和大果芽变70 DAB、120 DAB和170 DAB的汁胞进行内源激素测定。结果如图4所示,在70 DAB和120 DAB,大果芽变中IAA含量显著高于锦红,而在临近转色期的170 DAB,两者IAA含量均下降,且大果芽变中IAA含量下降幅度高于锦红(图4-A)。在70 DAB、120 DAB 和170 DAB,锦红和大果芽变中GA3含量均逐步下降,且大果芽变中GA3含量始终高于锦红(图4-B);两者ZT含量(ρ)较低,不足0.5 ng·mL-1,在70 DAB、120 DAB和170 DAB,大果芽变中ZT含量都显著低于锦红(图4-C);在70 DAB,大果芽变中JA含量显著高于锦红,随后两者JA含量均下降,到170 DAB,大果芽变中JA含量显著低于锦红(图4-D)。综上所述,大果芽变和锦红之间汁胞发育差异是受多激素调控的,而且生长素和赤霉素可能是引起两者汁胞发育不同的关键激素。
2.4 锦红及其大果芽变盛花后170 d内汁胞转录组测序质量评估
为了进一步探究大果芽变形成的分子机制,对锦红和大果芽变盛花后3个时间点的汁胞进行转录组测序。锦红记为WT,大果芽变记为MT。从表2可以看出,18个样品的原始数据经过严格的质量过滤后共得到201.74×106个clean_reads,各样品的Q20含量都高于97%,Q30含量高于91.5%,GC含量高于44%,说明测序结果真实可信。
2.5 差异基因分析
对WT与MT(WT vs MT)同一阶段的差异表达基因(DEGs)(|log2(Fold Change)|>1,FDR<0.05)进行统计。DEGs数量随着汁胞发育而减少,其中70 DAB时的DEGs最多,有1549个上调基因,1569个下调基因;170 DAB时的DEGs最少,有372个上调基因,239个下调基因(图5-A)。维恩图结果表明,70 DAB、120 DAB和170 DAB三个时间点存在88个DEGs重叠(图5-B)。
2.6 差异基因注释
通常情况下,不同时期共有的差异基因可能为关键基因。对70 DAB、120 DAB和170 DAB的88个共有差异基因进行GO和KEGG分析。GO注释结果表明,差异基因参与的生物学过程为细胞和代谢过程,分子功能体现在结合和催化活性方面(图6-A)。参与的KEGG通路是脂肪酸延长、光合作用-天线蛋白、角质、木栓质和蜡质生物合成、苯丙烷代谢和植物激素信号传导(图6-B)。
分别对同一阶段下的DEGs进行GO和KEGG分析。GO富集结果表明,在70 DAB、120 DAB和170 DAB,DEGs参与的生物学过程都是细胞和代谢过程,参与的细胞组分都是细胞解剖实体,参與的分子功能多集中在结合和催化活性两方面(图7-A~C)。KEGG分析结果表明,在70 DAB,DEGs主要富集在植物激素信号传导、MAPK信号通路、植物-病原菌互作、色氨酸代谢以及苯丙烷代谢(图7-D);在120 DAB,DEGs主要富集在光合作用-天线蛋白、植物-病原菌互作、MAPK信号通路、植物激素信号传导、光合作用的碳固定和苯丙烷代谢(图7-E);在170 DAB,DEGs主要富集在光合作用-天线蛋白、苯丙烷代谢、角质、木栓质和蜡质生物合成、类黄酮生物合成和MAPK信号通路上(图7-F)。三个时期的差异基因都富集到植物激素信号传导、MAPK信号通路、苯丙烷代谢、光合作用-天线蛋白通路上,但富集到信号通路上的基因和显著性存在差异。
2.7 植物激素信号传导基因的表达分析
激素分析结果表明,植物激素影响汁胞发育(图4)。KEGG分析结果表明,在70 DAB、120 DAB和170 DAB,分别有61、38和12个DEGs富集到植物激素信号传导途径上。24个DEGs在两个及以上时期差异表达,包括19个已知功能的DEGs和5个未知功能的DEGs。笔者对19个DEGs进行热图分析,结果表明,它们分布在生长素(IAA)、脱落酸(ABA)、细胞分裂素(CTK)、油菜素内酯(BR)等7种激素途径上。
如图8所示,在脱落酸信号中,蛋白磷酸酶基因PP2C56、PP2C24和PP2C8的表达模式相同,在70 DAB和120 DAB时上调表达;脱落酸受体PYL2在70 DAB和170 DAB上调表达。在生长素信号中,GH3.6、GH3.1和SAUR36及IAA27在生长素生物合成途径上起重要作用。其中,GH3.6在大果芽变果实发育早期上调表达;IAA27在大果芽变果实发育过程中上调表达,且呈上升趋势;SAUR36和GH3.1在70 DAB时上调表达。生长素促进蛋白6B和22D的表达模式不同,生长素促进蛋白6B在120 DAB上调表达,而生长素促进蛋白22D在70 DAB上调表达。细胞分裂素信号上的差异基因是双组分响应调节基因ARR2和ARR5。这2个基因在70 DAB时的表达趋势相反,但在120 DAB时均下调表达;茉莉酸信号上的基因TIFY10A和水杨酸信号上的基因TGA9在大果芽变中下调表达。细胞周期D3和木葡聚糖内转葡萄糖基酶XTH23属于油菜素内酯信号途径上的基因,细胞周期D3基因在120 DAB的大果芽变中显著下调表达,但在170 DAB时显著上调表达。XTH23的2个成员在70 DAB时上调表达,在120 DAB及170 DAB时下调表达。
3 讨 论
3.1 大果芽变果实变大的因素
果实大小是评价果实外观的重要性状,直接影响果实的经济效益。果实大小变异的产生方式主要有两种,一种是染色体加倍产生的大果型变异,另一种是体细胞变异产生的大果型芽变。在本研究中,流式细胞仪的鉴定结果表明,大果锦红的倍性仍是二倍体,说明大果锦红的产生不是由染色体数目加倍造成的。同时,大果芽变的果实质量是锦红的1.7倍,且自盛花后70 d,大果芽变的果肉横径均显著大于锦红。对果肉进一步分析发现,与锦红相比,大果芽变汁胞数量多且体积大,说明大果芽变的果实变大主要归于果肉厚度的变化,这与EI-Otmani等[10]的结果相一致。果实的最终大小受细胞分裂和细胞膨大控制。芽变产生的大果型基本是由果肉细胞变大和细胞数量增多所产生的。蒋爽等[24]的研究表明,大翠冠果实变大的原因是果肉细胞变大。舒莎珊等[25]的研究表明,潘庄大翠冠的果实增大是由细胞数量增多引起的。在本研究中,通过石蜡切片技术分析锦红和大果芽变汁胞内细胞形态,结果发现大果芽变汁胞薄壁细胞层数多于锦红,说明细胞数量增多导致大果芽变汁胞变大,从而使柑橘果肉增厚,果实变大。
在生产上,人们采用2, 4-DP[10,26]、3, 5, 6-TPA[9]等生长素调节剂来增大柑橘果实。研究发现他们可通过细胞大小影响汁胞发育,从而改变果肉厚度[10]。本研究中,在70 DAB和120 DAB,大果芽变中生长素含量显著高于锦红,说明生長素可能是引起大果芽变和锦红汁胞发育不同的重要激素。而在170 DAB时两者体内生长素含量有所下降,这与冯贵芝[27]的研究结果相同,原因可能是此时果实膨大速率开始减慢,逐步进入成熟期,果实体内各激素水平发生改变。赤霉素控制细胞的伸长,并与果实大小和果实质量呈正相关[28]。在本研究中,大果芽变内GA3含量在各发育时期均高于锦红,这表明大果芽变果实大小的变异可能与赤霉素有关。玉米素是一种嘌呤类的细胞分裂素。Gan等[19]研究表明,细胞分裂素参与调控番茄果皮厚度和果实大小。在本研究中,大果芽变中玉米素含量始终显著低于锦红,同石蜡切片的结果相矛盾,原因可能有两个,一是玉米素只是细胞分裂素的一种,不能代表汁胞内细胞分裂素含量;二是采样时间不对,细胞分裂素大多在果实发育早期起作用,而70 DAB及之后两个时期已进入膨大阶段。
3.2 大果芽变汁胞的转录组分析
本研究中对锦红和大果芽变70 DAB、120 DAB和170 DAB汁胞进行转录组分析,发现3个时期的差异基因数量相差较大,其中70 DAB时差异基因最多,170 DAB时差异基因最少,说明在70 DAB锦红和大果芽变之间的基因表达差异性更显著,该时期可能是引起两者最终果实大小差异的重要时期。KEGG结果表明,差异基因富集在植物激素信号传导途径,并且24个差异基因在两个及以上时期差异表达,其中19个已知功能的差异基因多是花后70和120 d时在锦红和大果芽变汁胞内差异显著。19个基因中,生长素类的差异基因数目最多,同时激素测定结果也显示生长素在几种激素内含量偏高,说明生长素合成相关基因在锦红和大果芽变汁胞发育过程中表达活跃并存在显著差异,造成两者汁胞内生长素含量不同,使他们汁胞发育存在差异,从而导致果实大小不同。GH3.6和IAA27在大果芽变中的表达量均显著高于锦红,Su等[15]的研究表明,SlIAA17通过影响细胞大小来参与调控果实大小。Yang等[29]发现,茉莉酸可能是影响蓝莓果实发育早期大小的关键植物激素,并协调其他植物激素共同促进果实生长发育,并指出茉莉酸信号相关基因TIFY9、TIFY10A表达量显著高于其他激素信号上的基因。这一发现同本研究结果类似。
此外,本研究中还发现油菜素内酯、细胞分裂素等激素相关基因在锦红和大果芽变汁胞内差异表达。双组分响应调节基因ARR能被细胞分裂素诱导并调节生长发育。在本研究中,ARR基因在锦红和大果芽变中表达水平存在差异,且汁胞内玉米素含量不同,说明他们可能受到细胞分裂素的诱导参与调控果实大小。CycD3是细胞周期蛋白基因,通过参与BR激素信号传导刺激细胞分裂[30]。在本研究中,锦红和大果芽变的CycD3表达量在花后70 d时差异不显著,而在花后120 d和170 d差异显著。同时,XTH23作为BR信号传导途径上的另一个重要基因,在锦红和大果芽变间表达量差异显著,说明油菜素内酯可能也参与柑橘果实大小的发育,但其基因功能还需进一步验证。
4 结 论
笔者通过对锦红和大果芽变果实内囊瓣数量、汁胞数量及体积进行测量和统计,初步明确汁胞发育是大果芽变的变异原因,石蜡切片进一步表明,细胞数量可能是引起汁胞变大的细胞学因子。通过对两者间激素含量的比较分析,认为植物激素能影响汁胞发育。通过分析锦红和大果芽变在盛花后70、120和170 d的转录组数据,筛选出IAA27、TIFY10A、CycD3等激素合成相关基因。
参考文献 References:
[1] 常平凡,宗颖生,李玉萍,解晓悦,刘永兰. 不同等级富士苹果价格差别分析[J]. 山西农业大学学报(社会科学版),2002,1(1):27-28.
CHANG Pingfan,ZONG Yingsheng,LI Yuping,XIE Xiaoyue,LIU Yonglan. Analysis of the price difference of Fuji apple of various varying grade[J]. Journal of Shanxi Agricultural University (Society Science Edition),2002,1(1):27-28.
[2] GOFFINET M C,ROBINSON T L,LAKSO A N. A comparison of ‘Empire apple fruit size and anatomy in unthinned and hand-thinned trees[J]. Journal of Horticultural Science,1995,70(3):375-387.
[3] GUARDIOLA J L,GARC?A-LUIS A. Increasing fruit size in Citrus. Thinning and stimulation of fruit growth[J]. Plant Growth Regulation,2000,31(1):121-132.
[4] MOSTAFA E,SALEH M. Response of balady mandarin trees to girdling and potassium sprays under sandy soil conditions[J]. Research Journal of Agriculture and Biological Sciences,2006,2(3):137-141.
[5] AS?N L,VILARDELL P,BONANY J,ALEGRE S. Effect of 6-BA,NAA and their mixtures on fruit thinning and fruit yield in ‘Conference and ‘Blanquilla pear cultivars[J]. Acta Horticulturae,2010,884:379-382.
[6] STERN R A,BEN-ARIE R. GA3 inhibits flowering,reduces hand-thinning,and increases fruit size in peach and nectarine[J]. The Journal of Horticultural Science and Biotechnology,2009,84(2):119-124.
[7] AGUSTI M,ALMELA V,ANDREU I,JUAN M,ZACARIAS L. Synthetic auxin 3,5,6-TPA promotes fruit development and climacteric in Prunus persica L. Batsch[J]. The Journal of Horticultural Science and Biotechnology,1999,74(5):556-560.
[8] AGUST? M,GARIGLIO N,CASTILLO A,JUAN M,ALMELA V,MART?NEZ-FUENTES A,MESEJO C. Effect of the synthetic auxin 2,4-DP on fruit development of loquat[J]. Plant Growth Regulation,2003,41(2):129-132.
[9] AGUST? M,ZARAGOZA S,IGLESIAS D J,ALMELA V,PRIMO-MILLO E,TAL?N M. The synthetic auxin 3,5,6-TPA stimulates carbohydrate accumulation and growth in citrus fruit[J]. Plant Growth Regulation,2002,36(2):141-147.
[10] EL-OTMANI M,AGUST? M,AZNAR M,ALMELA V. Improving the size of ‘Fortune? mandarin fruits by the auxin 2,4-DP[J]. Scientia Horticulturae,1993,55(3/4):283-290.
[11] STERN R A,FLAISHMAN M,APPLEBAUM S,BEN-ARIE R. Effect of synthetic auxins on fruit development of ‘Bing cherry (Prunus avium L.)[J]. Scientia Horticulturae,2007,114(4):275-280.
[12] STERN A R,BEN-ARIE R,APPLEBAUM S,FLAISHMAN M. Cytokinins increase fruit size of ‘Delicious and ‘Golden Delicious (Malus domestica) apple in a warm climate[J]. The Journal of Horticultural Science and Biotechnology,2006,81(1):51-56.
[13] 陳双双,钟嵘,黄春辉,徐小彪,贾东峰,陶俊杰. 不同浓度氯吡脲对‘东红猕猴桃果实品质的影响[J]. 江西农业大学学报,2022,44(3):549-559.
CHEN Shuangshuang,ZHONG Rong,HUANG Chunhui,XU Xiaobiao,JIA Dongfeng,TAO Junjie. Effects of different concentrations of chlorfenuron on fruit quality of ‘Donghong kiwifruit[J]. Acta Agriculturae Universitatis Jiangxiensis,2022,44(3):549-559.
[14] 郭淑萍,杨顺林,杨玉皎,张永辉,孟富宣,何建军,张俊松,金杰. GA3和CPPU对无核翠宝葡萄果实品质的影响[J]. 果树学报,2022,39(10):1834-1844.
GUO Shuping,YANG Shunlin,YANG Yujiao,ZHANG Yonghui,MENG Fuxuan,HE Jianjun,ZHANG Junsong,JIN Jie. Effect of GA3 and CPPU treatments on fruit quality of Wuhe Cuibao grape[J]. Journal of Fruit Science,2022,39(10):1834-1844.
[15] SU L Y,BASSA C,AUDRAN C,MILA I,CHENICLET C,CHEVALIER C,BOUZAYEN M,ROUSTAN J P,CHERVIN C. The auxin Sl-IAA17 transcriptional repressor controls fruit size via the regulation of endoreduplication-related cell expansion[J]. Plant and Cell Physiology,2014,55(11):1969-1976.
[16] PENG Z,LI W X,GAN X Q,ZHAO C B,PAUDEL D,SU W B,LV J,LIN S Q,LIU Z L,YANG X H. Genome-wide analysis of SAUR gene family identifies a candidate associated with fruit size in loquat (Eriobotrya japonica Lindl.)[J]. International Journal of Molecular Sciences,2022,23(21):13271.
[17] ZHAO J Y,JIANG L,CHE G,PAN Y P,LI Y Q,HOU Y,ZHAO W S,ZHONG Y T,DING L,YAN S S,SUN C Z,LIU R Y,YAN L Y,WU T,LI X X,WENG Y Q,ZHANG X L. A functional allele of CsFUL1 regulates fruit length through repressing CsSUP and inhibiting auxin transport in cucumber[J]. The Plant Cell,2019,31(6):1289-1307.
[18] ZHANG L,XU Y,LI Y T,ZHENG S S,ZHAO Z M,CHEN M L,YANG H J,YI H L,WU J X. Transcription factor CsMYB77 negatively regulates fruit ripening and fruit size in citrus[J]. Plant Physiology,2024,194(2):867-883.
[19] GAN L J,SONG M Y,WANG X C,YANG N,LI H,LIU X X,LI Y. Cytokinins are involved in regulation of tomato pericarp thickness and fruit size[J]. Horticulture Research,2022,9:uhab041.
[20] WANG X C,ZHAO M Z,WU W M,KORIR N K,QIAN Y M,WANG Z W. Comparative transcriptome analysis of berry-sizing effects of gibberellin (GA3) on seedless Vitis vinifera L.[J]. Genes & Genomics,2017,39(5):493-507.
[21] SU W B,SHAO Z K,WANG M,GAN X Q,YANG X H,LIN S Q. EjBZR1 represses fruit enlargement by binding to the EjCYP90 promoter in loquat[J]. Horticulture Research,2021,8:152.
[22] 朱凱杰. 脐橙果皮色泽变异及番茄红素β-环化酶的调控研究[D]. 武汉:华中农业大学,2020.
ZHU Kaijie. Color variation of navel orange flavedo and regulatory analysis of lycopene β-cyclase[D]. Wuhan:Huazhong Agricultural University,2020.
[23] 卜海东. ‘龙丰苹果大果变异形成机制解析[D]. 沈阳:沈阳农业大学,2021.
BU Haidong. Formation mechanism analysis of the ‘Longfeng apple large fruit variation[D]. Shenyang:Shenyang Agricultural University,2021.
[24] 蒋爽,骆军,王晓庆,李水根,周博强. 翠冠梨大果芽变果实组织切片、激素变化及转录组分析[J]. 果树学报,2022,39(10):1737-1747.
JIANG Shuang,LUO Jun,WANG Xiaoqing,LI Shuigen,ZHOU Boqiang. Tissue sections,hormone changes,and transcriptomes analysis of the large-fruited bud mutation of Pyrus pyrifolia ‘Cuiguan[J]. Journal of Fruit Science,2022,39(10):1737-1747.
[25] 舒莎珊,李鑫,骆军,白松龄,滕元文. ‘翠冠梨大果型芽变的细胞学及相关基因表达研究[J]. 果树学报,2017,34(6):660-669.
SHU Shashan,LI Xin,LUO Jun,BAI Songling,TENG Yuanwen. Studies on cytology and related gene expression pattern of a large-fruited bud mutant from ‘Cuiguan pear (Pyrus pyrifolia)[J]. Journal of Fruit Science,2017,34(6):660-669.
[26] YILDIRIM B,YE?ILO?LU T,?NCESU M,KAMILO?LU M U,?IMEN B,TAMER ?. Effects of 2,4-DP (2,4-dichlorophenoxypropionic acid) plant growth regulator on fruit size and yield of Valencia oranges (Citrus sinensis Osb.)[J]. New Zealand Journal of Crop and Horticultural Science,2012,40(1):55-64.
[27] 馮贵芝. 基于时空转录组的脐橙果实发育和成熟调控机理的研究[D]. 武汉:华中农业大学,2021.
FENG Guizhi. Research on regulating mechanism of navel orange development and maturation based on spatiotemporal transcriptome[D]. Wuhan:Huazhong Agricultural University,2021.
[28] ROKAYA P R,BARAL D R,GAUTAM D M,SHRESTHA A K,PAUDYAL K P. Effect of pre-harvest application of gibberellic acid on fruit quality and shelf life of mandarin (Citrus reticulata Blanco)[J]. American Journal of Plant Sciences,2016,7(7):1033-1039.
[29] YANG L,LIU L M,WANG Z Y,ZONG Y,YU L,LI Y Q,LIAO F L,CHEN M M,CAI K L,GUO W D. Comparative anatomical and transcriptomic insights into Vaccinium corymbosum flower bud and fruit throughout development[J]. BMC Plant Biology,2021,21(1):289.
[30] HU Y X,BAO F,LI J Y. Promotive effect of brassinosteroids on cell division involves a distinct CycD3-induction pathway in Arabidopsis[J]. The Plant Journal,2000,24(5):693-701.
猜你喜欢
保健医苑(2022年4期)2022-05-05
猪业科学(2022年2期)2022-04-21
基层中医药(2021年10期)2021-06-05
中国生殖健康(2020年2期)2021-01-18
中国生殖健康(2019年8期)2019-01-07
中国生殖健康(2018年2期)2018-11-06
湖南农业(2016年3期)2016-06-05
浙江柑橘(2016年4期)2016-03-11
中国当代医药(2015年7期)2015-03-01
发明与创新(2015年1期)2015-02-27
