
猪ESM-1 基因的结构及其遗传特性分析
2024-04-12 05:53郑美丽陈少康孙建华王晓凤
猪业科学 2024年3期
郑美丽 ,陈少康 ,孙建华 ,李 爽 ,郭 峰 ,王晓凤
(1.北京市畜牧总站,北京 100107;2.北京市通州区动物疫病预防控制中心,北京 101199)
内皮特异性分子1(endothelialspecf ic molecule-1,ESM-1),最初是由Lassalle等发现的。 Bechard证实了ESM-1是作为一种可溶性的蛋白多糖-硫酸软骨素分泌至血液中,又称为Endocan。早期文献报道了ESM-1基因的结构与类胰岛素样生长因子结合蛋白达到32%相同和55.3%相似,尤其是该基因N端结构与IGFBP家族高度同源。而IGFBP家族作为IGF家族的构成,已经有多个成员被证明与机体的生长发育密切相关。笔者对ESM-1基因的结构以及在不同猪种中的遗传特性进行了初步探究,以期为深入了解和研究ESM-1基因的结构和功能,将该基因作为影响猪肌肉生长发育的关键基因的研究提供参考。
1 材料与方法
1.1 样本的采集
从实验室群体样本中选取了三个国外代表性猪种(大白、长白、杜洛克),中国野猪(东北野猪)和中国地方猪种(二花脸、上高猪、民猪、内江猪、香猪)5个代表性猪种,每个猪种选取的个数如表1,试验样本为肌肉组织。

表1 样品选取个数
1.2 试验方法
1.2.1ESM-1基因的完整mRNA序列的获得
基于NCBI预测的ESM-1基因的CDS区序列,通过PCR和RACE试验获得ESM-1基因的完整mRNA序列。试验中所用到的引物序列如表2。
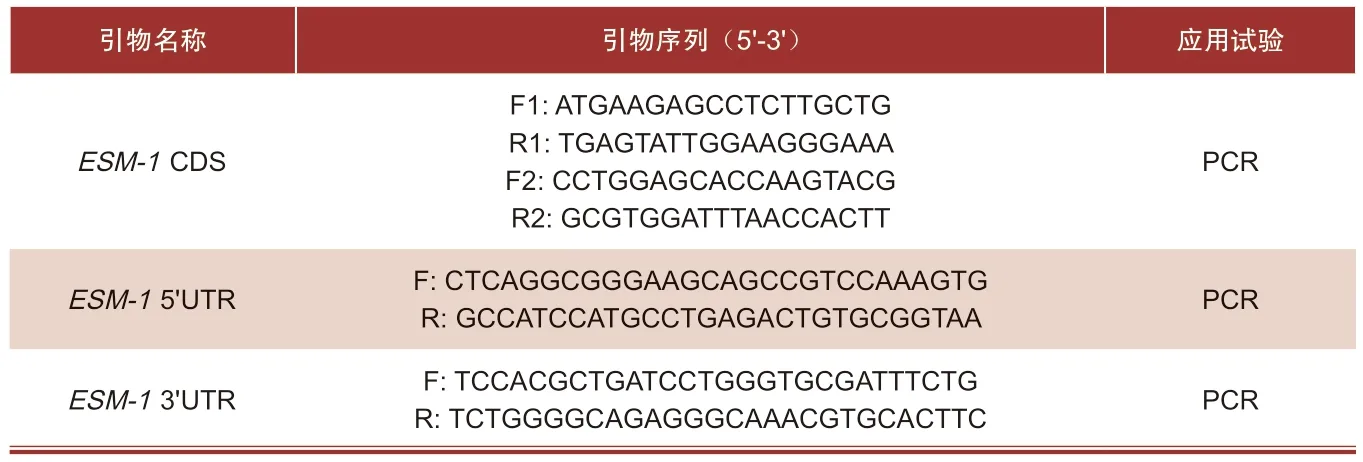
表2 引物序列
1.2.2 外显子SNPs的检测
以克隆测序得到的ESM-1基因的mRNA序列为模板,利用软件primer5.0设计引物如表3,引物位置编码按照以CDS区的ATG为0,到TGA结束为552 bp,ATG之前为“-”TGA后为“+”。
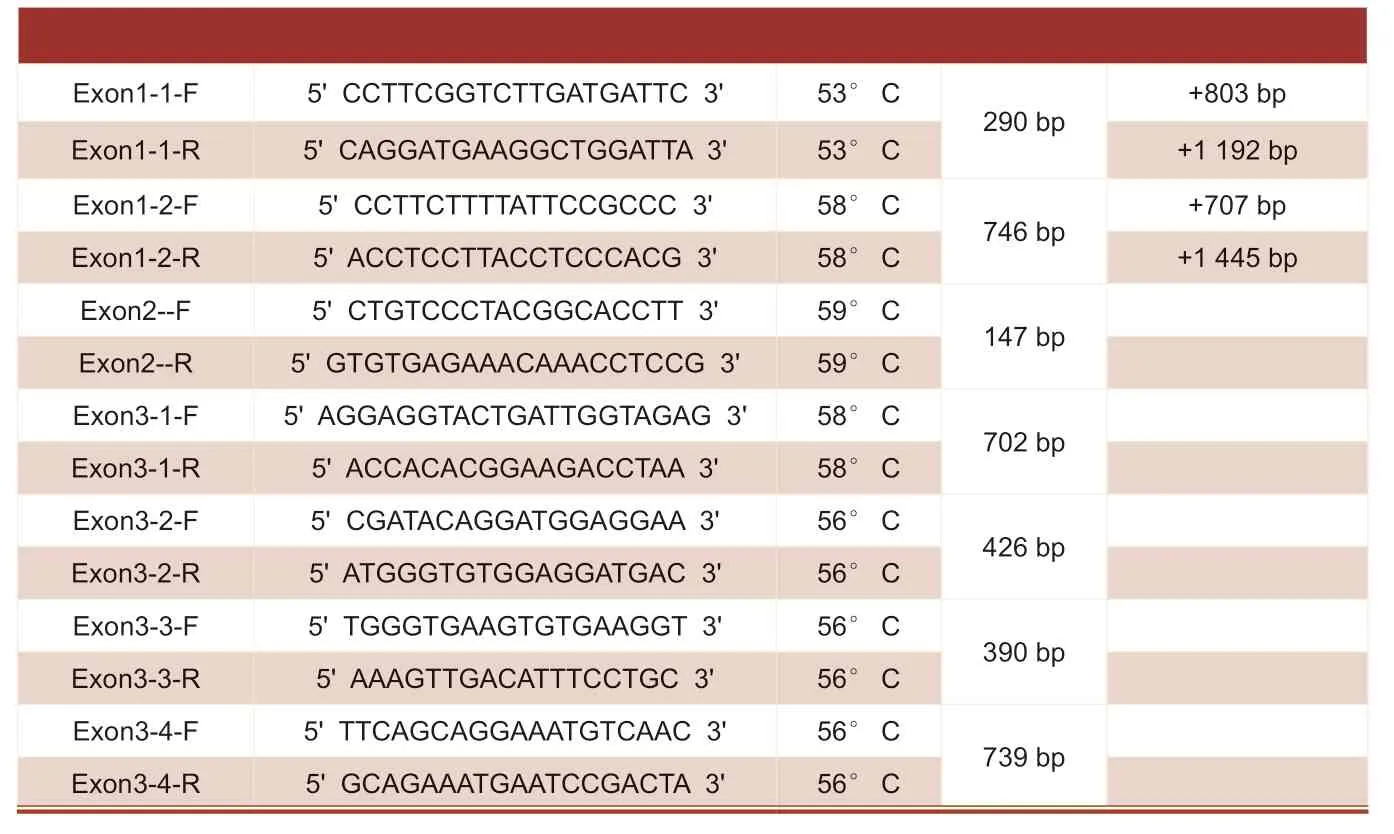
表3 引物序列
2 试验结果
2.1 猪ESM-1 基因的克隆
2.1.1 CDS区的克隆测序
根据NCBI上可查的猪ESM-1基因的预测CDS区序列,设计引物,PCR扩增,所得产物经琼脂糖凝胶电泳检测为单一明亮条带,如图1。将产物回收纯化并连接转化后,测序并将所得到的无载体序列与NCBI上序列进行比对分析,获得了猪ESM-1基因的CDS区准确序列552 bp。

图1 猪ESM-1 基因CDS 区扩增产物
2.1.2 mRNA的克隆测序
利用RACE技术并结合试剂盒中的巢式引物获得无载体序列的5'UTR和3'UTR序列,分别为439 bp和1 447 bp,如图2。经克隆测序及比对分析,确定为猪ESM-1基因的5'及3'UTR区域。结合前面试验得到的CDS区序列,经DNAMAN及ChromaPro拼接,形成全长2 437 bp的猪ESM-1基因的mRNA序列。在NCBI上将所得序列与猪的基因组信息进行了比对,序列全部符合,并且内含子边界符合“ GT-AG”法则。

图2 猪ESM-1 基因5'RACE(左)及 3'RACE(右)扩增产物
2.2 外显子SNPs 的检测
用3 中的7 对引物扩增,以NCBI上公布的猪基因组序列为野生型,经测序比对,在编码区和外显子发现了SNP位点,检测到的SNP位点如表4。进一步增加样本数量,进行群体验证,并通过酶切PCR产物,部分琼脂糖凝胶电泳图如图3,发现这几个SNP位点在不同猪种之间没有显著性突变差异,其CDS区无有意义突变。具体基因型及基因频率如表5。
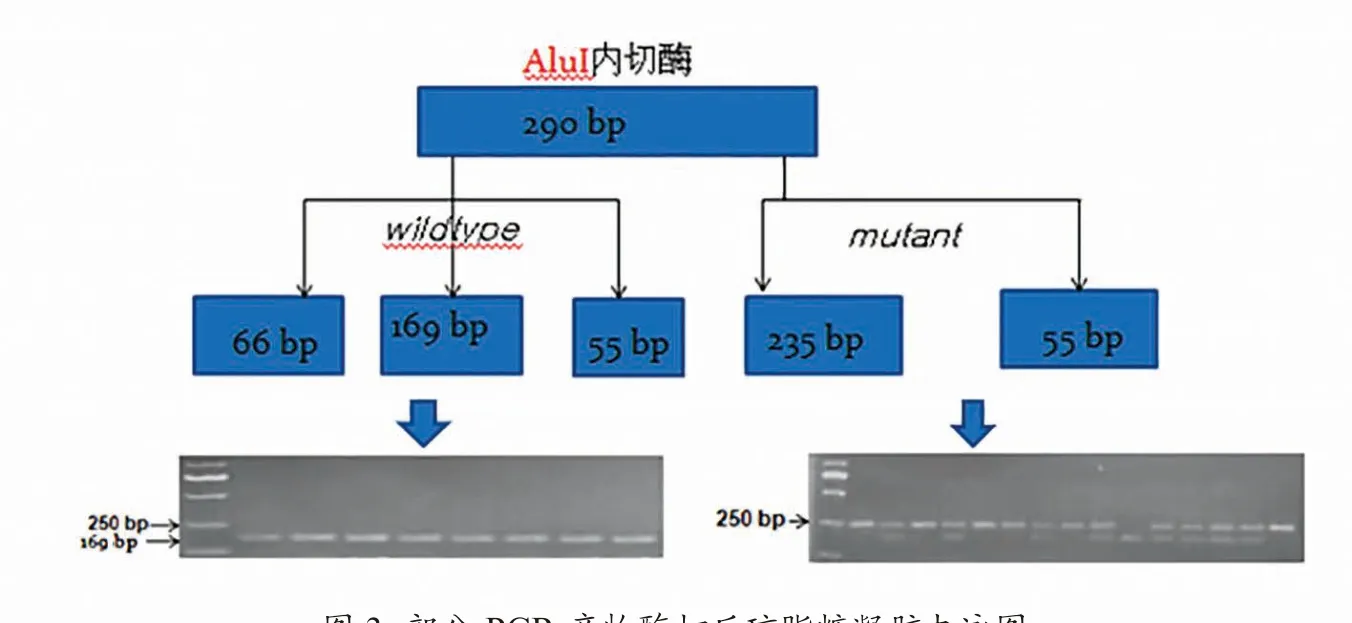
图3 部分PCR 产物酶切后琼脂糖凝胶电泳图

表4 检测到的突变信息

表5 各基因型及基因频率(1)第一外显子第8 位碱基突变位点
3 讨论与分析
目前猪的ESM-1基因在NCBI上只有预测的CDS区序列,而在本研究中,笔者利用了RACE技术,首次得到了猪ESM-1基因的完整mRNA序列,通过比对发现,所得到的序列全部符合,进一步分析该基因的结构,可知猪ESM-1基因由三个外显子组成,CDS区为552 bp,5'UTR和3'UTR序列,分别为439 bp和1 447 bp,如图4。

图4 猪ESM-1 基因mRNA 的结构示意图
笔者针对基因的三个外显子进一步做了遗传多态性分析,希望为该基因的功能研究提供一定的分子基础。单核苷酸的多态性(Single Nucleotide Polymorphisms,SNP),主要是指在基因组水平上由单个核苷酸的变异所引起的DNA序列的多态性。SNP所表现的多态性只涉及到单个碱基的变异,这种变异可由单个碱基的转换(transition)或颠换(transversion)所引起,也可由碱基的插入或缺失所致。在基因组的DNA中,任何碱基均有可能发生变异,因此SNP既有可能在基因序列内,也有可能在基因以外的非编码序列上。据目前研究可知,位于编码区内的SNP(coding SNP,cSNP)比较少,因为在外显子内,其变异率仅仅是周围序列的1/5,但它在遗传学方面的研究中却具有重要意义,因此cSNP的研究更受关注。另一方面,从对生物的遗传性状影响上来看,cSNP又可分为2种:一种是同义cSNP(synonymous cSN),即SNP所致的编码序列的改变并不影响其所翻译的蛋白质的氨基酸序列,突变碱基与未突变碱基的含义相同;另一种是非同义cSNP(non-synonymous cSNP),指碱基序列的改变可使以其为蓝本翻译的蛋白质序列发生改变,从而影响了蛋白质的功能,这种改变常是导致生物性状改变的直接原因。
虽然ESM-1基因在人和小鼠上的研究比较多,但目前针对ESM-1基因的遗传多态性研究几乎为空白,NCBI的数据库中也未查到相关记录。而笔者第一次针对猪的ESM-1基因进行了遗传多态性分析,并在外显子上发现了两个碱基突变位点。笔者根据1986年中国农业科学院畜牧研究所张仲葛主持编写的《中国猪品种志》,从中国六大地方猪(华北型、华中型、华南型、江海型、西南型、高原型)中尽量选取了代表性猪种,同时也选取东北野猪作为中国野猪的代表,大白、长白和杜洛克三种国外代表性猪种,对ESM-1基因在不同猪种中的遗传多样性进行了分析,试验检测到的两个突变位点分别位于5'UTR和CDS区上,但对后者分析,其突变并不会导致蛋白质的改变,为同义突变,因此认为猪ESM-1基因的CDS区具有较高的保守型,最后通过对两个SNPs位点进行群体验证,发现两处突变都没有明显的品种间差异。
4 研究进展与展望
ESM-1是作为一种可溶性的蛋白多糖-硫酸软骨素分泌至血液中,又称为Endocan。ESM-1由单一基因编码,定位在人的第5号常染色体上,mRNA全长2 090 bp,共由3个外显子组成。其中外显子2编码一个富含基因F(113FPFFQY I 18)结构的特殊区域,该区域可能是ESM-1基因的功能区域。最大的外显子3编码ESM-1蛋白C端33个氨基酸区域,该区域含有提前终止密码子和137位Ser O糖基化位点,该位点以共价键的形式连接一条黏多糖单链,也称GAG链或DS单链,该黏多糖单链与ESM-1的多种生物学功能紧密相关。目前的研究表明,ESM-1基因的mRNA能编码两种不同的蛋白,如图5。一种是ESM-1,该蛋白是一种相对分子质量为50×1 000的可溶性蛋白聚糖,是一条成熟的包含165氨基酸的核心蛋白,包含一个F113和F116的苯丙氨酸富集区,以及以共价键的形式连接在第137丝氨酸残基上的一条黏多糖单链。

图5 猪ESM-1 基因mRNA 的结构示意图
ESM-1基因目前的研究主要集中人和鼠上。首先早期试验证明该基因有助于血管内皮细胞的功能,并在内皮细胞依赖性病理过程中起一定的作用。Mark Aitkenhead等发现ESM-1基因在血管生成过程中发挥着重要的作用。一些涉及生长和血管生成的因子已经显示对ESM-1基因的表达有影响,同时有文献报道指出肝细胞生长因子(HGF)和血管内皮生长因子(VEGF)共同作用会导致该基因在细胞内的表达上调。有研究证明HHEX对血管的增生以及塑形都有着重大的作用,HHEX能通过减少血管内皮细胞生长因子受体(vascular endothelial growth factor receptor-1,VEGFR)如VEGFR-1、VEGFR-2等和血管生成相关的一些重要因子的表达来参与血管生成的调节,而研究者同时发现同源异形框因子(HHEX)可通过一个高度保守的HHEX反应元件直接抑制ESM-1的表达。那么,HHEX 对VEGFR-1和VEGFR-2等血管增生中重要因素的抑制作用是否与其直接抑制ESM-1的表达有关,这还有待进一步研究。
也有相关文献指出ESM-1基因与胰岛素样生长因子相关,能够促进细胞的生长和增殖。在文献中Philippe Lassalle等指出,该基因的N端与类胰岛素样生长因子结合蛋白达到32%相同和55.3%相似。而IGFs生长因子在血清,细胞外液及细胞培养液中都与特异性的结合蛋白(Binding Proteins,BPs)结合以无活性的复合物形式存在,具有延长循环水平的IGFs半衰期和稳定IGFs血清浓度的作用,因此进一步推测ESM-1基因对生命体的生长发育有影响。
最后ESM-1研究比较多的一方面即其与各种肿瘤的发生有关。一些肿瘤如肾、乳腺、子宫以及直肠等肿瘤组织中存在内皮细胞特异性分子(ESM-1)表达的上调,ESM-1基因在肿瘤血管生产和肿瘤生长过程中发挥着重要作用。该基因能通过肿瘤相关性的炎症、血管增生、淋巴管增生、肿瘤细胞本身等方面对肿瘤进行调节。有文献探究了ESM-1基因在肝癌发生中,影响肿瘤细胞的存活、迁移和迁徙。而肿瘤的产生本身就是一种细胞的异常增生现象,这也进一步证明ESM-1基因对细胞的生长具有重要的作用。Grigoriu 等的研究发现,肺癌患者血清中的ESM-1浓度,不仅与肿瘤的类型、分期和患者的预后、肿瘤的进展相关,更能在一定程度上指导肺癌的治疗。同样,Leroy 等发现肾透明细胞癌患者血清中,ESM-1的浓度可作为肾透明细胞癌患者术后治疗和抗血管增生治疗的一个有效监测指标。在诸如肺癌、乳腺癌、子宫癌、肾癌、涎腺腺样囊性癌、肝癌、脑神经胶质细胞瘤等许多人体实体肿瘤中ESM-1都高表达,同时也发现该因子的高表达与肿瘤的预后、转移、血管增生等因素有关。ESM-1与肿瘤发生、发展的重要因素(如慢性炎症、肿瘤细胞本身、血管增生、淋巴管增生等)都有着重要的关联。如果能有效地抑制ESM-1的表达,就有可能从多方面对肿瘤的发生、发展进行抑制。因此,开发针对ESM-1的靶向治疗药物有可能开拓肿瘤治疗的一个新篇章。
目前,对于ESM-1基因表达的研究还处于资料积累的阶段,关于其对家畜尤其是猪的肌肉生长影响的研究还较少,该基因是否可以调控猪的肌肉生长发育,影响猪的生产性状,仍需进一步研究。
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29
猪业科学(2022年10期)2022-11-03
电子科技大学学报(2022年5期)2022-10-29
猪业科学(2021年6期)2021-08-12
猪业科学(2021年3期)2021-05-21
中国生殖健康(2020年4期)2021-01-18
教学考试(高考生物)(2020年6期)2020-11-23
食品与生物技术学报(2020年8期)2020-01-06
科学24小时(2019年5期)2019-06-11
发明与创新(2019年9期)2019-03-26
